
Abstract
The objective was to evaluate the animal thermal comfort indices from two integrated crop-livestock-forestry (ICLF) systems. For this, temperature–humidity index (THI), black globe temperature and humidity index (BGHI), and the radiant thermal load (RTL) were assessed. Two ICLF (ICLF-1 and ICLF-2) systems and one control system were established. On the ICLF systems, the arboreal component was the eucalyptus tree (Eucalyptus grandis × Eucalyptus urophylla; H13 clone), planted in simple wide-spaced rows. The ICLF-1 system had a tree spacing of 14 × 2 m with 357 trees per hectare, and the ICLF-2 had a tree spacing of 22 × 2 m with 227 trees per hectare. The control system had five scattered native trees per hectare, pertaining to Gochnatia and Dipteryx species. The forage component in all three systems was piatã-grass (Brachiaria brizantha cv. BRS Piatã). The experimental design was a randomized block in a sub-subplot design scheme with four replications. The presence of shade provided by the trees offered better conditions of animal comfort when compared with the condition of full sun. The ICLF-1 system, with higher tree density, provided better indicators for thermal comfort, THI, BGHI, and RTL when compared with the condition of full sun, while ICLF-2 was no different than ICLF-1 for BGHI.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The concern regarding environmental conditions of production systems has increased in the past half century, particularly related with those that affect animal welfare. The increasing public pressure and increased awareness of the importance of animal husbandry have triggered researches for new production strategies capable of meeting the principles of sustainability and animal welfare (Pires and Paciullo 2015). Thermal comfort is one of the issues raised in animal welfare, and is an essential condition for optimal productivity of grazing animals (Navarini et al. 2009). Climatic elements (e.g., air temperature, relative humidity, solar radiation and wind speed) are highly associated with thermal comfort. Brazil, because it is located in the intertropical zone, presents a high incidence of solar radiation throughout the year (Alves 2012). As a result, livestock raising on pastures is under great influence of weather conditions since animals are susceptible to slight climatic changes (Navarini et al. 2009).
Most of the animals that produce meat, milk, hide, wool and eggs are considered homeothermic animals because of their ability to control body temperature almost to a constant, varying within a narrow range (Silva 2000). These animals, when subjected to great solar radiation, high temperatures, and increased relative humidity, enter in a physiologic state known as heat stress. Under these conditions, in order to reestablish thermal neutrality, the animals trigger mechanisms of thermal regulation, among which are physiological responses (e.g., increase in heart rate, respiratory rate and metabolic rate), and behavioral responses, such as reduction in grazing time (Glaser 2003). As a consequence, the heat stress results in impairment of feed intake, growth and reproductive performance (Marques et al. 2006). To overcome this situation, producers can use natural shading, which is a tool used to improve animal welfare and animal thermal comfort by reducing the great levels of solar radiation and temperature, and improving air humidity, which enhance the performance of animals (Schütz et al. 2009; Gurgel et al. 2012; Baliscei et al. 2013).
A well designed integrated crop-livestock-forestry (ICLF) system is a good choice for producers willing to diversify their revenue sources without decreasing animal production, in early stages of tree development (Oliveira et al. 2014). These systems, which integrate trees, crops, pasture and animals in the same area on a timescale, are particularly indicated to enhance thermal comfort and animal performance because of the presence of shade from trees, contributing to the improvement of local microclimate and animal welfare (Thornton et al. 2009; Broom et al. 2013). Additionally, tree’s arrangements on ICLF systems contribute to alleviate the effects of extreme temperatures, reduce the impact of rain and wind, promoting comfort and serving as shelter for animals, in addition to improve soil and environment (Dias and Souto 2006). Several studies reported additional income when integrated systems are utilized. In Brazil, an integrated crop-livestock system that dealt with recycling of nutrients and power conversion, adopting manure treatment in intensified agricultural systems, had an effective contribution to improve the environment and the economic performance (Buller et al. 2015). In addition to economic advantages, other benefits arise from the use of trees in integrated systems such as carbon sequestration, especially when utilizing eucalyptus (Almeida et al. 2013). The ICLF systems help to mitigate greenhouse gas emissions as well as to accumulate soil C (Salton et al. 2014).
Considering the aforementioned, our objective was to evaluate animal thermal comfort indices from integrated agrosilvopastoral systems utilizing eucalyptus (Eucalyptus grandis × Eucalyptus urophylla) as the arboreal component and piatã-grass (Brachiaria brizantha cv. BRS Piatã) as the forage component in the Brazilian savannah.
Materials and methods
This study was conducted at the Embrapa Beef Cattle, in Campo Grande, Mato Grosso do Sul State, Brazil (54°37′W, 20°27′S, and 530 m of altitude). This location is geographically localized at the central portion of the state (which is located in the mid-west region of the country) in the neotropis zone of the biogeography region of savannah (Sandeville Jr. 2004). According to the Köppen-Geiger climate classification (Kottek et al. 2006), the Research Unit is located in the transition between Cfa and AW tropical humid, characterized by a regular rain distribution, with an average annual precipitation of 1560 mm, dry season occurring during the coldest months (May to August) and rainy season during the summer, with an average annual temperature of 23 °C.
Experimental area
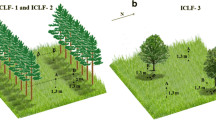
The establishment of the experimental area utilized in the present study was previously described by Oliveira et al. (2014). Briefly, the experimental area was established in 2008 and had 18 ha. Previously to the establishment of the ICLF systems, the area was subjected to herbicide desiccation, heavy disking, and sub-soiling, and had no trees. After liming and gypsum application, fertilization and soybean planting, the tree component (Eucalyptus grandis × Eucalyptus urophylla; H13 clone) was implanted in simple wide-spaced rows (displacement of −20.41°S and −54.71°W relative to the east–west axis). Two ICLF systems (ICLF-1 and ICLF-2) were established, differing on tree spacing and density (14 × 2 m with 357 trees per ha, and 22 × 2 m with 227 trees per ha, respectively). During the experimental period, the initial average height of the trees was 16 m and the final, 20 m. After tree planting, the forage component (piatã-grass; Brachiaria brizantha cv. BRS Piatã) was planted in the area. An integrated crop-livestock system (control; CON) was utilized in study and was composed of five scattered native trees (Gochnatia and Dipteryx species) per hectare, which is commonly found in ordinary production systems of the region. In this area, soybean planting was performed before the establishment of the forage component, in a similar way of the other systems. An aerial photography of the experimental site and the tree species present in the three systems are illustrated in Fig. 1.

Aerial photography of the experimental site and the representation of the tree species present in the integrated crop-livestock-forest (ICLF) systems (ICLF-1 and ICLF-2) and the control (CON) system
Regarding the arboreal component found in the three systems, the Eucalyptus urograndis, developed in Brazil from the crossing of E. urophyla and E. grandis, belongs to the Myrtaceae family. This species is characterized by its large size when it reaches its maturity, and has a high and straight trunk with smooth bark, gray or brown and persistent leaves (Lindon et al. 2001). On the other hand, the native species Gochnatia polymorpha belongs to the family Asteraceae, characterized by being a perennial tree, with simple white-tomentous leaves in the inferior face and matte in the superior face (Brandão et al. 2002). The species Dipteryx alata belongs to the Fabaceae (Leguminosae: Papillonoldeae) family, is a native from Bolivia and Colombia and is a perennial to slightly deciduous tree (Guarim Neto 1986) with compound leaves (Ferreira et al. 1998). During the evaluation period, both Gochnatia and Dipteryx trees have reached their maturity, presenting on average 11 and 14 m of height, respectively.
Thermal measurements
Starting in 2011, dry bulb, wet bulb and black globe temperatures, and wind speed data were measured from the paddocks to characterize the thermal conditions of each production system, during four consecutive days in July (winter), October (spring), December (summer), and April 2012 (autumn). Dry bulb temperature was measured using a digital thermo-hygrometer (model Perceptec1020, Perceptec Soluções e Tecnologia Ltda., São Paulo, Brazil) with data logger programmed to register temperature and humidity. Wet bulb temperature was measured using a visual reading psychrometric set (−10 to 50 °C scale; Incoterm, Porto Alegre, RS, Brazil); measurement of black globe temperature was achieved by using an adapted thermometer, as proposed by Souza et al. (2002). Wind speed was measured by using a portable digital anemometer (model HMM 489, Homis Controle e Instrumentação Ltda., São Paulo, SP, Brazil), in 3-min intervals, obtaining minimum and maximum velocity. All devices were placed at 1.5 m from the soil surface, which correspond to the center of mass of beef cattle (Navarini et al. 2009). Measurements were performed at 07:00, 09:00 and 11:00, corresponding to the morning hours, and 13:00, 15:00 and 17:00, corresponding to the afternoon hours (local time: GMT −04:00) on a transect perpendicular to the tree rows (Fig. 2). In each transect, two sampling points (A and B) were assessed: point A was located 2 m from the tree row and B in a central location between tree rows for ICLF-1 and 2, whereas for CON, point A was located 2 m from the tree and B in a central location of the paddock. Meteorological data were collected daily by a meteorological station (A702–INMET) distant approximately 4 km from the experimental area (Fig. 3).

Schematic diagram of sampling points (A and B) in the integrated crop-livestock-forest (ICLF) systems (ICLF-1, with 357 trees per ha and ICLF-2, with 227 trees per ha), and the control (CON) system (with five remaining native trees per hectare)

Weather data collected daily from a meteorological station (A702—Inmet) between July 2011 and June 2012. Tmax maximum temperature, Tmin minimum temperature, RHmin minimum relative humidity, *months of experimental evaluation
Shade disposition of ICLF-1 and ICLF-2 systems was measured during four consecutive days in each season from measurements between two tree rows in shaded areas using a field measuring tape at 07:00, 09:00, 11:00, 13:00, and 17:00.
Thermal comfort indices
From the microclimatic data obtained from each system, animal thermal comfort indices were calculated for each season (winter, spring, summer, and autumn). Temperature–humidity index (THI) was calculated using the equation as described by Thom (1958):
in which dbt is the dry bulb temperature and wbt is the wet-bulb temperature.
The black globe temperature and humidity index (BGHI) was calculated using the equation proposed by Buffington et al. (1981):
in which bgt is the black globe temperature.
Radiant thermal load (RTL) was calculated using the equation described in Esmay (1978):
in which σ is the Stefan–Boltzmann constant, 5.67 × 10−8 kg s−3 K−4(W m−2); MRT is the mean radiant temperature; ws is the wind speed (m s−1); and t is the air temperature.
Statistical analysis
The experimental design utilized was a randomized sub-subplot block, with four replications (paddocks). Main plot consisted of three systems (ICLF-1, ICLF-2, and CON), and subplot was the four seasons (spring, summer, autumn, and winter). For microclimate variables (dry bulb-, wet bulb- and black globe-temperature), sub-subplot was day hours (07:00; 09:00, 11:00, 13:00, 15:00, and 17:00), and for thermal comfort indices (THI, BGHI, and RTL), sub-subplot was day periods (morning and afternoon).
The microclimate variables (dry bulb-, wet bulb- and black globe-temperature), and percentage of shaded area data were analyzed using the non-parametric test of Kruskal–Wallis, and when having statistical significance, the Student–Newman–Keuls test was performed. Because there were two systems to compare the percentage of shaded area (ICLF-1 and ICLF-2), the Mann–Whitney test was employed. Analysis of data regarding THI, BGHI and RTL were achieved by ANOVA using the GLM procedure of SAS (SAS Inst., Inc., Cary, NC, USA; version 9.1) and means were separated for comparison by Tukey’s Studentized Test. Significance for all analyses was set at P ≤ 0.05.
Results
Thermal measurements
Maximum, minimum and average dry bulb-, wet bulb- and black globe-temperature data are depicted in Table 1. The maximum dry bulb temperature registered was 38.0 °C during the summer in the ICLF-2, close to what was registered in the CON system (37.9 °C). For wet-bulb temperature, the maximum value was registered during the autumn in the ICLF-2 system (26.4 °C); however, in terms of average value, the value recorded in the ICLF-2 was identical to CON (23.5 °C). During the summer, in the CON system, we observed the greatest black globe temperature registered throughout the experimental period (45.2 °C). Similarly, the CON system had the greatest wind speed during the autumn (5.1 m s−1). The average wind speed was 2.73 m s−1 in the systems.
Differences (P < 0.01) were observed among seasons for dry bulb-, wet bulb- and black globe-temperature (Fig. 4). Winter and summer had dry bulb-temperature of 32.1 and 31.0 °C, respectively, which were greater (P < 0.05) if compared with spring and autumn (24.6 and 23.7 °C, respectively). There were no differences (P > 0.05) between winter and summer, and between spring and autumn. For wet-bulb temperature, the average value recorded during the summer was greater (P < 0.05) than the value recorded during the autumn, followed by winter and spring (average values = 23.4, 21.4, 20.3, and 19.8 °C, respectively). These two last seasons had similar (P > 0.05) values for wet bulb temperature. Similarly to dry-bulb temperature, black globe temperature was greater (P < 0.05) during the summer (35.0 °C) and winter (34.8 °C) compared with autumn (27.6 °C) and spring (26.9 °C). There were no differences (P > 0.05) between winter and summer, and spring and autumn.

Average values of dry bulb (DBT), wet bulb (WBT) and black globe (BGT) temperatures during the four seasons. Different lowercase letters indicate that averages differ by the Student–Newman–Keuls test at P ≤ 0.05
Comparing the systems, we observed differences (P < 0.01) for wet bulb temperature (Fig. 5). In the ICLF-1, wet-bulb temperature was lesser (P < 0.01) among systems. Conversely, dry bulb- and black globe- temperatures did not differ (P > 0.05) among systems.

Average values of dry bulb (DBT), wet bulb (WBT) and black globe (BGT) temperatures in the integrated crop-livestock-forest (ICLF) systems (ICLF-1, with 357 trees per ha and ICLF-2, with 227 trees per ha), and the control (CON) system (with five remaining native trees per hectare). * Different lowercase letters indicate that averages differ by the Student–Newman–Keuls test at P ≤ 0.05
Dry bulb-, wet bulb- and black globe-temperatures did differ (P < 0.01) between day hours (Table 2). At 11:00, 13:00 and 15:00, dry bulb- and black globe-temperature were greater than 09:00 and 17:00, with least values registered at 07:00 (22.5 and 23.5 °C for dry bulb- and black globe-temperatures, respectively). The least wet bulb temperature was recorded at 07:00; and at 15:00 the greatest value was recorded (22.4 °C), not differing (P > 0.05) from 11:00 (21.9 °C) and 13:00 (22.0 °C).
Shade projection throughout the day on ICLF-1 and ICLF-2, in each season, can be visualized in Fig. 6. In both systems, we observed a larger shade area during the autumn and winter, concentrated within the central parts between tree rows. Conversely, during spring and summer, we observed smaller shading, with light concentrating between tree rows and shades closer to tree rows. Percentage of shade did differ among hours of the day (P = 0.02), seasons (P < 0.01), and systems (P = 0.04), as depicted in Table 3. At 07:00, 09:00 and 17:00, greater percentage of shaded area was observed compared with mid-day hours (11:00, 13:00, and 15:00). For seasons, less percentage of shaded area was observed during spring (32.3%) and summer (29.3%), whereas during the autumn (57.8%) and winter (62.4%) was greater. The greatest average shade projection was observed on ICLF-1 (51.9%) whereas ICLF-2 had an average shaded area of 39.0%.

Shade pattern of the integrated crop-livestock-forest (ICLF) systems (ICLF-1, with 357 trees per ha and ICLF-2, with 227 trees per ha), according to the displacement of the sun between 07:00 and 17:00
Thermal comfort indices
Thermal comfort indices varied according to day period (morning or afternoon; Table 4), with greater values observed during the afternoon (THI = 77.1; BGHI = 80.4; and RTL = 574.6 W m−2) and least values during the morning (THI = 74.0; BGHI = 76.8; and RTL = 539.5 W m−2) hours.
The least average value of THI was observed in the ICLF-1, intermediate values in ICLF-2, and the greatest value in CON, which presented the greatest averages in all observed hours (Fig. 7). In all systems, the greatest heat stress condition observed occurred at 13:00, with averages of 77.3, 78.5, and 79.4 for ICLF-1, ICLF-2, and CON, respectively. As expected, least values were observed in the early morning hour (07:00) with averages of 69.1, 69.7, and 71.0, respectively. Regarding systems, differences were observed (Table 4). The least value observed was in the ICLF-1 (74.9) and the greatest in the CON system (76.4), but ICLF-2 did not differ from ICLF-1 and CON systems. Differences were observed as a function of seasons (Table 4). The THI during the summer did not differ from winter, and autumn did not differ from spring. The greatest THI values were observed in the summer and winter, and the least values in the autumn and spring.

Temperature–humidity index (THI) of the integrated crop-livestock-forest (ICLF) systems (ICLF-1, with 357 trees per ha and ICLF-2, with 227 trees per ha), and the control (CON) system (with five remaining native trees per hectare) measured between 07:00 and 17:00
The BGHI data throughout the day in the systems are represented in Fig. 8. The least value was observed in the ICLF-1, intermediate values were observed in ICLF-2, and we observed the greatest averages in the CON system in all observed hours. Similarly to THI, the greatest heat stress condition observed occurred at 13:00, with averages of 80.6, 82.3, and 84.5, and the least values were observed in the early morning hour (07:00) with averages of 69.6, 69.8, and 73.3, for ICLF-1, ICLF-2, and CON respectively. There was a system effect on BGHI (Table 4). The CON system had greater value compared with ICLF-1 and ICLF-2 (80.4, 77.0, and 78.7, respectively). Similarly to THI, differences were observed as a function of seasons (Table 4). The BGHI during the summer did not differ from winter (greatest values), and autumn did not differ from spring (least values).

Black globe temperature and humidity index (BGHI) of the integrated crop-livestock-forest (ICLF) systems (ICLF-1, with 357 trees per ha and ICLF-2, with 227 trees per ha), and the control (CON) system (with five remaining native trees per hectare) measured between 07:00 and 17:00
Radiant thermal load data throughout the day in the evaluated systems are represented in Fig. 9. The least RTL value was observed in the ICLF-1 system, followed by ICLF-2. We observed the greatest values in the CON system in all evaluated hours, among systems. At 13:00 was the hour that provided greater heat condition, with average values of 579.3, 606.9, and 656.8 W m−2 for ICLF-1, ICLF-2, and CON, respectively, and the least values were observed in the early morning hour (07:00; 451.2, 441.8 and 505.1 W m−2 for ICLF-1, ICLF-2 and CON systems, respectively). A system × season interaction effect was observed (Table 5). No differences were observed among season in ICLF-1 and ICLF-2, with average values of 526.7 and 549.1 W m−2, respectively. The least value observed in the CON system was during the spring (549.2 W m−2), and the greatest values were in the summer (643.9 W m−2). During the autumn and spring, no differences among systems were observed. The lesser values were observed during the winter and summer in the ICLF-1 compared with the CON system, and no differences were observed for ICLF-2 relative to ICLF-1 and CON.

Radiant thermal load (RTL) in the integrated crop-livestock-forest (ICLF) systems (ICLF-1, with 357 trees per ha and ICLF-2, with 227 trees per ha), and the control (CON) system (with five remaining native trees per hectare) measured between 07:00 and 17:00
Discussion
Agroforestry practices such as ICLF systems are interesting because of their benefits, particularly regarding soil and animal components. On the perspective of animals, we observed that thermal comfort indices improved in ICLF systems because of a reduction of solar radiation because of the tree component of systems. Although we recorded dry bulb temperature above 35 °C (which is critical for Nellore cattle; Silva 2008) during the winter and summer in all systems, the thermal comfort indices were ameliorated in ICLF systems. Conversely, Baliscei et al. (2013), evaluating systems located in a lower parallel (22°44′ S, 52°28′ W, and 453 m of altitude), reported average dry bulb temperature of 24.1 and 24.4 °C in silvopastoral systems with double Eucalyptus tree row and ordinary pastures, respectively. The temperatures recorded in the present study are not within the range considered by Baêta and Souza (2010) as the best climate conditions for cattle, which are temperatures within 10 and 27 °C, relative air humidity within 60 and 70%, and wind speed between 1.38 and 2.22 m s−1. Moore et al. (2012) warned that high air temperature associated with high solar radiation and humidity provide an unfavorable thermal environment to animals.
One of the factors that helps in losing body heat is the wind. In the present study, although some wind speed values were greater than the range recommended by Baêta and Souza (2010), we noted that trees served as a windbreak in ICLF systems. In fact, a decrease in the average wind speed is noted as the arboreal density increases in these systems. Despite the benefits of the reduction of wind speed in the ICLF systems, attention should be given to densification, height and disbranching of the arboreal component, since the environment of very dense areas can be thermally uncomfortable to animals, particularly because of the increase of the temperature in the forest understory. The same attention should be given to shading because, although it provides greater thermal comfort to animals, it interferes with the growth of grasses, especially tropical species.
The differences observed among seasons for wet bulb temperature can be attributed to the greater rainfall during the summer, followed by autumn, winter and spring, as presented in Fig. 3. One of the hypotheses for the lower wet bulb temperature in ICLF-1 is related with the less amount of rain that reaches the soil because of the barrier formed by the trees. Both dry bulb and black globe temperatures had a similar pattern, with an increase of the values throughout the day and decrease at the end of the afternoon. The black globe temperature provides information of combined effects of air temperature, solar radiation and wind speed (Kelly and Bond 1971). In the present study, this parameter was regular although critical at some periods of the day according to the Ferreira et al. (2006) classification (from thermal comfort = 23 °C to severe heat stress conditions = 44 °C). It is noteworthy that dry bulb and black globe temperatures were expected to be greater during the summer than during the winter. However, we did not observe this because of the occurrence of atypical days during the winter (Fig. 4). At the same experimental area of this study, between July and September 2013, Karvatte Júnior et al. (2016) reported that the average air temperature was 43.5 °C (ICLF-1), 37.8 °C (ICLF-2), and 39.4 °C (CON) when trees were 26 m in height. The high air temperatures observed corroborate the need in providing to animals ways to mitigate the heat, even during the winter in regions that are similar to the site evaluated in the present study.
The reduction of shading between 11:00 and 15:00 is due to the rotation movement of the Earth. The greater values observed for dry bulb and black globe temperatures between these hours are because of the increase of the sunny areas. The great percentage of sunny areas in ICLF systems during the summer is because of the higher incidence of solar radiation caused by the summer solstice in the southern hemisphere, which coincided with the evaluation month (December). The greater percentage of shading in the ICLF-1 compared with ICLF-2 is because of the greater tree density (357 vs. 227 trees per hectare, respectively), since the distance between tree rows is shorter in the ICLF-1 than ICLF-2 (14 vs. 22 m, respectively).In forage-based cattle production systems, providing shading in an adequate proportion helps in the reduction of the heat load from solar radiation, shielding animals to heat stress (Kazama et al. 2008).
In all evaluated systems, the higher values of thermal comfort indices were observed at 13:00, except for THI in the ICLF-2. Kawabata et al. (2005) reported an increase of BGHI values until 12:00, corroborating the results observed in this study. Increments of 4.0, 4.5, and 6.1% were observed for THI, BGHT, and RTL, respectively, during the afternoon hours relative to the morning hours. The higher values observed for these indices are because of the fact that the maximum temperatures recorded were between 11:00 and 15:00. Similar pattern for these indices were also reported by Navarini et al. (2009) and Kawabata et al. (2005).
According to Baêta and Souza (2010), the THI can be used to classify the environment: values up to 70 define situation of thermal comfort; values ranging from 71 and 78 are considered critical; from 79 to 83, danger; and values above 83, emergency. Thus, the THI values observed in the current study for day hours are characterized as critical. The BGHI values, according to Baêta (1985), were classed by the National Weather Service as follows: comfort (up to 74); alert (74 to 79); danger (79 to 84); and emergency (above 84). Based on BGHI values observed in the present study, the ICLF-1 and ICLF-2 systems can be classed as alert and the CON system, as emergency. Based on the THI and BGHI results, the CON system presented the worst thermal comfort conditions to the animals. The ameliorated thermal condition found in the ICLF-1 and ICLF-2 can be explained by the presence of more trees compared with the CON system, which reduce the incidence of solar radiation in the forest understory, and consequent less warming of the surrounding air.
Characterizing the seasons based on the THI, autumn, winter and spring were classed as critical, and summer as danger. For BGHI, according to Baêta (1985), the environment was thermally classified by the National Weather Service as in danger situation in the winter and summer, and alert in the autumn and spring. According to Porfírio-da-Silva (2003), in the region of the Mato Grosso do Sul state, the pasture areas are under a climatic condition that determines median to severe caloric heat stress to animals without shelter from October to May. Elevated values for comfort indices in all seasons are a reflex of high temperatures observed.
The RTL is one of the main components in the study of energy balance and is used to evaluate the thermal conditions of an environment (Silva et al. 1990). The systems presented an increasing pattern for RTL values throughout the morning period, with a peak at 13:00, followed by a decrease in the afternoon period. This observation is also associated with dry bulb and wet bulb temperatures. Our values demonstrate a tendency of reduction and less variation on RTL throughout the seasons as the tree density increases. This fact is because of the presence of trees provides a more favorable microclimate to the animal, primarily because of the capacity of blocking the solar radiation. Similar results were reported by Baccari Junior (2001) who observed a reduction of 30–50% of the heat load on the animals when provided shading.
In conclusion, the arboreal component in cattle production systems provides better indices of thermal comfort because of the microclimate changes provided by shading. The denser system (ICLF-1, 357 trees per hectare) presented an improvement of 2.4% in the THI and 4.2% in BGHI. In addition, reductions of 12.4 and 16.3% in the RTL during the summer and winter seasons, respectively, compared with the CON system were observed. The ICLF-2 was no different than ICLF-1 for BGHI. Improvements in microclimate conditions, as observed in the ICLF systems, potentially provide better thermal comfort for animals. However, because integrated systems have great interdependence between components (tree, forage and animal), other factors should be considered, especially shade tolerance of the utilized forage. Therefore, the challenge is to scale the amount of proper shade in the system in order to equate the needs of each component.
References
Almeida RG, Andrade CMS, Paciullo DSC, Fernandes PCC, Cavalcante ACR, Barbosa RA, Valle CB (2013) Brazilian agroforestry systems for cattle and sheep. Trop Grassl 1:175–183
Alves FV (2012) O componente animal em sistemas de produção em integração. In: Bungenstab DJ (ed) Sistemas de integração lavoura-pecuária-floresta: a produção sustentável, 2nd edn. Embrapa, Brasilia
Baccari Junior F (2001) Manejo Ambiental da Vaca Leiteira em Climas Quentes. EDUEL, Londrina
Baêta FC (1985) Responses of lactating dairy cows to the combined effects of temperature, humidity and wind velocity in the warm season. Dissertation, University of Missouri
Baêta FC, Souza CF (2010) Ambiência em Edificações Rurais: Conforto Animal. Editora UFV, Viçosa
Baliscei MA, Barbosa OR, Souza W, Costa MAT, Krutzmann A, Queiroz EO (2013) Microclimate without shade and silvopastoral system during summer and winter. Acta Sci Anim Sci 35:49–56. doi:10.4025/actascianimsci.v35i1.15155
Brandão M, Laca-Buendia JP, Macedo JF (2002) Grevillea robusta A. Cunn. In: Árvores nativas e exóticas do Estado de Minas Gerais. EPAMIG, Belo Horizonte
Broom DM, Galindo FA, Murgueitio E (2013) Sustainable, efficient livestock production with high biodiversity and good welfare for animals. Proc R Soc B 280:2013–2025. doi:10.1098/rspb.2013.2025
Buffington DE, Collazo-Arocho A, Canton GH, Pitt D, Thatcher WW, Collier RJ (1981) Black globe-humidity index (BGHI) as comfort equation for dairy cows. Trans Am Soc Agric Eng 24:711–714. doi:10.13031/2013.34325
Buller LS, Bergier I, Ortega E, Moraes A, Bayma-Silva G, Zanetti MR (2015) Soil improvement and mitigation of greenhouse gas emissions for integrated crop–livestock systems: case study assessment in the Pantanal savanna highland, Brazil. Agric Syst 137:206–219. doi:10.1016/j.agsy.2014.11.004
Dias PF, Souto SM (2006) Análise de fatores aplicada na avaliação da influência de leguminosas arbóreas, nas características químicas de solo sob pastagem. Rev Univ Rural Sér da Vida 26:24–32
Esmay ML (1978) Principles of animal environment. Avi Publishing, Westport
Ferreira RA, Botelho SA, Davide AC, Malavasi MM (1998) Caracterização morfológica de fruto, semente, plântula e muda de Dipteryx alata Vogel—Baru (Leguminosae Papilionoideae). Cerne 4:73–87
Ferreira F, Pires MFA, Martinez ML, Coelho SG, Carvalho AU, Ferreira PM, Facury Filho EJ, Campos WE (2006) Parâmetros fisiológicos de bovinos cruzados submetidos ao estresse calórico. Arq Bras Med Vet Zootec 58:732–738. doi:10.1590/S0102-09352006000500005
Glaser FD (2003) Aspectos comportamentais de bovinos da raça Angus a pasto frente à disponibilidade de recursos de sombra e água para imersão. Master’s dissertation, Universidade de São Paulo, São Paulo
Guarim Neto G (1986) Plantas ornamentais de Mato Grosso. Boletim FBCN 21:105–115
Gurgel EM, Seraphim OJ, Silva IJO (2012) Método de avaliação bioclimática da qualidade da sombra de árvores visando ao conforto térmico animal. Energ Agric 27:20–34. doi:10.17224/EnergAgric.2012v27n2p20-34
Karvatte Júnior N, Klosowski ES, de Almeida RG, Mesquita EE, Oliveira CC, Alves FV (2016) Shading effect on microclimate and thermal comfort indexes in integrated crop-livestock-forest systems in the Brazilian Midwest. Int J Biometeorol 60:1–9. doi:10.1007/s00484-016-1180-5
Kawabata CY, Castro RC, Savastano Júnior H (2005) Índices de conforto térmico e respostas fisiológicas de bezerros da raça holandesa em bezerreiros individuais com diferentes coberturas. Eng Agríc 25:598–607. doi:10.1590/S0100-69162005000300004
Kazama R, Roma CFC, Barbosa OR, Zeoula LM, Ducatti T, Tesolin LC (2008) Orientação e sombreamento do confinamento na temperatura da superfície do pelame de bovinos. Acta Sci Anim Sci 30:211–216. doi:10.4025/actascianimsci.v30i2.4702
Kelly C, Bond TE (1971) Bioclimatic factors and their measurement. In: National Academic of Sciences (ed) A guide to environmental research on animals. National Academic of Sciences, Washington, pp 71–92
Kottek M, Grieser J, Beck C, Rudolf B, Rubel F (2006) World map of the Köppen-Geiger climate classification updated. Meteorol Z 15:259–263. doi:10.1127/0941-2948/2006/0130
Lindon F, Gomes H, Campos A (2001) Anatomia e morfologia externa das plantas superiores. Lidel, Lisbon
Marques JA, Caldas Neto SF, Groff AM, Simonelli SM, Corasa J, Romero L, Zawadski F, Araújo PF (2006) Comportamento de bovinos mestiços em confinamento com e sem acesso a sombra durante o período de verão. Campo Dig 1:54–59
Moore DA, Duprau JL, Wenz JR (2012) Short communication: effects of dairy calf hutch elevation on heat reduction, carbon dioxide concentration, air circulation, and respiratory rates. J Dairy Sci 95:4050–4054. doi:10.3168/jds.2012-5397
Navarini FC, Klosowski ES, Campos AT, Teixeira RA, Almeida CP (2009) Conforto térmico de bovinos da raça nelore a pasto sob diferentes condições de sombreamento e a pleno sol. Eng Agríc 29:508–517. doi:10.1590/S0100-69162009000400001
Oliveira CC, Villela SDJ, Almeida RG, Alves FV, Behling Neto A, Martins PGMA (2014) Performance of Nellore heifers, forage mass, and structural and nutritional characteristics of Brachiaria brizantha grass in integrated production systems. Trop Anim Health Prod 46:167–172. doi:10.1007/s11250-013-0469-1
Pires MFA, Paciullo DS (2015) Bem-estar animal em sistemas integrados. In: Alves FV, Laura VA, Almeida RG (eds) Sistemas agroflorestais: A agropecuária sustetável, 1st edn. Embrapa, Brasilia
Porfírio-da-Silva V (2003) Sistemas silvipastoris em Mato Grosso do Sul. Para quê adota-los? In: Proceedings ofthe Seminário Sistemas Agroflorestais e Desenvolvimento Sustentável. Embrapa Gado de Corte, Campo Grande, pp 1–13
Salton JC, Mercante FM, Tomazi M, Zanatta JA, Concenço G, Silva WM, Retore M (2014) Integrated crop-livestock system in tropical Brazil: toward a sustainable production system. Agric Ecosyst Environ 190:70–79. doi:10.1016/j.agee.2013.09.023
Sandeville E Jr (2004) A divisão natural das paisagens brasileiras. Paisagem Ambiente Ensaios 18:71–98
Schütz KE, Rogers AR, Cox NR, Tucker CB (2009) Dairy cows prefer shade that offers greater protection against solar radiation in summer: shade use, behavior and body temperature. Appl Anim Behav Sci 116:28–34
Silva RG (2000) Introdução à bioclimatologia animal. Nobel, São Paulo
Silva RG (2008) Biofísica ambiental: Os animais e seu ambiente. Funep, Jaboticabal
Silva IJ, Ghelfi Filho OK, Consiglero FR (1990) Materiais de cobertura para instalações animais. Eng Rural 1:51–60
Souza CF, Tinôco IFF, Baêta FC, Ferreira WPM, Silva RS (2002) Avaliação de materiais alternativos para confecção do termômetro de globo. Ciênc Agrotec 26:157–164
Thom EC (1958) Cooling degrees: day air-conditioning, heating and ventilating. Trans Am Soc Heat Refrig Air Cond Eng 55:65–72
Thornton PK, Van de Steeg J, Notenbaert A, Herrero M (2009) The impacts of climate change on livestock and livestock systems in developing countries: a review of what we know and what we need to know. Agric Syst 101:113–127. doi:10.1016/j.agsy.2009.05.002
Acknowledgements
The authors thank the Brazilian funding agencies CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior), for a scholarship granted to C.C. de Oliveira and P.G.M.A Martins (Grant no. 23038.008980/2013-90), and FUNDECT (Fundação de Apoio ao Desenvolvimento do Ensino, Ciência e Tecnologia do Estado de Mato Grosso do Sul), for financial support. In addition, the authors express appreciation to the staff from the Embrapa Beef Cattle Research Unit for assistance during this study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
de Oliveira, C.C., Alves, F.V., de Almeida, R.G. et al. Thermal comfort indices assessed in integrated production systems in the Brazilian savannah. Agroforest Syst 92, 1659–1672 (2018). https://doi.org/10.1007/s10457-017-0114-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-017-0114-5