
Abstract
Tetranychus urticae (Acari: Tetranychidae) is a phytophagous mite that forms huge colonies. All active members of a colony (immatures and matures, females and males) spin silken threads. These mites construct a common web that protects the colony from external aggression. The silk coverage is well-known to provide advantages to the colony but very little is known about the characteristics of the threads themselves. Here is the first quantification of the diameter of silken threads spun by two different stages (adult females and larvae) and its relationship with body size of the spinning individuals. Moreover, we observed how silk was deposited on the substrate through their two pedipalps. Threads were observed by means of transmission electron and fluorescence microscopy. Silken threads spun by larvae (0.055 ± 0.018 μm) were significantly thinner than threads spun by adult females (0.111 ± 0.038 μm). In the first step of the silk depositing behaviour, the mite attached the thread to the substrate by putting its pedipalps in contact with the surface (adhesion, double silken threads). When walking, silken threads became detached from the substrate and spitted up (silken threads were free). Finally, silken threads adhered to the surface. The presence of single and double threads makes thread diameter highly variable.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Silk produced by arthropods is often described as fibrous biopolymer filament or thread, which displays a wide diversity of physical properties and chemical constituents (Foelix 1996; Vollrath 1999). Silk is produced by a variety of insects; it is especially common for protection during pupal metamorphosis (silken cocoons) in the Lepidoptera and Hymenoptera (e.g. honey bees and sphecid wasps), and is sometimes used in nest construction (e.g. weaver ants). Also non-insect arthropods produce silk, most notably arachnids such as spiders (Craig 1997).
The function of silk is manifold: protective shelter (e.g. cocoon produced by Lepidoptera), structural support (e.g. silk egg stalk produced by Neuroptera), reproduction (e.g. restriction of female movement during mating by Thysanoptera), foraging (e.g. underwater silk prey capture nets by Trichoptera), or dispersal (e.g. Lepidoptera or spiderlings) (Craig 1997). Moreover, some insects (e.g. Lepidoptera) and spiders use their silk as lifeline or as a substrate for pheromonal compounds among communicating individuals (e.g. caterpillars, spiders) (Anderson and Morse 2001; Bernard and Krafft 2002; Clark et al. 1999; Fitzgerald 1993; Saffre et al. 1999).
The best-known example of weaving arthropods is the mulberry silkworm, Bombyx mori. Its silk can be woven into textiles and is obtained from cocoons of silkworms reared in captivity (sericulture). Silkworm threads are naturally extruded from two silkworm glands and are 5–10 μm wide (Menachem 2006). Another well-known example is spiders that weave among the strongest and toughest threads in the living world. Various specialized silks have evolved with properties suitable for different uses. The diameter for their silk ranges from a few 100 nm–50 μm (Foelix 1996; Vollrath 1999).
In our study, we are interested in the silk spun by the two-spotted spider mite Tetranychus urticae, a widespread phytophagous mite that is known as a major pest in many cultures. It can be found on plants of numerous species (Bolland et al. 1998). One of the characteristics of T. urticae is its abundant silk production: while walking silk is deposited continuously (Saito 1977). Spider mite silk is produced in the whole podocephalic complex (Mills 1973). It is secreted by tubular and reniform propodosomal glands and excreted through orifices in the pedipalps (eupathids) (Blauvelt 1945; Grandjean 1948). Various types of secretion (such as serous, mucous, ion-rich) are found both in females and males (Mothes and Seitz 1981).
Silk of T. urticae has been reported to play four main roles: (1) protection against biotic agents like mites predators (Helle and Sabelis 1985; Sabelis and Bakker 1992), (2) protection against abiotic agents, such as rain and acaricides (Gerson 1985), (3) sex pheromone substrate (Cone et al. 1971; Penman and Cone 1974) and (4) support for locomotion and dispersal (Saito 1977, 1979; Yano 2008; Clotuche et al. 2011). Although substantial work has been done on the role of silk in the biology of T. urticae (Cone et al. 1971; Davis 1952; Hazan et al. 1975; Helle and Sabelis 1985; Hoy and Smilanick 1981; Linke 1953; Penman and Cone 1972, 1974; Sabelis and Bakker 1992; Saito 1977, 1979; Yano 2008), the silk itself remains poorly studied.
Here, we present a first study on the threads produced by the two-spotted spider mite. Our aim was to measure the diameter of silken threads and to search for correlations with the developmental stage and body size of mites. Measurements were done on larvae and adult females. Emerging larvae begin to feed and soon afterwards spin some loose silken threads, while newly emerged adult females produce webbing even before they feed (Hazan et al. 1974). We also made behavioural observations on how silken threads are spun and deposited by the two pedipalps.
Materials and methods
Rearing of mites
Mites (strain LS-VL) used in this study were collected in October 2000 from roses (Ghent, Belgium) (Van Leeuwen et al. 2008). In the laboratory, the two-spotted mites were reared on bean (Phaseolus vulgaris). Leaves were placed on damp cotton wool in Petri dishes (85 mm in diameter, 13 mm deep). Stocks were maintained in climate room at 26°C, 50–60% relative humidity and L16:D8 photoperiod.
Silk collection and measurement
The first type of observations aimed at counting the silken threads deposited per individual, and measuring the threads’ cross-sectional area and the body size of the laying mites. For each experiment, one mite was deposited on a grid for a period of 20 h and allowed to deposit the silken threads. The grid was a Formvar/Carbon grid for transmission electron microscopy (TEM) (200 mesh Cu, 3.05 mm diameter; Ted Pella, Redding, CA, USA), consisting of a thin cast film of Formvar strengthened with a thin evaporated layer of carbon. The grids with the spider mites were individually deposited on thin plastic discs (5.5 mm diameter) (Fig. 1a). Plastic discs were placed on damp cotton wool in 85-mm Petri dishes to prevent mites from escaping.

Experimental design for collecting silken threads on the Formvar/Carbon grid. a Individuals were placed in the set-up (grid + plastic disc) for 20 h. Afterwards, grids were removed and observed under a microscope. b Pictures of larvae on grids were taken to measure their body sizes. The size of the grid holes (200 mesh) served as size reference
In the set-up, since mites could not feed or defecate (or very little), the observed silken threads were clean. Within 20 h, the spider mites had spun enough silken threads onto the grids to allow their observation. Mites were then removed from the grids with great care by using a brush with one hair. Grids were kept safely in a grid storage box and then were immediately observed (less than 1 h after removing the mites) under a microscope in order to prevent degradation of the threads.
One 1-day-old adult female was tested per grid (N = 10 grids). Since larvae were less mobile (and therefore spun less), we put three freshly hatched larvae (less than 1-day old) on a grid (N = 10 grids). Body length of the tested adults were measured after being frozen (24 h at −20°C) using a micrometer (Graticules, Pyser-SGI). Because larvae were small and fragile, we did not use the micrometer to measure their body length. Instead, photos (Panasonic WV-CP450/G numeric camera with a 1:1.2/12.5–75 macroscopic lens) of grids were taken before removal of the larvae. The size of the grid holes (200 mesh) served as reference to determine the size of larvae (Fig. 1b). Photos were analysed with ImageJ® software.
Silk images were captured using a JEM-100SX tungsten-filament TEM (JEOL, 10,000× magnification). Output was 6.3 × 8.8 cm black-and-white film negative, which was digitally scanned. Twelve photos were taken per grid. To obtain a large number of threads by photo and therefore by individual (grid), only the intersections of two or more threads were photographed. A total of 10 grids (threads spun by either one adult female or by three larvae) per stage were observed. Once the photos were digitized, silken threads were analyzed with ImageJ®. We counted the threads deposited per individual and the cross-sectional area of silk (diameters) was measured 6× over the length of each silken thread.
Silk deposit observation
The second type of observations aimed at describing silk deposit (link between walking and silk secretion through the pedipalps). The coloration technique described in Clotuche et al. (2009) was used to visualize the silk. A standardized adult female (2 days old) was introduced on one lens for a period of 48 h. Once the mite was removed, cover glasses were colored with a Calcofluor white solution (DMSO, 0.5%) (Clotuche et al. 2009). Images of colored cover glasses were captured using a Leica DMR epifluorescence microscope. To observe and photograph threads, we used a Leica DAPI filter and the image capture program Orca AG Hamamatsu digital charge-coupled device (CCD) camera driven by Hokawo 2.1 software. Silk of spider mites was observed at 10×, 20× and 40× magnifications.
Adult females were also observed under the microscope to examine their weaving behaviour (on cover glasses and bean leaf discs).
Data analysis
As data obtained for the silken thread diameters were not normally distributed, we only used non-parametric tests. Kruskal–Wallis test was used to determine whether the mean silk diameter observed per grid differed among the 10 grids analyzed for each developmental stage (larvae and adult females). Mann–Whitney U test was used to assess differences in thread diameter and body size between the two stages studied (adult female vs. larva). Linear regression was used to determine the relationship between mean thread diameter and individual body size. Kolmogorov–Smirnov test was used to determine whether the two datasets were different; it was used to assess whether the thread diameter distributions for larvae and females corresponded to a bimodal distribution (the addition of two normal curves, one for the single threads and the other for the double threads). Tests were performed using GraphPad Prism version 5.01 for Windows (GraphPad Software, San Diego, CA, USA; http://www.graphpad.com).
Results
Silken threads of larvae
The diameter of threads secreted by larvae ranged from 0.021 to 0.113 μm (mean ± SD = 0.055 ± 0.018, N = 207; Fig. 2a). Regarding the mean silk diameter observed per grid among the 10 grids analyzed (grids comparison), the mean silk diameter per grid was statistically different (Kruskal–Wallis test: KW = 60.01, p < 0.0001, N = 10). The 10,000× magnification of the TEM did not allow an accurate assessment of the total length of threads. Instead we estimated the number of threads spun per larva based on the total number of threads seen in the 12 photos. We counted 307 silken threads spun by 30 larvae (3 larvae per grid); during 20 h each larva spun between 8 and 13 threads (mean ± SD = 10.3 ± 1.9 threads per larva).

Frequency distribution of silken thread diameters for a larvae (class width: 0.0025 μm) and b females (class width: 0.005 μm). Diameters of silken threads for both stages did not show a normal distribution (Kolmogorov–Smirnov test)
Silken threads of adult females
The diameter of threads secreted by adult mites ranged from 0.042 to 0.207 μm (mean ± SD = 0.111 ± 0.038, N = 307; Fig. 2b). Adult females spun thicker threads than larvae (Mann–Whitney test: U = 4,973, p < 0.0001, Fig. 3). Regarding the mean silk diameters observed per grids analyzed (grids comparison), we found significant differences among the mean silk diameter per grid (Kruskal–Wallis test: KW = 28.54, p = 0.0008, N = 10 grids). An adult female spun 16–28 threads in 20 h (mean ± SD = 20.7 ± 3.5 threads per female). Adult females spun more threads than larvae (Mann–Whitney test: U = 0.000, p = 0.0002, N = 10 grids).

The width of silken threads was influenced by the stage of the mites (Mann–Whitney test, U = 5,466, p < 0.0001; N = 307 for larvae, N = 207 for adults)
Relationship between mite size and thread diameter
The diameter of silken threads was not correlated with mite body length of either stage (linear regressions, adult females: y = 0.0001x + 0.0683, spearman r = 0.13, p = 0.73, R² = 0.148, N = 10; larvae: y = 0.4366x − 0.0097, spearman r = 0.28, p = 0.46, R² = 0.255, N = 9). Mean body length of adult females: 0.348 ± 0.047 mm, of larvae: 0.149 ± 0.011 mm.
Spinning scenario
During microscope analysis, we observed that some threads were composed of two single ones (Y shape, Fig. 4a). Diameters of double threads were the sum of single threads (linear regression, y = 1.012x, Spearman r = 1.0, p = 0.0004, R² = 0.99, N = 7).

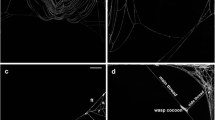
a A double silken thread spun by an adult female, made from two single ones (TEM, ×10,000). b Single silken threads spun by adult females (fluorescence microscope). c The two pedipalps of a female. Silken threads are spun through two eupathids (micrograph of LR Tiedt, NWU, RSA)
The spinning of two categories of silk, single and double threads, can explain the distribution of silken threads observed (Fig. 2). Indeed, the overall distribution of the diameters of silken threads spun by adult females did not fit a Gaussian distribution (Kolmogorov–Smirnov test: KS = 0.084, p = 0.001), but fitted a bimodal distribution with 50% single threads (diameter 0.07 ± 0.019 μm) and 50% double threads (0.14 ± 0.038 μm) (KS = 0.133, p > 0.05, N = 207; Fig. 2b). Similarly, the observed distribution of diameters of threads spun by larvae did not fit a Gaussian distribution (KS = 0.071, p = 0.021), but fitted a bimodal distribution with 20% single threads (0.03 ± 0.008 μm) and 80% double threads (0.06 ± 0.016 μm) (KS = 0.101, p > 0.2, N = 307; Fig. 2a).
The observations of pictures of silken threads with the fluorescent technique (Clotuche et al. 2009) and direct microscope observations of mites during spinning led to the following ‘spinning scenario’ (see also Saito 2009): first, the mite sits still and adheres the silk to the substrate by putting its pedipalps in contact with the surface; the thread is double stranded. Once the mite starts walking, threads begin to split up (two single strands). During spinning behaviour, mites always spun one double or two single silken threads, we never observed mites spinning only one single thread. In some cases, the deposited silk followed the mite displacements because the mite kept its pedipalps onto the substrate surface (Fig. 4b). The distance between the two silken threads in such double tracks (mean ± SD = 20.29 ± 10.65 μm, N = 7, adult females) was precisely the distance between the two pedipalps (around 20 μm for a young adult female, according to LR Tiedt; Fig. 4c).
Discussion
The fibrous biopolymer filaments or threads spun by T. urticae are much thinner (ca. 0.055 μm for larvae, ca. 0.111 μm for adults) than the silken threads from other spinning arthropods—for example, trichopteran threads reach 1–4 μm diameter, and those from lepidopterans 15–65 μm (Engster 1976). In spiders, silk diameter is highly variable within and between species, ranging from a few hundred nanometers to 50 μm (Foelix 1996; Vollrath 1999). The diameter of T. urticae silks appears even thinner than macroscopic threads of collagen (2–20 μm) (Gross and Schmitt 1948) and elastin (4.5–6.0 μm) (Gotte et al. 1972). Compared to other spinning arthropods, the tiny silken threads found in T. urticae may have different physical and mechanical properties (e.g., strength or elasticity).
The high variation in silken thread diameter observed can be explained by the presence of single and double threads. In females, the bimodal distribution of silken thread diameters is obvious. In larvae, we observed more noise in the distribution, probably because the larvae are not all female. The small larvae (ca. 150 μm) can have their pedipalps so close to each other that relatively many threads are double stranded (80%) compared to single strands (20%). It would be interesting to study how and when T. urticae choose to attach two silken threads by an accurate observation of the use of their pedipalps (for immature and mature stages, males and females). They may bind single silken threads in order to deposit thicker threads on the substrate.
Apart from single or double strands, many factors can influence the diameter of silken threads. For example, it can vary with the speed at which threads are drawn (Madsen et al. 1999), with the length of the threads, and with environmental temperature and/or humidity (Denny 1976; Gosline et al. 1986; Madsen et al. 1999). In our experiment, mites did not have access to food. Starvation may possibly influence the size of the thread to avoid energy spill.
We observed that a female mite deposited 2× more silken threads than a larva. By assuming a cylindrical shape, the volume of one thread is 4× larger if spun by an adult than by a larva (thread volume = πr² × length, with radult = 2 × rlarva). Therefore, the total amount of silk deposited is expected to be around 8× higher for an adult female than for a larva. Because females are bigger (almost 3× the larval length) there spinnerets are also larger. Moreover, adult females have more spendable energy and they move more and faster that larvae. It will be interesting to know how the spinnerets of mites influence the shape of the silken threads (as shown for two species of trichopteran larvae, Engster 1976).
It is well-known that spiders are able to manipulate the diameter of silken threads and thereby alter the mechanical properties of the silk (Garrido et al. 2002; Vollrath and Köhler 1996). Furthermore, spiders can increase the diameter of threads in their orb webs in proportion to their body weight, thus increasing the strength of webs to support them (Vollrath and Köhler 1996). As for some spiders, T. urticae use the silk as a lifeline which must support their body weight and this is probably a constraint for thread thickness. By controlling the diameter, spider mites can alter the physical performance of web structures while using the same material to spin different threads. The fact that they spin two single threads or one double thread at a time probably enables them to build various structures (i.e., it increases the degrees of freedom of their silk spinning behaviour). It would be interesting to observe how the mites build their webs over time under different environmental conditions. Some trade-offs could come into play when the mite is spinning silk. For example, spinning two single threads could allow the individual to build a more complex structure (web) but less resilient than if the web has been woven with double silken threads (trade-off between web complexity and resilience).
The ability to disrupt silk production could have an interesting potential in management of T. urticae colonies. Indeed, together with a high fecundity and short life cycle, webbing and silk production should have contributed to the ecological success of T. urticae and its status of serious plant pest. Because the uses of chemicals are increasingly avoided in order to protect environment and human health (Cranham and Helle 1985; Van Leeuwen et al. 2005), an alternative solution could be to focus on silk. Indeed, an improved control of this pest species requires further information on the structure of the silk glands and threads, on the biochemical composition of silk as well as on biosynthesis pathways. Moreover, as the complete genome of T. urticae is known, it would be possible to suppress the gene for silk production and therefore to obtain individuals unable to produce silk. This would lead to increased vulnerability of spider mite populations to (more species of) natural enemies (e.g. phytoseiid species). Normally, silk produced by the two-spotted spider mite prevents many predators from entering (and controlling) the colony—only phytoseiids with the proper dorsal chaetotaxy are able to penetrate the web (Sabelis and Bakker 1992). This would represent a breakthrough in the biological control of this pest.
A lot of questions remain unanswered. Further investigation is necessary to look for the differences in physical–chemical properties between the different types of threads produced by this mite species and their specific biological role. The unravelling of the spider mites’ silk secrets is only in its infancy.
References
Anderson JT, Morse DH (2001) Pick-up lines: cues used by male crab spiders to find reproductive females. Behav Ecol 12:360–366
Bernard A, Krafft B (2002) Silk attraction: base of group cohesion and collective behaviours in social spiders. C R Biol 325:1153–1157
Blauvelt WE (1945) The internal morphology of the common red spider mite (Tetranychus telarius L.). Cornell Univ Agr Expt Sta Mem 270:1–11
Bolland HR, Flechtmann CHW, Gutierrez J (1998) World catalogue of the spider mite family (Acari: Tetranychidae). Brill Academic Publishers, Leiden
Clark RJ, Jackson RR, Waas JR (1999) Draglines and assessment of fighting ability in cannibalistic jumping spiders. J Insect Behav 12:753–766
Clotuche G, Le Goff G, Mailleux AC, Deneubourg JL, Detrain C, Hance T (2009) How to visualize the spider mite silk? Microsc Res Tech 72:659–664
Clotuche G, Mailleux AC, Astudillo Fernandez A, Deneubourg JL, Detrain C, Hance T (2011) The formation of collective silk balls in the spider mite Tetranychus urticae Koch. Plos one 6(4):1–9
Cone WW, Predki S, Klostermeyer EC (1971) Pheromone studies of the two-spotted spider mite. 2. Behavioral response of males to quiescent deutonymphs. J Econ Entomol 64:379–382
Craig CL (1997) Evolution of arthropod silks. Annu Rev Entomol 42:231–267
Cranham JE, Helle W (1985) Pesticide resistance in Tetranychidae. In: Helle W, Sabelis MW (eds) Spider mites, their biology, natural enemies and control, vol 1A. Elsevier, Amsterdam, pp 405–422
Davis DW (1952) Influence of population density on Tetranychus multistis. J Econ Entomol 45:652–654
Denny M (1976) Physical properties of spider’s silk and their role in design of orb-webs. J Exp Biol 65:483–506
Engster MS (1976) Studies on silk secretion in the Trichoptera (F. Limnephilidae). Cell Tiss Res 169:77–92
Fitzgerald TD (1993) Trail and arena marking by caterpillars of Archips Cerasivoranus (Lepidoptera: Tortricidae). J Chem Ecol 19(7):1479–1489
Foelix RF (1996) The biology of spiders. Oxford University Press, New York
Garrido MA, Elices M, Viney C, Pérez-Rigueiro J (2002) Active control of spider silk strength: comparison of drag line spun on vertical and horizontal surfaces. Polymer 43:1537–1540
Gerson U (1985) Webbing. In: Helle W, Sabelis MW (eds) Spider mites, their biology, natural enemies and control, vol 1A. Elsevier, Amsterdam, pp 223–233
Gosline JM, Demont ME, Denny MW (1986) The structure and properties of spider silk. Endeavour 10:37–43
Gotte L, Mammi M, Pezzin G (1972) Scanning electron microscope observations on elastin. Conn Tiss Res 1:61–67
Grandjean F (1948) Quelques caractères des Tétranyques. Bull Mus Hist Nat Paris 20:517–524
Gross J, Schmitt FO (1948) The structure of human skin collagen as studied with the electron microscope. J Exp Med 88:555–569
Hazan A, Gerson U, Tahori AS (1974) Spider mite webbing. I. The production of webbing under various environmental conditions. Acarologia 16:68–84
Hazan A, Gerson U, Tahori AS (1975) Spider mite webbing—II. The effect of webbing removal on egg hatchability. Comp Biochem Physiol 51:457–462
Helle W, Sabelis MW (1985) Spider mites: their biology, natural enemies and control, vol 1A. Elsevier, Amsterdam, p 405
Hoy M, Smilanick JM (1981) Non random prey location by the phytoseiid predador Metaseiulis occidentalis: differential responses to several spider mite species. Entomologica experimentalis et applicata 29:241–253
Linke W (1953) Investigation of the biology and epidemiology of the common spider mite, Tetranychus althaeae v. Hanst., with particular consideration of the hop as the host. Pflanzenschutz Nachrichten Bayer 6:181–232
Madsen B, Shao ZZ, Vollrath F (1999) Variability in the mechanical properties of spider silks on three levels: interspecific, intraspecific and intraindividual. Int J Biol Macromol 24:301–306
Menachem L (2006) Handbook of fiber chemistry, 3rd edn. CRC press, New York
Mills LR (1973) Morphology of glands and ducts in the two-spotted spider mite, Tetranychus urticae Koch, 1836. Acarologia 15(2):218–236
Mothes U, Seitz KA (1981) Functional microscopic anatomy of the digestive system of Tetranychus urticae (Acari, Tetranychidae). Acarologia 22:257–270
Penman DR, Cone WW (1972) Behaviour of male two-spotted mites in response to quiescent female deutonymph and to web. Ann Entomol Soc Am 65:1289–1293
Penman DR, Cone WW (1974) Role of web, tactile stimuli and female sex pheromone in attraction of male two-spotted spider mites to quiescent female deutonymphs. Ann Entomol Soc Am 67:179–182
Sabelis MW, Bakker FM (1992) How predatory mites cope with the web of their tetranychid prey: a functional view on dorsal chaetotaxy in the Phytoseiidea. Exp Appl Acarol 16:203–225
Saffre F, Furey R, Krafft B, Deneubourg JL (1999) Collective decision making in social spiders: dragline-mediated amplification process acts as a recruitment mechanism. J Theor Biol 198:507–517
Saito Y (1977) Study of the spinning behaviour of the spider mite (Acarina: Tetranychidae). I. Method for the quantitative evaluation of the mite webbing, and the relationship between webbing and walking. Jpn J Appl Entomol Zool 21:27–34
Saito Y (1979) Study of the spinning behaviour of spider mites. III. Responses of mites to webbing residues and their preferences for particular physical conditions of leaf surfaces (Acarina: Tetranychidae). Jpn J Appl Entomol Zool 23:82–91
Saito Y (2009) Plant mites and sociality. Diversity and evolution. Springer, Tokyo, p 187
Van Leeuwen T, Van Pottelberge S, Tirry L (2005) Comparative acaricide susceptibility and detoxifying enzyme activities in field-collected resistant and susceptible strains of Tetranychus urticae. Pest Manag Sci 61(5):499–507
Van Leeuwen T, Vanholme B, Van Pottelberge S, Van Nieuwenhuyse P (2008) Mitochondrial heteroplasmy and the evolution of insecticide resistance: non-Mendelian inheritance in action. In: Proceedings of the national academy of sciences of the United States of America, vol 105, no 16, pp 5980–5985
Vollrath F (1999) Biology of spider silk. Int J Biol Macromol 24:81–88
Vollrath F, Köhler T (1996) Mechanics of silk produced by loaded spiders. Proc R Soc 263:387–391
Yano S (2008) Collective and solitary behaviors of Twospotted Spider Mite (Acari: Tetranychidae) are induced by trail following. Ann Entomol Soc Am 101(1):247–252
Acknowledgments
We are very grateful to Ferdinand Desneux of the ISV (Institut des sciences de la vie, UCL, Belgium) who gave microscope assistance and to Claude Remacle who allowed us to use the transmission electron microscope. Anne-Marie Faber provided fluorescent microscope assistance. Thanks to Louwrens R. Tiedt who gave us the micrographs of pedipalps. We thank Thomas Van Leeuwen who has supplied the T. urticae strain used in our experiments. We also thank Aina Astudillo Fernandez, Guillaume Le Goff and other members in our laboratory for valuable suggestions. This research was supported by Fonds pour la recherche dans l’Industrie et dans l’Agriculture (F.R.I.A.) of Belgium and by the Fund for Fundamental and Collective Research (FRFC, convention 2.4622.06). This paper is a publication BRC213 of the Biodiversity Research Center (Université catholique de Louvain).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Clotuche, G., Mailleux, AC., Deneubourg, JL. et al. The silk road of Tetranychus urticae: is it a single or a double lane?. Exp Appl Acarol 56, 345–354 (2012). https://doi.org/10.1007/s10493-012-9520-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10493-012-9520-6