
Abstract
The diurnal questing behavior of Amblyomma mixtum Koch was monitored in the laboratory while held on a 12:12 diurnal cycle, and outdoors, using actographic chambers to determine the daily incidences of questing for a host. Nymphs and adults increased their questing activity during the morning. Adult activity peaked in the morning and continued until afternoon, whereas nymphal activity peaked in the afternoon. Nymphs and adults returned to the bottom of the chambers at night. The questing pattern for nymphs held outdoors was similar to that observed indoors. Both nymphs and adults had a higher average questing height when held at 95 % RH than at 56 % RH. The increase in average questing height was found to be due to fewer ticks questing–and for adults, a decrease in the mean questing height–at lower RH.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
“Amblyomma cajennense Fabricius sensu lato” was reclassified into six distinct species (Nava et al. 2014) based on developmental features (Guglielmone et al. 1992), cross breeding experiments (Labruna et al. 2011; Mastropaolo et al. 2011), and genetic analysis (Beati et al. 2013). The name A. mixtum was reinstated. Amblyomma mixtum is found from south Texas to Panama, in some Caribbean Islands and Ecuador (Beati et al. 2013; Nava et al. 2014). In the drier climate of South Texas A. mixtum represents 94 % of ticks on dry ice traps (Beck et al. 2011). This tick aggressively feeds on humans; representing 20 % of ticks submitted to the Texas Department of Health Services while only being found in southern Texas (Williamson et al. 2010). Amblyomma mixtum feeds on some birds but especially on mammals (Guzmán-Cornejo et al. 2011; Nava et al. 2014). Amblyomma mixtum is a vector of Rickettsia spp, Ehrlichia chaffeensis, Borrelia spp, Coxiella burnetti, and Venezuelan equine encephalomyelitis virus (Bermúdez et al. 2009; Billings et al. 1998; de Rodaniche 1949; Gordilla-Perez et al. 2009; Linthicum et al. 1991; Sanders et al. 2008; Williamson et al. 2010).
Questing (seeking a host) is the vertical movement of a tick on vegetation, followed by waiting for a host (Parola and Raoult 2001). Questing ticks are either actively questing (with front legs stretched out waiting to latch on to the passing host) or passively questing (with front legs folded waiting for stimuli to become active). Hosts provide these stimuli via movement (vibrations and/or shadows blocking the light), CO2 or the pheromones they release. Therefore the presence of an observer (a potential host) influences tick behavior (McMahon and Guerin 2002). Thus when observing tick questing it is critical to avoid disturbing them. Tick questing height and time of questing (whether diurnal or nocturnal) effects what hosts a tick may attach to. In addition the relative humidity (RH) can effect tick questing behavior (Madden and Madden 2005). Actographic chambers have been used for observing tick questing under controlled conditions (Madden and Madden 2005). Here we report on the questing behavior of A. mixtum adults and nymphs under 95 and 56 % RH.
Materials and methods
Tick collection
The study site was on the grounds of Texas A&M International University in Laredo, TX (27°57′N 99°44′W) with an elevation of 155 m. Approximately 0.5 kg of dry ice was set on a trap (a 1 × 0.8 m white flannel cloth) and left for 2.5 h. Traps were placed from 0900 to 1000 hours and collected from 1200 to 1300 hours. Traps were placed in sealed bags and taken to the laboratory for tick removal and identification (Cooley and Kohls 1944; Jones et al. 1972; Keirans and Durden 1998; Kohls 1958). Ticks were placed in the actographic chamber and allowed to acclimate for 24–120 h prior to data collection.
Actographic chamber
An actographic chamber was constructed of two 0.225 cm thick acrylic (polymethyl methacrylate) panels (Fig. 1). The sides of the adult chamber were formed by a 0.4 cm thick acrylic strip glued with plastic epoxy. The front panel was secured using 6 mm machine screws spaced every 5 cm. The chambers for adults were 115 × 5.8 × 0.4 cm, and for nymphs were 85 × 13 × 4 cm (with wooden side panels). Mosquito netting was glued over a 3 cm deep water well which allowed for 100 % RH at the bottom of the chamber. A plastic tube at the top of the well allowed the addition of water as needed. Air entered the chamber through a plastic tube at 7 cm height, which left a 7 cm zone at the bottom of the chamber where the ticks could recover from the drier conditions above. Two small air outlets drilled into the top of the chamber allowed air out of the chamber. The chamber was mounted on white contact paper covered plywood (130 × 30 cm) and marked every 10 cm to monitor questing height. The first 10 cm zone was broken into two 5 cm zones (not shown). For outdoor use the nymph chamber was modified with a 3 cm diameter opening at a height of 50 cm in the back of the chamber covered with mosquito netting. A large well of water and a 9 V fan were used to blow humidified air into the chamber (resulting in a RH of 75 % at 20 °C) to prevent the chamber from drying out. The temperature of the outdoor chamber varied from 21.6 to 41 °C. Either 200 nymphs or 20–46 adults were placed in each chamber.

The acrylic actographic chambers used for adults were attached to a board for support and with markings every 10 cm. Humidified air entered the chamber at 7 cm of height. Below this point humidity would be higher than the rest of the chamber as a water well covered with nylon mesh was provided at the bottom of the chamber. Humidified air exited through the top of the chamber through small air vents
Environmental conditions of the indoor actographic chambers
Continuous air flow was maintained at 1.3 L/min. The 95 % RH was generated by passing air through 500 mL of water in an Erlenmeyer flask to achieve 100 % RH, then through 500 mL of a saturated solution of potassium nitrate (KNO3) in a second flask to reduce the RH to 95 % (Madden and Madden 2005). A cotton plug was placed into the hose that led into the chamber to prevent any aerosolized potassium nitrate from entering the chamber and killing the ticks. The 56 % RH was generated in a similar fashion except both flasks had 500 mL of a saturated solution of calcium nitrate (CaNO3). The resulting RH was tested using a humidity meter.
The light chamber was a cart covered with heavy cloth to prevent outside light from entering the chamber. Two sets of lights were placed inside of the cart with two full spectrum bulbs of 20 watts each. Light was cycled on a 12 h photoperiod with low light conditions (1 set on) from 0700 to 0915 hours and from 1735 to 1850 hours, and high light from 0915 to 1735 hours with both sets on. Ticks were counted hourly. During the dark hours a red light was used to prevent disturbing them. Ixodid ticks generally do not respond to low irradiance red light (Carroll and Pickens 1987; Leuterer and Gothe 1991). Student’s t tests were performed by using Microsoft Excel 2010.
Results
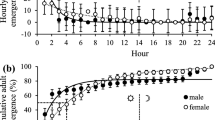
Increased questing activity of A. mixtum nymphs was observed in the laboratory during the light cycle. As shown in Fig. 2, tick questing increased from the morning, 0700 hours, until 1700 hours. At 1735 hours the light intensity was decreased by half, and nymph questing steadily declined until dark. Few ticks were observed questing during the night (data not shown). Ticks maintained under 95 % RH were observed to start questing earlier than ticks maintained under 56 % RH (p < 0.001). This resulted in a decreased average questing height in the morning hours, but by 1700 hours the average questing height was similar under both conditions (Fig. 2a). The difference in mean questing height was explained by a decreased percentage of nymphs questing at 56 % RH (Fig. 2b). Amblyomma mixtum nymphs maintained at 56 % RH for prolonged periods of time became quiescent before the ticks maintained at 95 % RH (data not shown). A similar unimodal pattern with a peak at 1600 hours was observed under natural light (under a shade tree) conditions (Fig. 3). Nymphs quested at similar heights in indoor and outdoor experiments except more nymphs were at the lowest height outdoors (Table 1). Over 30 % of all questing nymphs under all conditions tested quested above 30 cm, and over 65 % above 10 cm. Unless they were disturbed, 36 % (SEM = 3.3) of nymphs were actively questing. If they were disturbed by gently shaking or blowing CO2 into the chamber many ticks became active for 20–30 min before returning to normal questing.

a Mean tick height in cm of Amblyomma mixtum nymphs maintained in an environment with 95 % RH (black line) or 56 % RH (dashed line). b Percentage of A. mixtum nymphs found at a height of 10 cm or more in the actographic chamber. (Error bars show the standard error of the mean)

Mean (± SEM) percentage of Amblyomma mixtum nymphs found at a height of 10 cm or more in the actographic chamber maintained in an outdoor environment under a mesquite tree to provide shade from direct sunlight
Amblyomma mixtum adults quested primarily during the light cycle of the day (Fig. 4). At 95 % RH adult questing activity peaked early (by 0800 hours) before the second set of lights came on. They remained questing until beginning a gradual decline of mean questing height after 1500 hours (2 h before the second set of lights were turned off). At 56 % RH fewer adults quested (p < 0.001). A cross over experiment was done in which the adults that had been held at 56 % RH were now provided 95 % RH. After 10 days at 95 % RH the mean percentage of questing adults from 1000 to 1500 hours was 63 % (SEM = 3.5). This demonstrates that ticks at the lower RH were still viable but quiescent under unfavorable conditions. For adults where the RH was decreased from 95 to 56 % the mean percentage of questing adults dropped rapidly from 70 % (SEM = 1.8) at 5 days, to 49 % (SEM = 1.1) at 10 days and finally to 17 % (SEM = 1.1) at 15 days. The pattern of peak questing from 0800 to 1500 hours remained until 15 days when too few ticks were questing to create a discernable pattern. Adults quested primarily at heights below 60 cm (Table 1). Although adults were occasionally found questing at the top of the chamber (115 cm) only 3.5 % (SEM = 0.6) or 1.0 % (SEM = 0.5), at the 95 and 56 % RH levels respectively, quested above 60 cm. A higher percentage of adults quested at higher heights at 95 % RH than at 56 % RH (p < 0.001).

a Mean tick height in cm of Amblyomma mixtum adults maintained in an environment with 95 % RH (black line) or 56 % RH (dashed line). b Percentage of A. mixtum adults found at a height of 10 cm or more in the actographic chamber. (Error bars show the standard error of the mean)
Discussion
Both nymph and adult A. mixtum quested primarily during the day and generally returned to the bottom of the chambers during the night, during which very low activity was seen (2400–0400 hours data not shown). The advantage of diurnal questing is not readily apparent, because nocturnal rodents would still be active at ground level after dark. Questing height usually correlates with host preference. When rodents are the favored host for I. ricinus nymphs (drier months) they quested at lower heights. The opposite was the case (moister months) when primary hosts were larger animals (Randolph 2004; Randolph and Storey 1999). The unimodal questing pattern for A. mixtum is similar to the one observed for A. americanum. Amblyomma americanum nymphs and adults increased from low numbers per drag at 0600 hours to higher numbers per drag at 1500–1600 hours (Schulze et al. 2001; Schulze and Jordan 2003). In contrast I. scapularis adults had a bimodal questing pattern peaking in the morning and evening in the fall, with a unimodal evening peak in the spring. I. scapularis nymphs had a unimodal dusk peak (Schulze and Jordan 2003), while I. ricinus nymphs peaked after sunset (Perret et al. 2003). These different questing patterns are in part due to host preferences of these ticks and different life stages of ticks. In our study the majority of both adult and nymphal A. mixtum quested above 10 cm during the day. This is consistent with the preference of A. mixtum for medium to large sized diurnal mammals as hosts (Guzmán-Cornejo et al. 2011; Nava et al. 2014). Further research is needed into the host preferences of A. mixtum.
The visible spectrum of A. mixtum is unknown. Amblyomma variegatum Fabricius has a single peak at 480 nm (Kaltenrieder et al. 1989). In a number of different species of ixodid ticks there typically are one or two peaks in electroretinogram responses to light, a lower one of 350–380 nm is sometimes found and a conserved higher peak of 470–510 nm (Carrol and Pickens 1987; Kaltenrieder et al. 1989; Leuterer and Gothe 1991). Electroretinogram responses in ixodid ticks typically are very low at 650 nm in the red range of the visible spectrum. Rhipicephalus (Boophilus) microplus larvae (Waladde and Rice 1982) and Dermacentor variabilis did not respond to red light but 48 % of Hyalomma dromedarii did (Carrol and Pickens 1987). At lower irradiance H. truncatum primarily responds to blue light (Leuterer and Gothe 1991). Neither A. mixtum adults nor nymphs responded to brief exposure to red light, but if the red light was shone very close to the chamber some would gradually become active. Similar to observations in other tick species (Waladde and Rice 1982) if the regular light was disrupted by creating a shadow in the chamber the ticks became active for a period of 20–30 min. Thus A. mixtum probably responds more to green and blue light than red light as in other ixodid tick species.
Both temperature and saturation deficit affect tick questing behavior (Randolph 2004). Commonly few ticks quest during the middle of the day due to increased saturation deficits at lower RH (Randolph 2004). In contrast A. mixtum primarily quested during the day. At low RH nymphs quested primarily in the afternoon. Adults at low or high RH quested starting in the morning and remained questing until late in the day. Amblyomma mixtum is most dehydration tolerant of the studied ixodid ticks. The 50 % mortality rate of A. mixtum adults is 507 days under ideal conditions (85 % RH and 23 °C) and 37 days under dehydrating conditions (35 % RH and 23 °C) (Strey et al. 1996). However, high saturation deficit is associated with quiescence prior to mortality (Randolph 2004, Randolph and Storey 1999). More than 50 % of A. mixtum ticks subjected to more than 5 exposures of 24 h to 0 % RH were able to recover ambulatory status (Strey et al. 1996). Similarly we observed that adult A. mixtum would often become quiescent at low RH. However when conditions were changed to 95 % RH many recovered and began questing again.
Questing ticks search for favorable habitats and humidity to prevent physiological stress (Randolph 2004). Ixodes ricinus nymphs were more mobile during desiccating conditions, suggesting they may be looking for better conditions for off-host survival (Perret et al. 2003). The effect of humidity on the horizontal movement of A. mixtum was not tested in this study. Another limitation of container experiments is ticks may have a lower rate of survival in the container than under natural conditions. For instance R. appendiculatus larvae positioned themselves near stomata on plant grass stems (Branagan 1973). Thus in the natural environment ticks may quest at higher percentages or at different heights than predicted by laboratory experiments. In the field A. mixtum prefers areas with more dense or mixed vegetation than open areas with short grasses (Beck et al. 2011; Smith 1975). These areas are associated with a higher RH at ground level due to the increased brush density and decreased sunlight at ground level. Further studies are needed to determine A. mixtum questing activity under these conditions.
Understanding the parameters that control the life cycle of A. mixtum is essential to generate predictive models for risk of disease transmission. Efficient management of diseases transmitted by this tick may be greatly enhanced by the development of these predictive models. The knowledge of the questing behavior of A. mixtum will help humans avoid contact with these ticks and their various pathogens.
References
Beati L, Nava S, Burkman EJ, Baros-Battesti DM, Labruna MB, Guglielmone AA, Cáceres AG, Guzmán-Cornejo CM, León R, Durden LA, Faccini LH (2013) Amblyomma cajennense (Fabricius 1787) (Acari:Ixodidae), the Cayenne tick: phylogeography and evidence for allopatric speciation. BMC Evol Biol 13:267
Beck DL, Zavala J, Montalvo EO, Quintana FG (2011) Meteorological indicators for Amblyomma cajennense population dynamics in the Tamaulipan Biotic Province in Texas. J Vector Ecol 36:135–146
Bermúdez SE, Eremeeva ME, Karpathy SE, Samudio F, Zambrano ML, Zal-divar Y, Motta JA, Dasch GA (2009) Detection and identification of rickettsial agents in ticks from domestic mammals in eastern Panama. J Med Entomol 46:856–861
Billings AN, Yu XJ, Teel PD, Walker DH (1998) Detection of a spotted fever group rickettsia in Amblyomma cajennense (Acari: Ixodidae) in South Texas. J Med Entomol 35:474–478
Branagan D (1973) Observations on the development and survival of the Ixodid tick Rhipicephalus appendiculatus Neumann, 1901 under quasi-natural conditions in Kenya. Trop Anim Health Prod 5:153–165
Carroll JF, Pickens LG (1987) Spectral sensitivity to light of two species of ticks (Acarina: Ixodidae). Ann Entomol Soc Am 80:256–262
Cooley RA, Kohls GM (1944) The genus Amblyomma (Ixodidae) in the United States. J Parasitol 30:77–111
de Rodaniche EC (1949) Experimental transmission of Q Fever by Amblyomma cajennense. Am J Trop Med Hyg S1–29:711–714
Gordillo-Perez G, Vargas M, Solorzano-Santos F, Rivera A, Polaco OJ, Alvarado L, Munoz O, Torres J (2009) Demonstration of Borrelia burgdorferi sensu stricto infection in ticks from the northeast of Mexico. Clin Microbiol Infect 15:196–498
Guglielmone AA, Mangold AJ, Oyola BC (1992) Ciclo de vida de Amblyomma cajennense (Fabricius 1787) (Acari: Ixodidae) en condiciones de laboratorio. Rev Med Vet (B Aires) 73:184–187
Guzmán-Cornejo C, Robbins RG, Guglielmone AA, Montiel-Parra G, Pérez TM (2011) The Amblyomma (Acari: Ixodida: Ixodidae) of Mexico: Identification keys, distribution and hosts. Zootaxa 2998:16–38
Jones EK, Clifford CM, Keirans JE, Kohls GM (1972) Ticks of Venezuela (Acarina: Ixodoidea) with a key to the species of Amblyomma in the Western Hemisphere. Brigh Young Univ Sci Bull 17:1–40
Kaltenrieder M, Labhart T, Hess E (1989) Spectral sensitivity, absolute threshold, and visual field of two species, Hyalomma dromedarii and Amblyomma variegatum. J Comp Physiol A 165:155–164
Keirans JE, Durden LA (1998) Illustrated key to nymphs of the tick genus Amblyomma (Acari:Ixodidae) found in the United States. J Med Entomol 35:489–495
Kohls GM (1958) Amblyomma imitator, a new species of tick from Texas and Mexico, and remarks on the synonymy of A. cajennense (Fabricius) (Acarina: Ixodidae). J Parasitol 44:430–433
Labruna MB, Soares JF, Martins TF, Soares HS, Cabrera RR (2011) Cross-mating experiments with geographically different populations of Amblyomma cajennense (Acari: Ixodidae). Exp Appl Acarol 54:41–49
Leuterer G, Gothe R (1991) On the reaction of adult Rhipicephalus evertsi mimeticus and Hyalomma truncatum to horizontally incidenting optical radiation of various wavelengths ranges and different irradiances and to optical radiation of a sun-simulating wavelength spectrum. Parasitol Res 77:353–358
Linthicum KJ, Logan TM, Bailey CL, Gordon SW, Peters CJ, Monath TP, Osorio J, Francy DB, McClean RG, Leduc JW, Graham RR, Jahrling PB, Dohm DJ (1991) Venezuelan equine encephalomyelitis virus infection in and transmission by the tick Amblyomma cajennense (Arachnida: Ixodidae). J Med Entomol 28:405–409
Madden SC, Madden RC (2005) Seasonality in diurnal locomotory patterns of adult blacklegged ticks (Acari: Ixodidae). J Med Entomol 42:582–588
Mastropaolo M, Nava S, Guglielmone AA, Mangold AJ (2011) Biological differences between two allopatric populations of Amblyomma cajennense (Acari: Ixodidae) in Argentina. Exp Appl Acarol 53:371–375
McMahon C, Guerin PM (2002) Attraction of the tropical bont tick, Amblyomma variegatum, to human breath and to the breath components acetone, NO and CO2. Naturwissenschaften 89:311–315
Nava S, Beati L, Labruna MB, Cáceres AG, Mangold AJ, Guglielmone AA (2014) Reassesssment of the taxonomic status of Amblyomma cajennense (Fabricius 1787) with the description of three new species, Amblyomma tonelliae n. sp., Amblyomma interandinum n. sp. and Amblyomma patinoi n. sp., and reinstatement of Amblyomma mixtum Koch 1844, and Amblyomma sculptum Berlese 1888 (Ixodida: Ixodidae). Ticks Tick borne Dis 5:252–276
Parola P, Raoult D (2001) Ticks and tickborne bacterial diseases in humans: an emerging infectious threat. Clin Infect Dis 32:897–928
Perret JL, Guerin PM, Diehl PA, Vlimant M, Gern L (2003) Darkness induces mobility, and saturation deficit limits questing duration, in the tick Ixodes ricinus. J Exp Biol 206:1809–1815
Randolph SE (2004) Tick ecology: processes and patterns behind the epidemiological risk posed by ixodid ticks as vectors. Parasitology 129:S37–S65
Randolph SE, Storey K (1999) Impact of microclimate on immature tick-rodent host interactions (Acari: Ixodidae): implications for parasite transmission. J Med Entomol 36:741–748
Sanders DM, Parker JE, Walker WW, Bucholz MW, Blount K, Kiel JL (2008) Field collection and genetic classification of tick-borne Rickettsiae and Rickettsiae-like pathogens from South Texas: Coxiella burnetii isolated from field-collected Amblyomma cajennense. Ann N Y Acad Sci 1149:208–211
Schulze TL, Jordan RA (2003) Meteorologically mediated diurnal questing of Ixodes scapularis and Amblyomma americanum nymphs. J Med Entomol 40:395–402
Schulze TL, Jordan RA, Hung RW (2001) Effects of selected meteorological factors on the diurnal questing of Ixodes scapularis and Amblyomma americanum Acari: Ixodidae). J Med Entomol 38:318–324
Smith MW (1975) Some aspects of the ecology and life cycle of Amblyomma cajennense (Fabricius 1787) in Trinidad and their influence on tick control measures. Ann Trop Med Parasitol 69:121–129
Strey OF, Teel PD, Longnecker MT, Needham GR (1996) Survival and water-balance characteristics of unfed adult Amblyomma cajennense (Acari: Ixodidae). J Med Entomol 33:63–73
Waladde SM, Rice MJ (1982) The sensory basis of tick feeding behavior. In: Obenchain FD, Galun R (eds) Physiology of ticks. Pergamon, Oxford, pp 71–118
Williamson PC, Billingsley PM, Teltow GH, Seals JP, Turnbough MA, Atkinson SF (2010) Borrelia, Ehrlichia, and Rickettsia spp. in ticks removed from person, Texas, USA. Emerg Infect Dis 16:441–446
Acknowledgments
We thank Joseph Brazil and Kimberly Hart for review of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Beck, D.L., Orozco, J.P. Diurnal questing behavior of Amblyomma mixtum (Acari: Ixodidae). Exp Appl Acarol 66, 613–621 (2015). https://doi.org/10.1007/s10493-015-9928-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10493-015-9928-x