
Abstract
Skin cancers and extrinsic aging are delayed consequences of cumulative UV radiation insults. Exposure of human keratinocytes to UVB has been previously shown to trigger premature senescence. In order to explore the involvement of the cyclin-dependent kinase inhibitor p16INK-4a in UVB-induced premature senescence, we developed an original model of repeated sublethal exposures of human keratinocytes deficient in p16INK-4a. We did not observe any significant increase of senescence-associated β-galactosidase activity positive cells following UVB exposure in this cell line in contrast to primary keratinocytes, suggesting a role for p16INK-4a in UVB-induced senescence. However, we detected sustained DNA damage, prolonged cell cycle arrest, and induction of markers of epidermal differentiation like involucrin and filaggrin as consequences of the repeated exposures. Keratinocytes exposed to the same dose of UVB in a single exposure died. Furthermore, the abundance of the keratins 6, 16 and 17 was increased in keratinocytes exposed repeatedly to UVB suggesting an alternative differentiation. This model allows the induction of a state of differentiation observed in vivo with differentiation uncoupled from premature senescence.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Skin protects the organism against environmental insults, including ultraviolet radiation (UV). The epidermis covers the skin surface and is a multilayered epithelium based on constantly regenerating keratinocytes. Keratinocytes proliferate in the basal layer before moving upwards to the suprabasal layers through a complex differentiation programme that culminates in fully differentiated dead cells in the cornified superficial layer, maintaining a strong impermeable barrier (Eckert et al. 1997). Skin and the epidermis are variably but repeatedly exposed to UV, resulting in epidermal malignancies that represent a serious health threat around the world (Ichihashi et al. 2003). UV also cause skin photoaging which refers to the effects of long-term UV exposure and UV damage surimposed on intrinsically aged skin (Rabe et al. 2006; Rittie and Fisher 2002). UVB (290–320 nm) are the most deleterious components of sunlight reaching the earth surface (Ichihashi et al. 2003). UVB interact with cellular chromatophores and produce reactive oxygen species (ROS) that cause severe cellular oxidative damage and activate multiple cellular signaling pathways related to cell growth, differentiation, and replicative senescence, but also linked to connective tissue degradation and inflammation (Rittie and Fisher 2002). DNA is the major chromophore for UVB, explaining the high mutagenicity of UVB (Ichihashi et al. 2003).
Senescent cells remain metabolically active and contribute to skin ageing (Gandarillas 2000). The senescent phenotype is characterized namely by an increase of cell surface, sustained expression of cyclin-dependent kinase inhibitors, senescence-associated β-galactosidase activity (SA β-gal) and decreased proteasome activities (Blackburn 2005; Dimri et al. 1995; Petropoulos et al. 2000). Replicative senescence is characterized by a permanent arrest of cell proliferation (Hayflick and Moorhead 1961) resulting from the critical erosion of telomeres after serial cellular divisions (Blackburn 2005). Senescence can also be prematurely induced by oxidative stress, DNA damaging agents or oncogene activation (Ben-Porath and Weinberg 2005; Toussaint et al. 2002). Lewis et al. (2008) demonstrated that premature senescence can occur in primary keratinocytes after exposure to a single dose of UVB through a mechanism involving p53 and the cyclin-dependent kinase inhibitor p21WAF−1 expression. However, in vivo, the cyclin-dependent kinase inhibitor p16INK-4a was shown to be overexpressed in the dermis and epidermis of aged persons (Ressler et al. 2006). This prompted us to test whether keratinocytes lacking p16INK-4a can still undergo premature senescence after exposure(s) to UVB. In order to develop a model which would allow further functional studies with stable overexpressions or invalidations of genes, we used a line of human keratinocytes lacking p16INK-4a and expressing human telomerase (hTERT), while retaining other growth controls and the ability to fully differentiate in vitro in a reconstructed epidermis (Dickson et al. 2000). Expression of hTERT does not prevent stress-induced senescence in normal cells induced by either acute sublethal short exposures to H2O2, UV, UVB or gamma-irradiation (de Magalhaes et al. 2002; Gorbunova et al. 2002; Matuoka and Chen 2002). Telomerase expression alone cannot immortalize human keratinocytes. However, telomerase-expressing human keratinocytes that exhibit an impaired expression of p16INK-4a are immortal (Dickson et al. 2000). Premature senescence, or stasis, has already been induced in a variety of immortal and transformed cells (for reviews, (Dierick et al. 2003; Schmitt 2007)). In addition, expression of telomerase allows studying the effects of DNA damaging agents like UVB on premature senescence, independently of critical telomere erosion. Indeed it has been shown that stress-induced premature senescence, due to exposure to acute sublethal short exposures to H2O2 or UVB, can take place independently of critical telomere shortening (Dumont et al. 2001).
Materials and methods
Reagents and cell culture
Immortal human keratinocytes ectopically expressing the catalytic subunit of telomerase and characterized by inactivated p16INK-4A gene (N-hTERT keratinocytes) were a kind gift of Pr J. Rheinwald (Dept of Medicine & Harvard Skin Disease Research Center, Boston MA, USA). N-hTERT keratinocytes were grown at 37°C under a 5% CO2 atmosphere in Epilife medium containing human keratinocyte growth supplement (HKGS, Cascade Biologics, Portland OR, USA). Cell cultures were maintained at subconfluence in order to keep control cells (not exposed to UVB) in proliferating conditions, allowing the identification of growth-arrest in cells exposed to UVB. Apoptosis was induced by incubating the cells for 4 h with 500 nM staurosporine (Sigma, Bornem, Belgium) in Epilife medium.
Primary keratinocytes were isolated by the trypsin float technique from normal adult human skin obtained during plastic surgery and were cultured similarly to the N-hTERT cell line. Experiments were performed using primary keratinocytes at low (<4) culture passage.
Exposure(s) to UVB
For one single exposure to UVB, 200,000 keratinocytes were seeded in 25 cm2 flasks 2 days before exposure: the cells were then washed with phosphate-buffered saline pH 7.4 (10 mM phosphate, 0.9% NaCl) (PBS), bathed in 3 ml of PBS and exposed to UVB between 150 and 3,000 mJ/cm2 using three TL 20 W/01 lamps (Philips, Eindhoven, The Netherlands), emitting UVB peaking at 311 nm, and placed at 30 cm above the flasks. The emitted radiation was checked under a flask lid using a UVR radiometer with UVB sensor (Bioblock Scientific, Tournai, Belgium).
For repeated exposures to UVB, keratinocytes were plated (2,000 cells/cm2) 64 h before the first exposure. The cells were washed with PBS and exposed to UVB between 100 and 400 mJ/cm2 in a thin layer of PBS. This exposure was repeated 4 times per day for 2 days. The time interval between exposures 1–4 and 5–8 was 2.5 h, while 16 h separated exposure 4 and 5 in order to mimic the UV-free night period. After each exposure, fresh medium was provided to the cells. Control cells (CTL) were submitted to the same conditions but with no exposure to UVB.
Cell viability
Cell viability was measured 16 h after the last exposure to UVB, using the classical MTT method (Mosmann 1983).
Measurement of DNA synthesis
After 40 h the last exposure to UVB, 1 μCi [3H]–thymidine (DuPont NEN, Boston MA, USA) was added to the medium for 24 h. The DNA-incorporated radioactivity was quantified 64 h later after the last exposure using scintillation counting (Packard Instrument Company, Meriden CT, USA). Data were normalized to the cellular protein content assayed by the Folin method before [3H]–thymidine incorporation.
Confocal microscopy
Cells were fixed with 4% paraformaldehyde (Merck, Darmstadt, Germany) and permeabilized with 1% Triton X-100 (Sigma–Aldrich, Bornem, Belgium). For detection of Cyclobutane Pyrimidine Dimers (CPDs), specific procedures were achieved according to the manufacturer’s instructions (MBL®, Woburn, MA, USA). The following antibodies were used: rabbit anti-active caspase-3 (Promega, Leiden, The Netherlands), mouse anti-filaggrin (Neomarkers, Fremont CA, USA) and rabbit anti-involucrin (Harbor Bio-Products, Boechout, Belgium), as well as specific Alexa Fluor 488 goat anti-rabbit or anti-mouse IgG conjugates (Molecular Probes Leiden, The Netherlands). To visualize the nucleus, the cells were incubated with TO-PRO-3 (Molecular Probes). The coverslips were observed with a TCS confocal microscope (Leica, Solms, Germany) using a constant multiplier. Triplicates were performed.
Protein extraction and Western blot analyses
Cells were lysed (10 mM Tris pH 7.4, 100 mM NaCl, 10% glycerol, 1% NP-40, 0.1% SDS, 0.5% sodium deoxycholate) at 4, 16, 40 or 64 h after the last exposure to UVB and proteins analyzed by SDS–PAGE and Western blotting using the following antibodies: mouse anti-PARP (BD Pharmingen, Erembodegem, Belgium), mouse anti-p21WAF−1 (Santa Cruz, CA, USA), mouse anti-p53 (Upstate, Charlottesville, VA, USA), mouse anti-cytokeratin 6 (K6), anti-cytokeratin 16 (K16) and anti-cytokeratin 17 (K17) (Neomarkers, Fremont, CA, USA), or α-tubulin antibody (Sigma, Bornem, Belgium) and horseradish peroxidase-linked secondary anti-mouse and anti-rabbit antibodies (GE Healthcare, Diegem, Belgium). Triplicates were performed.
Colorimetric assay of DNA binding of p53
DNA-binding activity of p53 was assayed using the TransAM assay according to the manufacturer’s instructions (Active Motif, Carlsbad, CA, USA). Briefly, 5 μg of nuclear proteins were incubated for 1 h in a 96-wells plate coated with a double-stranded oligonucleotide containing the p53 consensus binding site (5′-GGACATGCCCGGGCATGTCC-3′). Total p53 bound to DNA was detected with a rabbit antibody against p53. Colorimetric detection at 450 nm was performed using a secondary HRP-conjugated anti-rabbit IgG antibody. The results are expressed as mean ± SD from three independent experiments.
Detection of oxidized proteins
Immunoblot detection of carbonyl groups was performed with the OxyBlot™ protein oxidation detection kit (Chemicon®, Temecula, CA, USA) according to the manufacturer’s instructions with 2 μg of total proteins. To assess protein loading, membranes were transiently stained with 0.1% Ponceau in 5% acetic acid to visualize proteins transferred to membranes. The results presented are representative from three independent experiments.
Proteasome peptidase activities
At 16 and 64 h after the last exposure to UVB, the cells were trypsinized, harvested and resuspended in 250 μl of lysis buffer [5 mM Tris–HCl, pH 7.5, supplemented with 250 mM saccharose, 0.2 mM MgSO4, 20 mM EDTA, 0.4% Triton X-100, 2 mM DTT and a complete protease inhibitor cocktail (1:25) (Roche, Indianapolis, IN, USA). Cells were lysed with two freeze–thaw cycles from −80 to 4°C followed by a sonication of 2 × 10 s at 4°C. Cellular debris and organelles were removed from crude extracts by centrifugation at 15,000×g for 30 min at 4°C. Peptidase activities of the proteasome (chymotrypsin-like, peptidylglutamyl-peptide hydrolase and trypsin-like activities) were monitored using fluorogenic peptides in a temperature-controlled microplate fluorimetric reader (Fluostar Galaxy, bMG, Stuttgart, Germany) as described in (Bulteau et al. 2002).
Real time RT-PCR
Total RNA was extracted (Total RNAgent extraction kit, Promega, Madison, WI, USA) and reverse transcribed using the SuperScript II Reverse Transcriptase (Invitrogen, Carlsbad, CA, USA). Real time PCRs were performed with the Power SYBR Green PCR Master Mix (Applied Biosystems, Foster City, CA, USA) using the ABI PRISM 7900HT fast Real-Time PCR system (Applied Biosystems). The sequences of the forward and reverse primers are respectively: 5′-ctg gag act ctc agg gtc gaa-3′ and 5′-cca gga ctg cag gct tcc t-3′ for p21WAF−1, 5′-gtg gcc acc caa aca taa ata ac-3′ and 5′-cct agc gga ccc gaa ata agt-3′ for involucrin, 5′-ggg cac tga aag gca aaa ag-3′ and 5′-cac cat aat cat aat ctg cac tac ca-3′ for filaggrin, 5′-gag acc tgt gac ctg gac aat G-3′ and 5′-gga agg agt gga ttt aga tca caa ga-3′ for osteonectin, 5′-tgt ggt tgc ctt gca cga t-3′ and 5′-gct tgt ggg tgt gac ctg agt-3′ for fibronectin, 5′-gga tga agg acc agt gtg aca ag-3′ and 5′-cag cga cct gga ggg att c-3′ for clusterin and 5′-acc cac tcc tcc acc ttt gac-3′ and 5′-gtc cac cac cct gtt gct gta-3′ for the housekeeping gene GAPDH. Values are average of triplicates ± SD.
Cytochemical staining and microfluidic detection of SA β-gal activity
Cytochemistry The detection of senescence-associated β-galactosidase activity (SA β-gal) was determined 64 h after the last exposure to UVB as described by Dimri et al. (1995). The proportions of S-A β-gal-positive cells are given as a percentage of the total number of cells counted in each well. Each result is the mean value obtained by counting 500 cells in three independent culture dishes ± SD.
Microfluidic chip detection of the SA β-gal activity It was achieved as described in (Debacq-Chainiaux et al. 2008). Briefly, to measure the SA β-gal at pH 6, cells were treated with bafilomycin A1 (Sigma, Bornem, Belgium). Fluorogenic membrane-permeable substrate of β-galactosidase C12FDG (Molecular Probes, Leiden, The Netherlands) was added. Cultures were incubated with carboxynaphtofluorescein diacetate (Molecular Probes) to label living cells. Approximately 500 events were recorded per experiment.
Statistical analysis
Statistical analysis was carried out with the Student’s t-test (NS: non significant, *P < 0.05, **P < 0.01 and ***P < 0.001 vs. (respective) control).
Results
Sublethal dose of UVB induces cellular senescence in primary keratinocytes but not in N-hTERT keratinocytes
The viability of primary or N-hTERT keratinocytes exposed to UVB at single doses between 150 and 3,000 mJ/cm2 was determined using the MTT assay, 16 h after the exposure (Fig. 1a). Primary and N-hTERT keratinocytes present a very similar curve with a significant dose-dependent cytotoxicity appearing at doses above 1,200 mJ/cm2.

Viability and SA β-gal detection in primary and in N-hTERT keratinocytes after a single exposure to UVB. a Viability of primary and N-hTERT keratinocytes after a single UVB exposure. At 16 h following a single exposure with various doses of UVB, cell viability was assessed by the MTT method. The results (means of triplicates ± SD) are expressed as percentages of cell survival compared with respective control cells (unexposed to UVB). Statistical analysis was carried out with Student’s t-test (NS: non significant, * P < 0.05, ** P < 0.01, *** P < 0.001 vs. dose 0). b A low dose of UVB induces cellular senescence in primary keratinocytes but not in N-hTERT keratinocytes. Primary and N-hTERT keratinocytes were exposed or not to UVB at 250 or 375 mJ/cm2. Proportion of SA β-gal positive keratinocytes was assessed by cytochemistry at 64 h after exposure. Results (means of triplicates ± SD) are expressed as ratio of positive SA β-gal cells/a respective total cell number of 500 events. (NS, non significant, * P < 0.05, ** P < 0.01 vs. respective control cells). A statistical test between control cells (no UVB) of primary and N-hTERT keratinocytes was also computed
SA β-gal is known to be expressed in keratinocytes of elderly humans (Dimri et al. 1995). Thus, primary and N-hTERT keratinocytes were exposed to a sublethal dose of UVB (250 or 375 mJ/cm2) and then analysed 64 h after the exposure. N-hTERT keratinocytes did not display any significant increase in the number of SA β-gal positive cells, whereas primary keratinocytes did (Fig. 1b), confirming the observations made by Lewis et al. (2008). There is a minor difference in the absolute percentage of positive cells in the controls between primary (14.4 ± 2.8%) and N-hTERT keratinocytes (5.4 ± 0.5%). The fact that primary cells had already been passaged a minimal number of times might explain this little difference. Primary keratinocytes are indeed particularly sensitive to culture conditions (Ramirez et al. 2001). Despite their slightly higher percentage of SA β-gal positive cells, the primary cells still show a large increase of their proportion of SA β-gal positive cells after exposure to UVB.
In everyday’s life, keratinocytes may be exposed daily and repeatedly to the sunlight, including UVB. Since there was no induction of SA β-gal activity in N-hTERT keratinocytes after a single exposure to UVB, we decided to expose this cell type to a series of exposures to UVB at sublethal doses, rather than exposing them to higher UVB doses. In human skin fibroblasts, repeated exposures to UVB are required to induce a premature senescence phenotype (Debacq-Chainiaux et al. 2008). In order to test whether this phenomenon also occurs in N-hTERT keratinocytes, a model of repeated sublethal exposures to UVB was established.
Repeated sublethal exposures to UVB do not induce apoptosis
N-hTERT keratinocytes were exposed four times a day to UVB during 2 days (Fig. 2a). Cell viability was measured 16 h after the last exposure (Fig. 2b). Viable cells just prior the first exposure to UVB were considered as 100%. In this model, cell culture conditions had to be set so that the control cells keep on proliferating. This allowed to characterize the main mechanisms of the growth arrest induced by UVB (see beneath), which could have been partly similar to mechanisms of growth inhibtion due to contacts between confluent control cells. For instance, these conditions allowed the control cell population, considered at 16 h after the last exposure of their stressed counterparts, to be doubled compared to a cell population considered before the first exposure, 48 h earlier (Fig. 2b): these control cells (Fig. 2b column 2) have proliferated normally as expected when compared to cells prior exposure (Fig. 2b column 1).

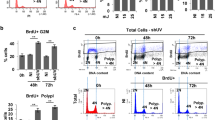
No apoptosis in N-hTERT keratinocytes after 8 exposures to UVB at 300 mJ/cm2. a Schedule of repeated exposures to UVB. N-hTERT keratinocytes are exposed eight times to UVB at a sublethal 300 mJ/cm2 dose (four daily exposures during 2 days). Cells were analyzed at various times after the last exposure. b Viability of N-hTERT keratinocytes after eight UVB exposures. N-hTERT keratinocytes were exposed to UVB at repeated doses of UVB from 200 to 400 mJ/cm2 dose with four daily exposures during 2 days. Control cells (CTL) were submitted to the same conditions without UVB. Cell viability was assessed 16 h after the last exposure. The results from triplicates are expressed as percentages of cell survival compared with viability before (column 1) any exposure (NS: non significant, *** P < 0.001). c1 Detection of active caspase-3 in N-hTERT keratinocytes exposed to a single UVB exposure. N-hTERT keratinocytes were exposed, or not (CTL), once to 300 or 2,400 mJ/cm2 UVB. The activation of caspase-3 was investigated by immunostaining using a specific anti-active caspase-3 antibody. The nuclei were stained with TO-PRO-3. c2 No activation of caspase-3 after eight sublethal UVB exposures. N-hTERT keratinocytes were exposed to UVB at a sublethal 300 mJ/cm2 dose with four daily exposures during 2 days. d Detection of cleaved form of PARP after a single or repeated exposure(s) to UVB. d1 N-hTERT keratinocytes were exposed once to UVB at 300 mJ/cm2 or 2,400 mJ/cm2. Total cell extracts at 4 and 16 h after the exposure were analysed by Western blotting with an anti-PARP-1 antibody. The full length PARP protein (116 kDa) and the fragment resulting from PARP cleavage (85 kDa) are indicated. α-tubulin protein level was used as a reference. d2 N-hTERT keratinocytes were exposed eight times to 300 mJ/cm2 UVB, with four exposures per day for 2 days. Cells treated or not with staurosporine at 500 nM for 4 h were used respectively as positive and negative (CTL−) controls. CTL d1 (day 1) corresponds to control cells before any exposure to UVB
Eight exposures to 300 mJ/cm2 UVB doses inhibit cell proliferation without any sign of lethality when compared to cells analyzed before any exposure to UVB (Fig. 2b). Repeated exposures represent a cumulated dose of 2,400 mJ/cm2 (8 × 300 mJ/cm2) but are not lethal, whereas a significant loss of cell viability is observed above 1,200 mJ/cm2 after a single exposure (Fig. 1a). A significant reduction in cell viability is observed after 8 repeated exposures to 400 mJ/cm2 UVB (Fig. 2b). Using immunodetection of active caspase-3 as a marker of apoptosis, we show that 8 exposures to 300 mJ/cm2 UVB are not pro-apoptotic (Fig. 2c2) in contrast to keratinocytes exposed to a single 2,400 mJ/cm2 UVB dose (Figs. 1a, 2c1). Thus, keratinocytes respond differently after fractionated doses of UVB, as they survive to the treatment. As expected, a single 300 mJ/cm2 UVB dose does not trigger detectable apoptosis during the 64 h following exposure (Fig. 2c1). This was confirmed by analysis of PARP cleavage as another marker of apoptosis (Fig. 2d1, d2).
A 300 mJ/cm2 UVB dose may be considered as relevant. For instance, a single exposure of human skin grafts to 500 mJ/cm2 UVB induces sunburn cells in the suprabasal layers of the epidermal tissue, 24 h after the exposure (Del Bino et al. 2004).
DNA damage and cell cycle arrest after repeated sublethal exposures to UVB
Exposure(s) of N-hTERT keratinocytes to UVB clearly result(s) in the formation of cyclobutane pyrimidine dimers (CPDs) in the nucleus of UVB-exposed keratinocytes (Fig. 3a, b). Keratinocytes exposed to a single 2,400 mJ/cm2 UVB dose undergo severe cell loss (Fig. 3b). CPDs disappear within 24 h after a single 300 mJ/cm2 UVB exposure. In keratinocytes exposed eight times to 300 mJ/cm2 UVB, CPDs remain detectable for at least 64 h (Fig. 3a). These cumulative effects were correlated with the dramatic decrease in cell proliferation still observed 64 h after the eighth exposure to low dose UVB. Indeed a significant 5.3-fold decrease in [3H]-thymidine incorporation was found (Fig. 3c). Keratinocytes exposed to a single 300 mJ/cm2 UVB dose do not display any decrease in DNA synthesis, suggesting that the response observed after repeated exposures is not due solely to the last exposure to UVB. N-hTERT keratinocytes exposed to a single 2,400 mJ/cm2 UVB dose display a dramatic decrease in cell proliferation compared to control cells (Fig. 3c).

Repeated exposures to sublethal UVB doses induce DNA damage and trigger to growth-arrest in N-hTERT keratinocytes. a Repeated exposures to UVB generate DNA damage. N-hTERT keratinocytes were exposed to 300 mJ/cm2 UVB, four times a day during 2 days. Micrographs of CPDs immunofluorescence (green) were obtained by semi-quantitative confocal microscopy. Nuclei were stained with TO-PRO-3 (blue). b CPDs detection after a single exposure to 300 or 2,400 mJ/cm2 UVB in N-hTERT keratinocytes. N-hTERT keratinocytes were exposed to 300 mJ/cm2 or 2,400 mJ/cm2 UVB as single doses. c Large decrease in [3H]-thymidine incorporation after repeated UVB exposures. N-hTERT keratinocytes were exposed either to 300 mJ/cm2 UVB eight times [four daily exposures during 2 days (8×)], or to 2,400 mJ/cm2 UVB as a single exposure (1×). Respective controls were included. The incorporation of [3H]-thymidine into DNA was analysed between 40 and 64 h after the last UVB exposure. The results are expressed as means ± SD of ratios between the counts per min (cpm) of radioactivity incorporated by the cells and the protein content assayed with the Folin method (μg/ml). Student’s t-test, NS: non significant, *** P < 0.001 (Color figure online)
P53 and p21WAF−1 abundance is increased after repeated exposures of N-hTERT keratinocytes to UVB, while SA β-gal is not
Active p53 protein allows DNA repair, apoptosis or cell cycle arrest by transcriptional activation of p53-related genes such as p21WAF−1, an inhibitor of cyclin-dependent kinases that blocks the cell cycle in G1/S (Cuddihy and Bristow 2004). Cell cycle arrest was confirmed by sustained increased abundance of p53 and p21WAF−1 at the protein level in N-hTERT keratinocytes until 64 h after the series of eight exposures to UVB (Fig. 4a) and also at the mRNA level for p21WAF−1 (Fig. 4b). Furthermore, DNA-binding activity of p53 was measured from 45 min to 64 h after the last exposure to UVB. A maximal increase was observed 90 min after the last exposure (2.9-fold increase in UVB exposed cells, Student’s t-test with P < 0.05 vs. non exposed cells).

Increased abundance of p53 and p21WAF−1, increased cell spreading, but not increased SA β-gal activity in N-hTERT keratinocytes repeatedly exposed to UVB. N-hTERT keratinocytes were treated with four daily exposures to 300 mJ/cm2 of UVB during 2 days (UVB). a Repeated exposures to UVB increased the relative protein abundance of p21WAF−1 and p53. Total proteins were extracted at 16, 40 and 64 h after the last exposure to UVB. Western blot was performed to detect p21WAF−1 and p53. α-tubulin protein was used as reference level. Results are representative of three independent experiments. b Relative increase in the steady-state level of the mRNA of p21WAF−1 after eight exposures to UVB in N-hTERT keratinocytes. Total RNA was extracted at 16 and 64 h after the eighth exposure to UVB and real-time RT-PCR was performed for human p21WAF−1. GAPDH was chosen as housekeeping gene. The results (means of triplicates ± SD) are expressed as fold increase found in cells exposed to UVB (UVB) compared with the mRNA abundance of the respective mRNA species in control cells (CTL) (* P < 0.05). c Morphology of N-hTERT keratinocytes after eight UVB exposures. Cells were visualized by phase-contrast microscopy and photographed just before the first exposure and 64 h after the last exposure (40× magnification and further 2.8× magnification in zoom area). d No increase of SA β-gal positive cells after the last UVB exposure of N-hTERT keratinocytes. N-hTERT keratinocytes (KC) were exposed, or not, eight times to UVB at 300 mJ/cm2. Primary keratinocytes exposed similarly to UVB were used as positive control. Histochemical detection of SA β-gal in UVB-exposed and control cells (blue) was assessed 64 h after the last exposure. Cells were visualized by phase-contrast microscopy (40× magnification). e Transient increase of the amount of oxidized proteins of N-hTERT keratinocytes after eight UVB exposures. At 16 and 64 h after the last exposure to UVB, total protein from cells exposed to UVB (UVB) or not (CTL) were extracted and detection of oxidized proteins was performed by oxyblots. Results presented are representative of three independent experiments (Color figure online)
Cells repeatedly exposed to UVB display an enlarged surface when compared to control cells 64 h after the last exposure (Fig. 4c). Such an increased spreading is very often observed in senescent cells (Lewis et al. 2008). However, the more specific marker senescence-associated β-galactosidase (SA β-gal) was not significantly increased 64 h after eight exposures to sublethal UVB doses in N-hTERT keratinocytes (Fig. 4d), in contrast to the observations made with primary keratinocytes which showed a 3.8-fold increase in SA β-gal activity positive-cells (Student’s t-test with P < 0.01 vs non exposed cells). In order to gain in sensitivity, SA β-gal activity in N-hTERT keratinocytes exposed eight times to UVB was evaluated using a quantitative microfluidic method that has been already used in the previous study of stress-induced premature senescence of human fibroblasts, used as positive control herein (Debacq-Chainiaux et al. 2008). A small but not significant increase in SA β-gal activity positive-cells of less than 8% was found in N-hTERT keratinocytes exposed eight times to UVB with respectively 9.4 ± 3 and 17.2 ± 7% of positive cells in CTL and UVB exposed cells (Student’s t-test P-value > 0.05 vs non exposed cells). It is difficult to assess whether such a small difference is biologically relevant.
Oxidized proteins accumulation is commonly seen as a hallmark of cellular aging and results, at least in part, from impaired degradation of abnormal proteins (Friguet 2006). Proteasome is the major proteolytic system implicated in removal of abnormal and oxidatively damaged proteins (Davies 2001). In human keratinocytes, previous studies have evidenced that the proteasome activity and content are decreased during replicative senescence as well as in cells aged in vivo (Petropoulos et al. 2000). Here, oxyblot experiments showed a transient increase in the amount of oxidized proteins at 16 h after the last exposure of keratinocytes to UVB, with return to basal level at 64 h after the last exposure (Fig. 4e). Thus oxidatively damaged proteins were efficiently processed. Furthermore, none of the 3 proteasome activities were modified within 64 h after eight exposures to UVB (data not shown) and no variation in the abundance of the 20S proteasome subunit was observed by Western blot (data not shown).
Lastly, the number of senescence-associated genes is very low if not unexisting in keratinocytes, beyond p21 and p53 which are not specific of senescence in cells able to differentiate. We tested the mRNA abundance of several senescence-associated genes namely fibronectin, osteonectin and clusterin which are overexpressed in senescent fibroblasts (Dumont et al. 2000). Surprisingly, clusterin, osteonectin and fibronectin genes were respectively weakly, moderately and well expressed in N-hTERT keratinocytes as well as in primary keratinocytes not exposed to UVB (data not shown). Secondly, we found that the abundance of mRNA of clusterin tends to decline, although not significantly, in N-hTERT keratinocytes exposed to UVB. Osteonectin and fibronectin mRNA abundance showed no significant difference in this experimental model.
Involucrin and filaggrin abundance is increased after repeated exposures to UVB
Cell cycle arrest and increased expression of p21WAF−1 are common features observed during epidermal differentiation and senescence (Gandarillas 2000). In order to investigate whether epidermal differentiation is triggered by repeated sublethal UVB exposures of N-hTERT keratinocytes, these cells were exposed to repeated exposures of UVB as described above. The involucrin and filaggrin proteins are markers of late epidermal differentiation in keratinocytes, detected in upper spinous (involucrin) and granular layers (involucrin, filaggrin) of the epidermis (Eckert et al. 1997). The mRNA abundance of these two proteins was significantly increased both at 16 and 64 h after the last exposure to UVB (Fig. 5a, b). Furthermore, according to immunofluorescent detection, the abundance of these two proteins was also markedly enhanced in UVB-exposed cells at 64 h after the last exposure to UVB (Fig. 5c).

Increased abundance of involucrin and filaggrin mRNA and proteins and keratins 6, 16 and 17 proteins after eight exposures to UVB in N-hTERT keratinocytes. N-hTERT keratinocytes were treated four times daily with 300 mJ/cm2 UVB during 2 days (UVB). a, b Relative increase in the abundance level of the mRNA of involucrin a and filaggrin b after eight exposures to UVB in N-hTERT keratinocytes. Total RNA was extracted at 16 and 64 h after the eighth exposure to UVB. Real-time RT-PCR was performed. GAPDH was chosen as housekeeping gene. The results (means of triplicates ± SD) are expressed as fold increase found in cells exposed to UVB (UVB) compared with the abundance of the respective mRNA species in control cells (CTL) at 16 h after the last exposure (* P < 0.05 and ** P < 0.01 vs respective control cells). c Increased protein abundance of involucrin and filaggrin at 64 h after eight exposures to UVB exposure in N-hTERT keratinocytes. Cells were labeled with a rabbit anti-involucrin antibody, or with a mouse anti-filaggrin antibody. Nuclei were stained with TO-PRO3. d Repeated exposures to UVB increased the relative protein abundance of the keratins K6, K16 and K17. Total proteins were extracted at 16, 40 and 64 h after the last exposure of N-hTERT keratinocytes to UVB. Western blot was performed to detect K6, K16 and K17. α-tubulin protein was used as reference level. Results are representative of three independent experiments
Searching for more evidence of a differentiation process in N-hTERT keratinocytes repeatedly exposed to UVB, we estimated the abundance of several keratins by Western Blot. First the abundance of K14 and K10 were estimated. K14 is a marker of basal proliferating cells explaining that its elevated abundance in cultured keratinocytes did not change after repeated exposures to UVB. K10, an early marker of differentiation, was not detectable (data not shown). This was expected since the K10 gene is induced in vitro only in confluent keratinocytes (Poumay and Pittelkow 1995). K10 is more expressed in the skin or in reconstructed epidermis than in monolayers of cultured keratinocytes (Gazel et al. 2003). The expression of K6, K16 and K17 coincides with alterations in activated keratinocytes and is interpreted as a manifestation of an alternative programme of differentiation (Bousquet and Coulombe 2002; Freedberg et al. 2001; Wong and Coulombe 2003). The expression of these keratins is also known to be induced after exposure of human skin to UVB (Del Bino et al. 2004). By Western blot analysis, we showed that the abundance of these keratins was increased in keratinocytes repeatedly exposed to UVB (Fig. 5d).
Discussion
In keratinocytes exposed to UVB, p53 and subsequently p21WAF−1 play a central role in mediating cell cycle arrest, either allowing DNA repair or leading to apoptosis (Decraene et al. 2005). The senescence response is maintained by p53, while p16INK4a provides a second barrier against the unlimited growth of human cells (Beausejour et al. 2003). P53 and p21WAF−1 protein abundance was increased in N-hTERT keratinocytes exposed 8 times to low dose UVB, suggesting that N-hTERT keratinocytes retain primary actors involved in cell cycle arrest of keratinocytes exposed to UVB. We found neither apoptosis nor any significant increase in SA β-gal activity in keratinocytes lacking p16INK-4a, expressing hTERT and repeatedly exposed to UVB. Proteasome is downregulated during replicative senescence of keratinocytes as well as in cells aged in vivo, possibly resulting in the accumulation of modified proteins (Petropoulos et al. 2000). Repeated exposures of N-hTERT keratinocytes to subcytotoxic doses of UVB trigger a severe, but transient, oxidative stress without impairment of proteasome function. Another study showed a decrease of the proteasome activities in primary keratinocytes upon single UVA/UVB exposure (Bulteau et al. 2002).
This work suggests that p16INK-4a is involved in UVB-induced senescence. Restoration of the expression of p16INK-4a in N-hTERT keratinocytes would be interesting in order to confirm the importance of p16INK-4a in the induction of senescence. However, the molecular event responsible for the complete loss of p16INK-4a expression in N-hTERT keratinocytes has not been elucidated yet in this unique cell line despite many efforts. Indeed, neither deletion of DNA sequence or CpG island methylation of the promoter was detected (Dickson et al. 2000). The expression of p16INK-4a is only detected in the granular layer of normal skin (Nakamura and Nishioka 2003) which is made of non proliferative cells. Ectopically expressing p16INK-4a in this proliferating cell line is technically difficult for several reasons and may not give any interpretable results. Only inducible/regulated expression of p16INK-4a can be envisaged since uncontroled overexpression of p16INK-4a will block proliferation and prevent any further experiments aimed at testing whether p16INK-4a controls senescence after exposure to UVB. One should use the native promotor of the gene plus its known regulatory sequences which represents a very long DNA sequence. This may not give any interpretable results since interactions will still be possible with the unknown mechanisms responsible for the lack of expression of p16INK-4a gene in the wild cell line.
Expression of hTERT alone does not prevent stress-induced senescence induced by either acute sublethal short exposures to H2O2, UV, UVB or gamma-irradiation (de Magalhaes et al. 2002; Gorbunova et al. 2002; Matuoka and Chen 2002). Telomerase expression alone cannot immortalize human keratinocytes. Gain of function due to concomitant absence of p16INK-4a and expression of hTERT are still possible. This question will remain a debate for long since p16INK-4a-deficient human keratinocytes are not immortal, which does not make them suitable for deep functional studies of stress response.
One very interesting point in the present study is that, in addition to overexpression of p53 and p21WAF−1, we observed an increased expression of involucrin and filaggrin in keratinocytes repeatedly exposed to UVB, suggesting that a differentiation state is induced rather than senescence, and that differentiation and senescence can be uncoupled in this model. Upregulation of involucrin and filaggrin has been found in vivo after exposure to UVB (Lee et al. 2002). Filaggrin expression reflects maturity in the level of epidermal differentiation (Lee et al. 2002). In vivo, an increase in epidermal thickness, mostly of the cornified layer, occurs after UV exposure and provides protection from further UV-induced damage since UVB need to cross an thickened protective layer before they can reach and affect proliferative basal keratinocytes (de Winter et al. 2001). Subsequently, epidermal differentiation will eventually lead to the elimination of damaged cells (Gandarillas 2000). Thus, our in vitro findings are consistent with in vivo studies that indicate alterations in the program of keratinocyte differentiation leading to abnormal parakeratosis a few days after UVB exposure (Del Bino et al. 2004; Lee et al. 2002). This reinforces the interest in this experimental model.
K6, K6 and K17 were already known to be induced after exposure of human skin to UVB (Del Bino et al. 2004). K17 is induced by interferon-γ (IFNγ) in keratinocytes (Jiang et al. 1994), during skin inflammation linked with psoriasis (Carlen et al. 2005), in skin allergy (Jiang et al. 1994; Komine et al. 1996) and in damaged epidermis (Bousquet and Coulombe 2002). This suggests that K17 expression may be specific of inflammatory skin conditions that might be reproduced in vitro in this model. K6 is induced in several skin diseases including abnormal differentiation (Stoler et al. 1988; Weiss et al. 1984) or hyperproliferative stratified epithelium such as in psoriasis (Carlen et al. 2005). K6 also participates in migration and wound healing processes (Paladini et al. 1996; Wong and Coulombe 2003). It was also shown in vivo that up-regulation of K16 leads to acanthosis and hyperkeratosis (Takahashi et al. 1994), suggesting that K16 may also act as a protective mechanism through epidermal thickening. Together with involucrin and filaggrin, the increased abundance of these keratins reflects more an alternative differentiation process than a typical differentiation like observed in terminal differentiation process. The expression of K6 and K16 may correspond to a modification of the terminal programme of differentiation after chronic UVB exposures (Sano et al. 2009). In a transcriptomic analysis, Li et al. (2001) have shown that the genes most strongly induced by UVB in primary keratinocytes are components of the cornified envelope, but not other earlier differentiation markers. This particular differentiation process leads to enhancement of the cornified, dead, protective layer of skin.
Berge et al. (2008) have shown that several inducers of the keratinocyte differentiation could have beneficial and anti-aging effect in vitro. The functional role of differentiation in the protection of the epidermis against the harmful effects of UVB suggests that maintaining the ability of keratinocytes to differentiate is crucial for the well-being of this tissue.
The effects of a single sublethal exposure to UVB are rapidly reversed (Del Bino et al. 2004). Therefore, this model based on multiple daily exposures to UVB is more likely to mimic the cumulative alterations due to repeated exposures to UVB. Human keratinocytes exposed to a single 2,400 mJ/cm2 UVB dose were apoptotic, whereas keratinocytes exposed to the same but fractionated UVB dose proceeded toward a state of differentiation that favours survival and resistance against any further stress. Altogether, our results suggest to carefully evaluate the in vitro models based on a single exposure of epidermal components to elevated UVB doses. Apparently, repeated sublethal exposures of keratinocytes to UVB can be responsible for sustained DNA damage including the formation of CPDs that will eventually predispose to skin cancer. Interestingly, Sano et al. (2009) have demonstrated that, in the epidermis of mice, alterations of K6 and K16 expression induced by the chronic UVB exposure were distinct from those elicited by a single acute UVB exposure. Indeed, the expression of K6 and K16 was quite long-lasting after 9 weeks of chronic UVB exposure. In contrast, K6 and K16 expression induced by a single exposure to UVB almost ceased within 2 weeks after that exposure. The long-lasting expression of K6 and K16 also suggest modifications of the differentiation state. This strengthens the interest of studying the effect of repeated exposures to UVB.
Lastly, a further advantage of this model is in the possibility to perform functional studies with this cell type which enlarges greatly its scientific value.
Abbreviations
- CPDs:
-
Cyclobutane pyrimidine dimers
- K:
-
Cytokeratin
- ROS:
-
Reactive oxygen species
- SA β-gal:
-
Senescence-associated β-galactosidase activity
- UVB:
-
Ultraviolet B
References
Beausejour CM, Krtolica A, Galimi F, Narita M, Lowe SW, Yaswen P, Campisi J (2003) Reversal of human cellular senescence: roles of the p53 and p16 pathways. EMBO J 22:4212–4222
Ben-Porath I, Weinberg RA (2005) The signals and pathways activating cellular senescence. Int J Biochem Cell Biol 37:961–976
Berge U, Kristensen P, Rattan SI (2008) Hormetic modulation of differentiation of normal human epidermal keratinocytes undergoing replicative senescence in vitro. Exp Gerontol 43:658–662
Blackburn EH (2005) Telomeres and telomerase: their mechanisms of action and the effects of altering their functions. FEBS Lett 579:859–862
Bousquet O, Coulombe PA (2002) Les kératines: un autre regard sur la biologie de la peau. Medecine/Sciences 18:45–54
Bulteau AL, Moreau M, Nizard C, Friguet B (2002) Impairment of proteasome function upon UVA- and UVB-irradiation of human keratinocytes. Free Radic Biol Med 32:1157–1170
Carlen LM, Sanchez F, Bergman AC, Becker S, Hirschberg D, Franzen B, Coffey J, Jornvall H, Auer G, Alaiya AA, Stahle M (2005) Proteome analysis of skin distinguishes acute guttate from chronic plaque psoriasis. J Invest Dermatol 124:63–69
Cuddihy AR, Bristow RG (2004) The p53 protein family and radiation sensitivity: yes or no? Cancer Metastasis Rev 23:237–257
Davies KJ (2001) Degradation of oxidized proteins by the 20S proteasome. Biochimie 83:301–310
de Magalhaes JP, Chainiaux F, Remacle J, Toussaint O (2002) Stress-induced premature senescence in BJ and hTERT-BJ1 human foreskin fibroblasts. FEBS Lett 523:157–162
de Winter S, Vink AA, Roza L, Pavel S (2001) Solar-simulated skin adaptation and its effect on subsequent UV-induced epidermal DNA damage. J Invest Dermatol 117:678–682
Debacq-Chainiaux F, Pascal T, Boilan E, Bastin C, Bauwens E, Toussaint O (2008) Screening of senescence-associated genes with specific DNA array reveals the role of IGFBP-3 in premature senescence of human diploid fibroblasts. Free Radic Biol Med 44:1817–1832
Decraene D, Smaers K, Maes D, Matsui M, Declercq L, Garmyn M (2005) A low UVB dose, with the potential to trigger a protective p53-dependent gene program, increases the resilience of keratinocytes against future UVB insults. J Invest Dermatol 125:1026–1031
Del Bino S, Vioux C, Rossio-Pasquier P, Jomard A, Demarchez M, Asselineau D, Bernerd F (2004) Ultraviolet B induces hyperproliferation and modification of epidermal differentiation in normal human skin grafted on to nude mice. Br J Dermatol 150:658–667
Dickson MA, Hahn WC, Ino Y, Ronfard V, Wu JY, Weinberg RA, Louis DN, Li FP, Rheinwald JG (2000) Human keratinocytes that express hTERT and also bypass a p16(INK4a)-enforced mechanism that limits life span become immortal yet retain normal growth and differentiation characteristics. Mol Cell Biol 20:1436–1447
Dierick J-F, Frippiat Salmon, Chainiaux Toussaint (2003) Cells, stress and tissue ageing. In: Osiewacz HD (ed) Modulating aging and longevity. Kluwer, Amsterdam, pp 101–125
Dimri GP, Lee X, Basile G, Acosta M, Scott G, Roskelley C, Medrano EE, Linskens M, Rubelj I, Pereira-Smith O et al (1995) A biomarker that identifies senescent human cells in culture and in aging skin in vivo. Proc Natl Acad Sci U S A 92:9363–9367
Dumont P, Burton M, Chen QM, Gonos ES, Frippiat C, Mazarati JB, Eliaers F, Remacle J, Toussaint O (2000) Induction of replicative senescence biomarkers by sublethal oxidative stresses in normal human fibroblast. Free Radic Biol Med 28:361–373
Dumont P, Royer V, Pascal T, Dierick JF, Chainiaux F, Frippiat C, de Magalhaes JP, Eliaers F, Remacle J, Toussaint O (2001) Growth kinetics rather than stress accelerate telomere shortening in cultures of human diploid fibroblasts in oxidative stress-induced premature senescence. FEBS Lett 502:109–112
Eckert RL, Crish JF, Robinson NA (1997) The epidermal keratinocyte as a model for the study of gene regulation and cell differentiation. Physiol Rev 77:397–424
Freedberg IM, Tomic-Canic M, Komine M, Blumenberg M (2001) Keratins and the keratinocyte activation cycle. J Invest Dermatol 116:633–640
Friguet B (2006) Oxidized protein degradation and repair in ageing and oxidative stress. FEBS Lett 580:2910–2916
Gandarillas A (2000) Epidermal differentiation, apoptosis, and senescence: common pathways? Exp Gerontol 35:53–62
Gazel A, Ramphal P, Rosdy M, De Wever B, Tornier C, Hosein N, Lee B, Tomic-Canic M, Blumenberg M (2003) Transcriptional profiling of epidermal keratinocytes: comparison of genes expressed in skin, cultured keratinocytes, and reconstituted epidermis, using large DNA microarrays. J Invest Dermatol 121:1459–1468
Gorbunova V, Seluanov A, Pereira-Smith OM (2002) Expression of human telomerase (hTERT) does not prevent stress-induced senescence in normal human fibroblasts but protects the cells from stress-induced apoptosis and necrosis. J Biol Chem 277:38540–38549
Hayflick L, Moorhead PS (1961) The serial cultivation of human diploid cell strains. Exp Cell Res 25:585–621
Ichihashi M, Ueda M, Budiyanto A, Bito T, Oka M, Fukunaga M, Tsuru K, Horikawa T (2003) UV-induced skin damage. Toxicology 189:21–39
Jiang CK, Flanagan S, Ohtsuki M, Shuai K, Freedberg IM, Blumenberg M (1994) Disease-activated transcription factor: allergic reactions in human skin cause nuclear translocation of STAT-91 and induce synthesis of keratin K17. Mol Cell Biol 14:4759–4769
Komine M, Freedberg IM, Blumenberg M (1996) Regulation of epidermal expression of keratin K17 in inflammatory skin diseases. J Invest Dermatol 107:569–575
Lee JH, An HT, Chung JH, Kim KH, Eun HC, Cho KH (2002) Acute effects of UVB radiation on the proliferation and differentiation of keratinocytes. Photodermatol Photoimmunol Photomed 18:253–261
Lewis DA, Yi Q, Travers JB, Spandau DF (2008) UVB-induced senescence in human keratinocytes requires a functional insulin-like growth factor-1 receptor and p53. Mol Biol Cell 19:1346–1353
Li D, Turi TG, Schuck A, Freedberg IM, Khitrov G, Blumenberg M (2001) Rays and arrays: the transcriptional program in the response of human epidermal keratinocytes to UVB illumination. FASEB J 15:2533–2535
Matuoka K, Chen KY (2002) Telomerase positive human diploid fibroblasts are resistant to replicative senescence but not premature senescence induced by chemical reagents. Biogerontology 3:365–372
Mosmann T (1983) Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods 65:55–63
Nakamura S, Nishioka K (2003) Enhanced expression of p16 in seborrhoeic keratosis; a lesion of accumulated senescent epidermal cells in G1 arrest. Br J Dermatol 149:560–565
Paladini RD, Takahashi K, Bravo NS, Coulombe PA (1996) Onset of re-epithelialization after skin injury correlates with a reorganization of keratin filaments in wound edge keratinocytes: defining a potential role for keratin 16. J Cell Biol 132:381–397
Petropoulos I, Conconi M, Wang X, Hoenel B, Bregegere F, Milner Y, Friguet B (2000) Increase of oxidatively modified protein is associated with a decrease of proteasome activity and content in aging epidermal cells. J Gerontol A Biol Sci Med Sci 55:B220–B227
Poumay Y, Pittelkow MR (1995) Cell density and culture factors regulate keratinocyte commitment to differentiation and expression of suprabasal K1/K10 keratins. J Invest Dermatol 104:271–276
Rabe JH, Mamelak AJ, McElgunn PJ, Morison WL, Sauder DN (2006) Photoaging: mechanisms and repair. J Am Acad Dermatol 55:1–19
Ramirez RD, Morales CP, Herbert BS, Rohde JM, Passons C, Shay JW, Wright WE (2001) Putative telomere-independent mechanisms of replicative aging reflect inadequate growth conditions. Genes Dev 15:398–403
Ressler S, Bartkova J, Niederegger H, Bartek J, Scharffetter-Kochanek K, Jansen-Durr P, Wlaschek M (2006) p16INK4A is a robust in vivo biomarker of cellular aging in human skin. Aging Cell 5:379–389
Rittie L, Fisher GJ (2002) UV-light-induced signal cascades and skin aging. Ageing Res Rev 1:705–720
Sano T, Kume T, Fujimura T, Kawada H, Higuchi K, Iwamura M, Hotta M, Kitahara T, Takema Y (2009) Long-term alteration in the expression of keratins 6 and 16 in the epidermis of mice after chronic UVB exposure. Arch Dermatol Res 301:227–237
Schmitt CA (2007) Cellular senescence and cancer treatment. Biochim Biophys Acta 1775:5–20
Stoler A, Kopan R, Duvic M, Fuchs E (1988) Use of monospecific antisera and cRNA probes to localize the major changes in keratin expression during normal and abnormal epidermal differentiation. J Cell Biol 107:427–446
Takahashi K, Folmer J, Coulombe PA (1994) Increased expression of keratin 16 causes anomalies in cytoarchitecture and keratinization in transgenic mouse skin. J Cell Biol 127:505–520
Toussaint O, Remacle J, Dierick JF, Pascal T, Frippiat C, Royer V, Magalhacs JP, Zdanov S, Chainiaux F (2002) Stress-induced premature senescence: from biomarkers to likeliness of in vivo occurrence. Biogerontology 3:13–17
Weiss RA, Eichner R, Sun TT (1984) Monoclonal antibody analysis of keratin expression in epidermal diseases: a 48- and 56-kdalton keratin as molecular markers for hyperproliferative keratinocytes. J Cell Biol 98:1397–1406
Wong P, Coulombe PA (2003) Loss of keratin 6 (K6) proteins reveals a function for intermediate filaments during wound repair. J Cell Biol 163:327–337
Acknowledgments
VV is Recipient of a FRIA fellowship. FDC and OT are respectively Post-Doctoral Researcher and Research Associate of the Belgian F·N.R.S. This work received financial support through FRFC Grant 2.4506.01 (Belgium) to YP and OT. We acknowledge the Région Wallonne/FSE for the First-Europe project ‘CosmUV’, the First-DEI project ‘Cosmet-X’, and the ‘Réseaux II Senegene’ and ‘Nanotoxico’ projects. We also thank the European Commission for the Integrated Projects ‘Proteomage’ (LSHM-CT-2005-518230); Coordination Action Link-Age (LSHM-CT-2005-513866); and ‘Matiss’ Marie Curie Project (MTKI-CT-2006–042768).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Bertrand-Vallery, V., Boilan, E., Ninane, N. et al. Repeated exposures to UVB induce differentiation rather than senescence of human keratinocytes lacking p16INK-4A . Biogerontology 11, 167–181 (2010). https://doi.org/10.1007/s10522-009-9238-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10522-009-9238-y