
Abstract
We isolated highly purified CD8+CD28+ and CD8+CD28− T cell populations from healthy young and elderly persons for gene expression profiling using Affymetrix oligonucleotide microarrays. We demonstrate that the gene expression profile of CD8+CD28− T cells is very similar in young and elderly persons. In contrast, CD8+CD28+ in elderly differ from CD8+CD28+ in young persons. Hierarchical clustering revealed that CD8+CD28+ in elderly are located between CD8+CD28+ in young and CD8+CD28− (young and old) T cells regarding their differentiation state. Our study demonstrates a dichotomy of gene expression levels between CD8+CD28+ T cells in young and elderly persons but a similarity between CD8+CD28− T cells in young and elderly persons. As CD8+CD28+ T cells from elderly and young persons are distinct due to a different composition of the population, these results suggest that the gene expression profile does not depend on chronological age but depends on the differentiation state of the individual cell types.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Aging is universal but the precise mechanisms linking the aging immune system to diseases in the elderly are poorly understood. Recent advances in the study of global patterns of gene expression with the use of microarray technology, coupled with extensive data analysis using bioinformatic tools, have provided new insights into the mechanism of the aging of the immune system. This method can provide a wealth of information at the level of gene expression and is a powerful method to identify genes and pathways involved in complex processes.
Aging has often been associated with a ‘decline’ in immune function. The immune system undergoes dramatic restructuring with age, leading to a decline in immune responses and an increased vulnerability of old individuals to infectious diseases. T cell immunosenescense has traditionally been associated with thymic involution, since a striking decline in the output of new thymus-derived T cells occurs with age (George and Ritter 1996). However, absolute T cell numbers remain comparable between young and old individuals (Stulnig et al. 1995), requiring new T cells to be generated extra-thymically or by clonal expansion of pre-existing T cells. One intriguing change observed in the T cell pool with aging is the marked increase in the proportion of CD8+ T cells lacking expression of the CD28 surface receptor (Fagnoni et al. 1996), a major costimulatory molecule required for functional T cell activation (Sperling and Bluestone 1996). The loss of CD28 expression on T cells is the most consistent biological indicator of aging in the human immune system, and the frequency of CD28− T cells is a key predictor of immunocompetence in the elderly. CD8+CD28− T cells are often oligoclonal in nature and do not proliferate well in response to antigenic stimulation (Posnett et al. 1994; Sansoni et al. 1997). These clonally expanded T cells can persist in humans for years (Schwab et al. 1997). CD8+CD28− T cells occur in virtually all healthy elderly subjects (Posnett et al. 1994), and an elevated number of this T cell subset is associated with numerous autoimmune conditions such as systemic lupus erythematosus (Kaneko et al. 1996) and rheumatoid arthritis (Fitzgerald et al. 1995). Additionally, persistent antigenic stimulation such as CMV infection drives the accumulation of CD8+CD28− T cells.
It was therefore of interest to analyze the gene expression profile of CD8+CD28+ and CD8+CD28− T cell subsets from young and elderly persons.
Material and methods
Purification of CD8+CD28+ and CD8+CD28− T cells
Peripheral blood was obtained from four young and four elderly healthy volunteers (Table 1). The health status of these persons was defined by the absence of severe diseases (diabetes, rheumatoid arthritis, dementia and cancer), acute infectious diseases and drugs which are known to interfere with the immune system. All participants had given their written informed consent and blood collection was approved by the Ethics Committee of Innsbruck Medical University. Peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll-Hypaque density gradient centrifugation (Amersham Pharmacia Biotech AB, Uppsala, Sweden). CD8+CD28+ and CD8+CD28− T cells were enriched from PBMCs in a series of separations using magnetic beads. To obtain sufficiently high subset numbers for analysis, purifications were only performed in CMV-seropositive persons who had high numbers of CD8+CD28− cells. PBMCs were depleted from non-T cells by application of a Pan T cell Isolation Kit II (Miltenyi Biotec, Bergisch Gladbach, Germany). CD8+ T cells were then positively selected using CD8 Multisort Microbeads (Miltenyi Biotec). After enzymatic removal of the CD8 Multisort Microbeads, CD8+ T cells were stained with an allophycocyanin-conjugated mAb recognizing CD28 (BD Pharmingen) and CD28+ cells were obtained by positive selection using anti-allophycocyanin microbeads and a LS column (Miltenyi Biotec). The purity of the obtained CD8+CD28+ population was more than 90%. The CD28− fraction was depleted of residual CD28+ cells applying an additional LD column (Miltenyi Biotec) resulting in a purity of more than 92%.
Affymetrix microarray experiment
Total RNA was extracted using TRI reagent (Sigma-Aldrich, Vienna, Austria) from freshly purified CD8+CD28+ and CD8+CD28− T cells. Glycogen (Roche) was added as a carrier for RNA precipitation at a concentration of 1 μg/ml. As a minimum amount of 1.5 μg RNA for microarray analysis was needed, a two-cycle target labeling protocol was used in our case to amplify total RNA (Affymetrix). Hybridization was done onto high-density oligonucleotide human genome array GeneChips U133 Plus 2.0 (Affymetrix). This chip comprises more than 54,000 probe sets and analyzes the expression level of over 47,000 transcripts and variants including 38,500 well-characterized human genes. The arrays were scanned at 3 μm resolution using a GeneChip® Instrument System. Affymetrix gene chip hybridization experiments were outsourced to the Microarray Facility Tuebingen, Germany, an authorized Affymetrix Service Provider. Both, experimental and data analysis workflow were fully compliant with the MIAME 2.0 Standard. Gene Chip Annotation Data for U133 Plus 2.0 Array were supplied by Affymetrix Inc.
Microarray data analysis
CARMAweb (Comprehensive R-based Microarray Analysis web service) was used for data preprocessing (background correction, quality control and normalization) (Rainer et al. 2006). Affymetrix microarray suite version 5 (MAS5) algorithm was applied for probe-level normalization and background correction. The coefficient of variability of CD8+CD28+ T cells between the two young persons and between the two old persons was 11 and 14%, respectively, while the coefficient of variability of CD8+CD28− T cells was 17 and 15%, respectively. To minimize the negative effects of random noise, filtering steps were applied. Genes that were consistently absent were excluded from analysis. Analyses were done using TM4 MeV software (Saeed et al. 2003). The resulting genes were then analyzed on the basis of their biological process, function or pathway from collectedly protein database by Panther classification system (Mi et al. 2005).
Validation of microarray results by using quantitative RT-PCR
cDNA first strand synthesis was reverse transcribed from 1 μg total RNA preparation using Reverse Transcription System (Promega) and oligo dT15 primers. Several genes that indicated a remarkable difference between subsets were selected for validation by means of quantitative RT-PCR with the LightCycler® 480 System (Roche Diagnostics, Basel, Switzerland). cDNA primers for amplification of target genes were designed using Primer3 software (Rozen and Skaletsky 2000); (Table 2). PCR was performed in a total volume of 22 μl with primers at a concentration of 2.7 μM. cDNA template of 2.5 × 10−2 μg total RNA (5 μl) and 11 μl 2× SYBR Green 1 Master (Roche Diagnostics) for 50 cycles. cDNA from 3 young and 3 elderly persons were checked to ensure consistency of the results. mRNA expression was normalized to GAPDH.
Results
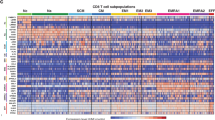
We used highly purified CD8+CD28+ and CD8+CD28− T cell populations from healthy young and elderly persons for gene expression profiling using Affymetrix oligonucleotide microarrays. Hierarchical clustering analysis was used to explore the relationship between CD8+ T cell subsets in different age groups. Firstly, we identified differentially expressed genes between CD8+CD28+ and CD8+CD28− in young and elderly persons. For this purpose, one-way ANOVA test with P > 0.05 was used and samples were divided into four groups based on different subsets and age groups (CD8+CD28+ young, CD8+CD28+ old, CD8+CD28− young, and CD8+CD28− old). A set of 4964 probe IDs corresponding to 4115 genes were identified to be differently expressed between the groups. Unsupervised hierarchical clustering revealed three distinct groups, namely CD8+CD28+ young, CD8+CD28+ elderly and CD8+CD28− young and elderly. The elderly CD8+CD28+ group is clustered between the CD8+CD28+ young and CD8+CD28− young and elderly subsets. Our analysis thus reveals that CD8+CD28− from young and elderly persons are clustered together (Fig. 1).

(a) Hierarchical clustering (Eisen et al. 1998) of CD8+CD28+ (28P) and CD8+CD28− T cells (28N) from two young (Y) and two elderly persons (O). A set of 4964 probe IDs corresponding to 4115 genes were identified as differentially expressed between CD8+ T cell subsets and age groups using one-way ANOVA test. Each row in the heat map represents a gene and each column represents a CD8+ T cell subset of a different donor. A red color indicates higher than median expression (up-regulation) and green indicates lower than median expression (down-regulation). The expression values are calculated in log 2 scale. A hierarchical clustering with complete linkage clustering model using Euclidean distance was performed that results in three different groups based on T cell subsets and age. This clustering also revealed that genes were grouped into 21 clusters based on the similarity on pattern and expression value. (b) A higher magnification of linkage distance showing three distinct groups, CD8+CD28+ from young, CD8+CD28+ from elderly and CD8+CD28− from both, young and elderly. (c) As an example, cluster 13 includes genes that are higher expressed in CD8+CD28+ T cells from elderly persons compared to all other T cell subsets
The hierarchical clustering also revealed that genes were clustered in 21 different patterns. Genes in cluster 3, 5, 7, 17 and 19 appear to represent genes that are typically expressed at a high level in young CD8+CD28+ T cells. Genes which are highly expressed in elderly CD8+CD28+ are represented in cluster 13, while clusters 2, 4, 6, 8, 9, 11, 14, 20 and 21 are marked for effector signature (Fig. 1a). This hierarchical clustering shows that the gene expression profile of CD8+CD28+ in elderly persons was closer to both CD8+CD28− in young and elderly persons than to the CD8+CD28+ T cells from young persons (Fig. 1a, b). This analysis also suggests that the CD8+CD28− T cell subsets from young and elderly persons have a similar gene expression profile. We also confirmed that our Affymetrix data are in accordance with the result obtained from RT-PCR as shown in some randomly selected genes in Fig. 2. For instance, granzymes (GZMB and GZMH) are typically higher expressed in CD8+CD28− T cells whereas CD28, CCR7 and MAL expression is high in CD8+CD28+ T cells, which is in accordance with the literature (Fann et al. 2005).

Differences in the gene expression profile between CD8+ T cell subsets from young and elderly persons analyzed by microarray (a) and quantitative RT-PCR (b). Bars represent mean gene expression level as percentage ± SEM relative to the CD8+CD28+ T cell population from young which was considered basal level. CD8+CD28+ T cells from elderly persons (grey), CD8+CD28− from young (black) and elderly persons (white). Abbreviations: GZMB, Granzyme B; GZMH, Granzyme H; ITGAM, integrin αM; CCR7, chemokine (C-C motif) receptor 7; MAL, T cell differentiation protein
Gene expression profile of CD8+CD28+ T cells from young persons
A list of 4115 genes which was found to be differentially expressed between CD8+ T cell subsets was used for further analysis. Pavlidis template matching method (PTM) (Pavlidis and Noble 2001) with P < 0.01 and 2-fold difference was performed to identify genes that are highly expressed in CD8+CD28+ cells from young persons compared to other subsets. 409 genes met these criteria. Later, we identified the biological processes in which the genes were involved, their molecular function or related pathways using panther classification category. Genes responsible for biological processes such as cell proliferation and differentiation as well as cell adhesion mediated signaling are overrepresentated in CD8+CD28+ T cells from young persons. CD8+CD28+ T cells subsets from young persons express PIK3CD, MAL and IL6R which play a role in cell growth and differentiation. But also lymph node homing markers such as CD62L (SELL) and CCR7 as well as other adhesion molecules (e.g. PECAM1, DCHS1, F11R, FN1 and NRCAM) were highly expressed. A complete list of genes with a higher expression in CD8+CD28+ T cells from young persons compared to other groups is shown in Table 3.
Gene expression profile of CD8+CD28+ T cells from elderly persons
The same procedure was applied to analyze the gene expression of CD8+CD28+ T cells from elderly persons. 96 genes were found to be highly expressed in CD8+CD28+ T cells from elderly persons compared to CD8+CD28+ T cells young and CD8+CD28− T cells (young and elderly). Genes with a typically high expression included GATA3, BIRC3, FAS, RGS1, and MAP3K1. Genes involved in inflammation mediated by cytokine and chemokine signaling pathway, induction of apoptosis, and signal transduction were IFNGR1, FAS and TNFRSF18 (Table 4).
Gene expression profile of CD8+CD28− T cells
CD8+CD28− T cells from young and elderly persons do not greatly differ in their gene expression profile. Genes encoding lytic granule proteins like granzyme B (GZMB), granzyme H (GZMH), perforin (PRF1), granulin (GRN) are highly expressed in CD8+CD28− T cells. We could also confirm previous findings that killer cell lectin-like rectors and killer Ig-like receptor family members such as KLRD1, KLRF1, KLRC3, KIR2DL5A, KIR3DL2 and KIR2DS2 were highly expressed in CD8+CD28− T cells. Additionally, we observed a high expression of genes playing a role in signal transduction in receptor-mediated cell-adhesion e.g. ITGAL and ITGAM, and in transcription e.g. GAS7 (Table 5).
Discussion
We demonstrate that the gene expression profile of CD8+CD28− T cells is very similar between young and elderly persons, while CD8+CD28+ in elderly persons have a gene expression profile different compared to CD8+CD28+ T cells from young persons. Hierarchical clustering also showed that CD8+CD28+ T cells in the elderly are clustered between CD8+CD28+ from young and CD8+CD28− T cells from young and elderly persons. Thus, three different clusters were produced by this analysis i.e. young CD8+CD28+, elderly CD8+CD28+ and CD8+CD28− from both young and elderly. This overall clustering classifies genes into 21 different sub-cluster based on expression level and pattern (Fig. 1).
As about half of the CD8+CD28+ T cell population in young persons is considered antigen-inexperienced or naïve, it was interesting to have a closer look at the genes that are typically highly expressed in this subsets. Genes responsible for T cell activation, cell proliferation and differentiation as well as adhesion molecules are highly expressed in young persons. For example, PIK3CD a member of the phosphoinositide 3-kinases (PI3Ks) family regulates numerous biological processes including cell growth, differentiation, survival, proliferation, migration and metabolism. It has been shown that PI3K activation during the first nine hours of T cell stimulation is essential for T cell proliferation (Okkenhaug and Vanhaesebroeck 2003). We also observed high expression of a hydrophobic protein called MAL in CD8+CD28+ T cells from young persons which is associated with human T cell differentiation (Alonso and Weissman 1987).
Another gene that may play an important role is the interleukin 6 receptor complex (IL6R and IL6ST). The IL6 receptor is a protein complex consisting of an interleukin 6 signal transducer (IL6ST/GP130/IL6-beta), a receptor subunit shared by many other cytokines. Dysregulation of the production of IL6 (a potent pleiotropic cytokine that regulates cell growth and differentiation and plays an important role in the immune response) and the receptor are implicated in the pathogenesis of many diseases, such as multiple myeloma, autoimmune diseases and cancer (Alexandrakis et al. 2003; Holub et al. 2002).
The CD8+CD28+ T cells in elderly persons over-express genes associated with the induction of apoptosis such as FAS and TNFRSF18. FAS is a member of the TNF-receptor superfamily and contains a death domain. FAS has been shown to play a central role in the physiological regulation of programmed cell death, and has been implicated in the pathogenesis of various malignancies and diseases of the immune system (Hiraide et al. 2005; Radfar et al. 2005).
It is widely accepted that cell-mediated immune functions decline with age, rendering an individual more susceptible to infection and possibly cancer, as well as to age-associated autoimmune diseases. The exact causes of the decline in T cell functions are not known. One possible cause could be the development of defects in the transduction of mitogenic signals following TCR stimulation. Signal transduction genes such as EPHA4, TIAM1 and RGS1 and cell structure and metabolism such as ANK3 are also upregulated in CD8+CD28+ cells from elderly persons. It has been shown that EPHA4 is highly expressed in lesional skin biopsy specimens in patients with cutaneous T cell lymphomas related to Sezary syndrome (Sz), a malignancy of CD4+ memory skin-homing T cells. The patients have erythroderma, lymphadenopathy, and peripheral blood involvement (van Doorn et al. 2004). Another signaling protein TIAM1 contributes to the invasion and metastasis of the human giant-cell lung carcinoma cells (Hou et al. 2004).
In accordance with previous studies, CD8+CD28− T cells express several different NK cell receptors (KIR) and produce high levels of granzyme, perforin and granulin, indicating an increased cytotoxic capacity (Fann et al. 2005). The gain of NK receptor expression on CD8+CD28− T cells may facilitate their effector functions as compensation for impaired proliferation (Tarazona et al. 2000).
In contrast to CD8+CD28+ T cells, CD8+CD28− T cells are very similar between young and elderly persons. Only few genes were found to be differentially expressed such as TNFRSF1A, a gene that plays a role in the apoptosis signaling pathway and CD274, a ligand of the programmed death (PD)-1 which is associated with T cell exhaustion and disease progression in HIV-specific cells (Day et al. 2006).
A critical question raised by these results is whether the observed differences between CD8+CD28+ in young and elderly persons reflect age-related differences in gene expression at the level of the single cell. It has been shown that CD8+CD28+ T cells are functionally heterogeneous and may contain different cell types. Using additional surface markers, this subset is for instance known to contain CD28+CD45RA+ and CD28+CD45RA− T cells (Nociari et al. 1999). Furthermore, the CD28+CD45RA− subset contains a subpopulation which expresses high levels of CD62L+ and CCR7+, and is referred to as central memory T cells (TCM) (Wills et al. 2002; Sallusto et al. 1999).
Recently we described that CD45RA+CD28+ T cells—a phenotype generally considered as naïve cells—from old persons differ from their young counterparts (Pfister et al. 2006). CD45RA+CD28+ T cells from elderly individuals have significant shorter telomeres and a restricted TCR repertoire in almost all 24 Vβ families. Cells from elderly persons also have a low expression of CD62L and CCR7. Additionally, they display an effector-like cytokine production and give rise to typical recall responses suggesting that this cell population is not exclusively naïve in the elderly (Pfister et al. 2006). All these facts imply that the CD8+CD28+ T cell subset is quite distinct between young and elderly persons and the difference in gene expression rather reflects the heterogeneous composition of this T cell population.
Taken together, our results confirm that a significant number of differentially expressed genes underlie the differentiation from naïve to effector and memory T cells. Many genes in each group fit well with known biological differences between these subsets, providing evidence that our data are valid. In conclusion, our study demonstrates a dichotomy of gene expression levels between CD8+CD28+ T cells in young and elderly persons and similarity between CD8+CD28− T cells from young and elderly persons. Together with previous findings from our group these results suggest that the gene expression profile does not depend on chronological age, but rather on the number of differently functioning cell types in a given population.
References
Alexandrakis MG et al (2003) Relationship between circulating serum soluble interleukin-6 receptor and the angiogenic cytokines basic fibroblast growth factor and vascular endothelial growth factor in multiple myeloma. Ann Hematol 82(1):19–23
Alonso MA, Weissman SM (1987) cDNA cloning and sequence of MAL, a hydrophobic protein associated with human T-cell differentiation. Proc Natl Acad Sci USA 84(7):1997–2001. doi:10.1073/pnas.84.7.1997
Day CL et al (2006) PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature 443(7109):350–354. doi:10.1038/nature05115
Eisen MB et al (1998) Cluster analysis and display of genome-wide expression patterns. Proc Natl Acad Sci USA 95(25):14863–14868. doi:10.1073/pnas.95.25.14863
Fagnoni FF et al (1996) Expansion of cytotoxic CD8+ CD28− T cells in healthy ageing people, including centenarians. Immunology 88(4):501–507. doi:10.1046/j.1365-2567.1996.d01-689.x
Fann M et al (2005) Gene expression characteristics of CD28null memory phenotype CD8+ T cells and its implication in T-cell aging. Immunol Rev 205:190–206. doi:10.1111/j.0105-2896.2005.00262.x
Fitzgerald JE et al (1995) Analysis of clonal CD8+ T cell expansions in normal individuals and patients with rheumatoid arthritis. J Immunol 154(7):3538–3547
George AJ, Ritter MA (1996) Thymic involution with ageing: obsolescence or good housekeeping? Immunol Today 17(6):267–272. doi:10.1016/0167-5699(96)80543-3
Hiraide A et al (2005) Fas polymorphisms influence susceptibility to autoimmune hepatitis. Am J Gastroenterol 100(6):1322–1329. doi:10.1111/j.1572-0241.2005.41053.x
Holub MC et al (2002) Soluble interleukin-6 receptor enhanced by oncostatin M induces major changes in gene expression profile of human hepatoma cells. Immunol Lett 82(1–2):79–84. doi:10.1016/S0165-2478(02)00022-6
Hou M et al (2004) Antisense Tiam1 down-regulates the invasiveness of 95D cells in vitro. Acta Biochim Biophys Sin (Shanghai) 36(8):537–540
Kaneko H et al (1996) Preferential elimination of CD28+ T cells in systemic lupus erythematosus (SLE) and the relation with activation-induced apoptosis. Clin Exp Immunol 106(2):218–229. doi:10.1046/j.1365-2249.1996.d01-849.x
Mi H et al (2005) The PANTHER database of protein families, subfamilies, functions and pathways. Nucleic Acids Res 33(Database issue):D284–D288. doi:10.1093/nar/gki078
Nociari MM, Telford W, Russo C (1999) Postthymic development of CD28−CD8+ T cell subset: age-associated expansion and shift from memory to naive phenotype. J Immunol 162(6):3327–3335
Okkenhaug K, Vanhaesebroeck B (2003) PI3K in lymphocyte development, differentiation and activation. Nat Rev Immunol 3(4):317–330. doi:10.1038/nri1056
Pavlidis P, Noble WS (2001) Analysis of strain and regional variation in gene expression in mouse brain. Genome Biol 2(10):RESEARCH0042
Pfister G et al (2006) Naive T cells in the elderly: are they still there? Ann N Y Acad Sci 1067:152–157. doi:10.1196/annals.1354.018
Posnett DN et al (1994) Clonal populations of T cells in normal elderly humans: the T cell equivalent to “benign monoclonal gammapathy”. J Exp Med 179(2):609–618. doi:10.1084/jem.179.2.609
Radfar S, Davrinche C, Hollande E (2005) Serial in vivo loss and in vitro gain of Fas expression and function in human cancerous pancreatic duct cells. Int J Cancer 115(2):214–223. doi:10.1002/ijc.20802
Rainer J et al (2006) CARMAweb: comprehensive R- and bioconductor-based web service for microarray data analysis. Nucleic Acids Res 34(Web Server issue):W498–W503
Rozen S, Skaletsky H (2000) Primer3 on the WWW for general users and for biologist programmers. Methods Mol Biol 132:365–386
Saeed AI et al (2003) TM4: a free, open-source system for microarray data management and analysis. Biotechniques 34(2):374–378
Sallusto F et al (1999) Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature 401(6754):708–712. doi:10.1038/44385
Sansoni P et al (1997) T lymphocyte proliferative capability to defined stimuli and costimulatory CD28 pathway is not impaired in healthy centenarians. Mech Ageing Dev 96(1–3):127–136. doi:10.1016/S0047-6374(97)01887-3
Schwab R et al (1997) Expanded CD4+ and CD8+ T cell clones in elderly humans. J Immunol 158(9):4493–4499
Sperling AI, Bluestone JA (1996) The complexities of T-cell co-stimulation: CD28 and beyond. Immunol Rev 153:155–182. doi:10.1111/j.1600-065X.1996.tb00924.x
Stulnig T et al (1995) Reference intervals for human peripheral blood lymphocyte subpopulations from ‘healthy’ young and aged subjects. Int Arch Allergy Immunol 108(3):205–210
Tarazona R et al (2000) Increased expression of NK cell markers on T lymphocytes in aging and chronic activation of the immune system reflects the accumulation of effector/senescent T cells. Mech Ageing Dev 121(1–3):77–88. doi:10.1016/S0047-6374(00)00199-8
van Doorn R et al (2004) Aberrant expression of the tyrosine kinase receptor EphA4 and the transcription factor twist in Sezary syndrome identified by gene expression analysis. Cancer Res 64(16):5578–5586. doi:10.1158/0008-5472.CAN-04-1253
Wills MR et al (2002) Identification of naive or antigen-experienced human CD8(+) T cells by expression of costimulation and chemokine receptors: analysis of the human cytomegalovirus-specific CD8(+) T cell response. J Immunol 168(11):5455–5464
Acknowledgements
The authors thank Michael Keller and Brigitte Jenewein for collecting blood samples. This work was supported by the Austrian Science Fund (Projects S9308-B05 and S9309-B05
).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lazuardi, L., Herndler-Brandstetter, D., Brunner, S. et al. Microarray analysis reveals similarity between CD8+CD28− T cells from young and elderly persons, but not of CD8+CD28+ T cells. Biogerontology 10, 191–202 (2009). https://doi.org/10.1007/s10522-008-9167-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10522-008-9167-1