
Abstract
The success of agroforestry in semi-arid areas depends on efficient use of available water and effective strategies to limit tree/crop competition and maximise productivity. On hillsides, planting improved tree fallows on the degraded upper section of bench terraces is a recommended practice to improve soil fertility while cropping continues on the lower terrace to maintain food production. This study examined the influence of tree fallows on soil water content (θ w ) and evaporation (E s ). Alnus acuminata Kunth (alnus), Calliandra calothyrsus Meissner (calliandra), Sesbania sesban L. (sesbania), a mixture of all three species, or sole crops (beans (Phaseolus vulgaris L.) or maize (Zea mays L.)) were grown on the upper terrace. The same sole crops were grown on the lower terrace. Four management regimes (unpruned, root, shoot and root + shoot pruned) were applied to the tree rows adjacent to the cropping area. Neutron probe and microlysimeter approaches were used to determine θ w and E s when the trees were c. 3.5 years old. Sesbania and alnus increased θ w by 9–18 % in the cropping area on the lower terrace but calliandra reduced θ w by 3–15 %. After heavy rain, E s comprised 29–38 % of precipitation in the tree-based treatments and 53 % under sole crops. Absolute values declined as rainfall decreased, but E s as a proportion of rainfall increased to 39–45 % in the tree-based treatments and 62 % for sole crops. Root + shoot pruning of alnus and the tree mixture increased θ w in the cropping area but had no significant effect in the other tree-based treatments. The results suggest that sesbania and alnus can be planted on smallholdings without compromising water supply to adjacent crops, whereas calliandra decreased water availability despite reducing E s . These results provide a mechanistic understanding of reported effects on crop yield in the same site.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Water supplies for agriculture are seriously threatened by global climate change (Gregory and Ingram 2000; Fischer et al. 2005). As two thirds of the World’s population is expected to experience water shortages by 2050 (Rosenzweig et al. 2004), it is vital to improve the efficiency with which available water is used for food production. Increasing populations in the African highlands have forced a move from traditional shifting cultivation to more intensive farming, although this has not been accompanied by mechanisation or application of fertiliser or irrigation (Swinkels et al. 1997; Ong et al. 2006, 2007), resulting in depletion of natural resources and declining per capita food production (Sanchez et al. 1997). The small size of land-holdings means that farmers cannot allocate separate areas for trees and crops. Agroforestry may be a viable option to sustain productivity while providing essential tree products and ecological services in areas such as SW Uganda where crop yields are <35 % of potential production and the shortfall of wood supply is c. 40 % (Siriri and Bekunda 2004; Siriri et al. 2010). Similar problems occur throughout the semi-arid and sub-humid tropics.
Traditional cropping systems often cannot fully utilise available rainfall due to losses by evaporation from the soil surface (E s ), runoff and drainage (Ong et al. 1996, 2006, 2007). As E s comprises 20–40 % of rainfall in East and West Africa (Wallace et al. 1999; Jackson and Wallace 1999; Wallace and Gregory 2002), this has major implications for crop production. Key factors affecting E s are soil moisture content, ground cover and microclimate (Wallace and Gregory 2002; Lin 2010). Strategies that manipulate these factors may be used to reduce E s and increase the proportion of rainfall available to crops (Lin 2010). Development of land use systems that use scarce resources efficiently is vital to improve food security as future climate change scenarios predict reduced or more erratic rainfall in sub-Saharan Africa (Wallace and Gregory 2002).
Integration of trees on cropland may improve productivity by: providing spatial and/or temporal complementarity of resource capture by trees and crops (Ong et al. 2006, 2007); increasing soil organic matter content, infiltration and water storage (Wallace 1996; Sun et al. 2008); improving soil physical properties and biological activity (Yamoah et al. 1986); and enhancing nutrient supplies through nitrogen fixation and reduced leaching and soil erosion (Sun et al. 2008). Agroforestry systems promoted in East Africa include improved fallows containing Sesbania sesban and rotational woodlots of Calliandra calothyrsus or Alnus acuminata (Siriri and Raussen 2003; Siriri et al. 2010). Planting trees on the degraded upper part of bench terraces subject to repeated down-slope cultivation and scouring by heavy rain is a recommended practice to improve fertility and provide valuable tree products while cropping continues on the more fertile middle and lower terrace (Raussen et al. 1999; Siriri and Raussen 2003; Siriri et al. 2010). Planting trees on contours reduces runoff and erosion and improves soil fertility under a wide range of climatic conditions (Sun et al. 2008). However, agroforestry does not always provide a solution, as competition with crops is common (Ong et al. 2006, 2007; Sun et al. 2008; Siriri et al. 2010).
Bench terraces create a matrix of conditions for crops as their concave shape limits productivity on the upper terrace due to limited soil depth, water retention and fertility, and causes waterlogging on the lower terrace after heavy rainfall. Crop yield on the upper terrace is much lower than on the more fertile middle and lower sections, and may contribute only 5 % of the total yield (Siriri et al. 2010). Some studies suggest that spatial or temporal separation of trees and crops may be used to limit competition (Cooper et al. 1996), but farmers report that detrimental interactions may still occur (Wajja-Musukwe et al. 2008; Sun et al. 2008). This is important as continued crop production on the middle and lower terrace is vital for food security as farmers await the benefits of trees grown on the upper terrace (Siriri et al. 2010). Effective strategies to limit competition and enforce complementarity are vital.
Schroth (1999) suggested two options, selection of trees with characteristics that encourage complementarity and management interventions that limit competition. However, characteristics that reduce competition by trees are not always consistent with their intended use by farmers, including timber production or revenue generation from greenhouse gas credits (TIST 2008). When farmers’ needs and ecological compatibility conflict, understanding and manipulation of the underlying processes are essential. Shoot and/or root pruning of trees may be used to control competition (Ong et al. 2006, 2007; Bayala et al. 2008; Siriri et al. 2010). Jones et al. (1998) reported that shoot pruning of Prosopis juliflora limited below-ground competition with sorghum, while Jackson et al. (2000) showed that this practice reduced water use by trees and improved recharge of the crop rooting zone. Chandrashekara (2007) recommended shoot pruning regimes and frequencies for 10 important tree species in Kerala, India to limit competition with understorey crops, while Wajja-Musukwe et al. (2008) showed that root pruning one side of tree rows in sub-humid Uganda had little effect on tree growth but reduced competition with adjacent crops; however, competition was increased on the unpruned side of the tree row. Previous studies in Uganda have shown that unpruned sesbania fallows on the upper terrace had little impact on crop yield on the lower terrace, but that shoot and/or root pruning of alnus and, especially, calliandra was needed to maintain crop yield (Siriri et al. 2010).
Although inclusion of trees in farming systems may alter microclimate in ways that reduce soil evaporation and offset rainfall losses due to canopy interception (Ong et al. 2006, 2007; Lott et al. 2009; Lin 2010; Siriri et al. 2010), Lott et al. (2009) concluded that competition for water negated the potential benefits of microclimatic amelioration for understorey maize. The influence of microclimatic amelioration associated with the closely spaced, fast growing trees used in improved fallows and rotational woodlots on E s and θ w is unknown. As the use of such systems by smallholder farmers is increasing in semi-arid and sub-humid areas, studies of effects on E s and θ w are essential to understand how these systems influence water use efficiency, crop yield and food security. This study aimed to: (1) determine the influence on E s and θ w of planting improved tree fallows on the degraded upper section of bench terraces while cropping continued on the middle and lower terrace; and (2) examine the effectiveness of root and shoot pruning in improving compatibility between trees and crops.
Materials and methods
Experimental design
The study was conducted at Kigezi High School, Kabale District, SW Uganda (1°15′S, 29°55′E, 1,850 m asl). The bimodal rainfall (c. 1,000 mm year−1) is greater and more evenly distributed during the long cropping season (September–February) than during the short cropping season (April–June). Most land is terraced to control runoff and erosion; terraces are typically 20 m wide, with a rise of c. 1.5 m between them, and are used for smallholder production of sorghum (Sorghum bicolor L.), maize (Zea mays L.), beans (Phaseolus vulgaris L.), peas (Pisum sativum L.), sweet potato (Ipomea batatas L.) and Irish potatoes (Solanum tuberosum L.; Fig. 1a). The soil was a haplic ferralitic sandy clay loam developed from phyllite parent material. Topsoil pH (0–15 cm) was 6.5, and clay content decreased from 37.4 to 27.1 % between the upper and lower terrace due to erosive transfer during heavy rain and repeated downhill cultivation using hoes; soil organic matter increased from 1.11 to 1.31 g kg−1. Mean bicarbonate EDTA extractable phosphorus and exchangeable potassium concentrations were respectively 27–36 mg kg−1 and 0.48–0.54 molc kg−1 (Siriri and Raussen 2003).

Schematic diagram showing variation in crop performance on bench terraces on hillslopes in SW Uganda: a sole crop and b trees grown on the upper terrace with crops on the middle and lower terrace. Circle location of measurements of soil evaporation and below-canopy solar radiation; filled triangle location of volumetric soil water content measurements
Trees were grown on the upper third of the terrace for 4 years, while cropping continued on the middle and lower parts of the terrace (Fig. 1b). Sole stands of Alnus acuminata Kunth (alnus), Calliandra calothyrsus Meissner (calliandra), Sesbania sesban (L.) Merr. var. sesban (sesbania), a mixture of all three, and a sole crop control treatment were grown on the upper terrace (6 m wide). Alnus and calliandra were planted in September 2000 and sesbania in March 2001 to ensure all tree species were all ready for harvest at the same time (Siriri et al. 2010). These species were chosen for their N-fixing capacity and ability to produce 24–27 t ha−1 of fuelwood and 30 t ha−1 of above-ground biomass under local climatic conditions (Siriri and Raussen 2003), and were planted in three rows at a density equivalent to 10,000 trees ha−1. A single row of each species was grown in the tree mixture; sesbania, the least competitive species, was located next to the cropping area, calliandra in the central row and alnus, believed to be the most competitive (Siriri and Raussen 2003; Siriri et al. 2010), was planted furthest from the cropping area.
An unbalanced split plot design containing three blocks was used (Siriri et al. 2010). Main treatment plots on the upper terrace (trees or sole crop; 26 m long × 6 m wide) were randomly allocated in each block. Four tree management sub-treatments (unpruned, root pruned, shoot pruned and root + shoot pruned) were randomly allocated within each main treatment using 5 × 6 m sub-plots. Main and sub-treatment plots were respectively separated by 4 and 2 m wide paths to minimise interference. Sole crops of maize (Zea mays L. cv. H622) or beans (Phaseolus vulgaris L. cv. K132) were grown on the middle and lower terrace (each 6 m wide; Fig. 1b) during the long and short cropping seasons respectively. Beans were planted at a 50 × 10 cm spacing and maize at 75 × 30 cm. A sole crop treatment was grown on the upper terrace for comparison with the tree-based treatments.
The tree management regimes were a compromise between effective control of competition and maximum production of woody biomass and green manure. Shoot pruning involved removing all branches from the lower third of the canopy of the tree row adjacent to the down-slope cropping area before each cropping season. Prunings were returned as green manure to the plots from which they came. Root pruning was achieved by digging and infilling trenches 0.5 m from the outer tree row to sever roots growing into the cropping area before each rainy season; these were 30 cm deep when the trees were young and 50 cm deep when they were over 3 years old. The former represents the depth achievable using hand hoes during normal field operations (Siriri et al. 2010); deeper pruning would have compromised tree establishment and growth.
Climatic conditions
Solar radiation, air temperature and atmospheric saturation deficit (SD) above the tree and crop canopies were recorded by an automatic weather station (BWS200, Campbell Scientific, Shepshed, UK). Rainfall (ARG100 tipping bucket raingauge), wind speed (RM Young Young Rain Sentry), wet and dry bulb air temperatures and atmospheric saturation deficit (CS215) and solar radiation (CS300 silicon pyranometer) were automatically measured and recorded.
Soil water evaporation (E s )
Microlysimeters were used to determine the substantial spatial variation in E s introduced by the integration of trees on bench terraces due to their simplicity and reliability. Microlysimeters comprise rigid enclosures containing small soil volumes (1–3 kg) which are placed in closely fitting pits in the soil and weighed daily. Lysimeters of the type described by Daamen et al. (1993) and modified as suggested by Jackson and Wallace (1999) were used. These comprised UPVC cylinders 160 mm in diameter and 100 mm deep. Their walls were perforated with 10 mm diameter holes to allow roots to explore the enclosed soil (Villalobos and Fereres 1990) and the lower end was chamfered to provide a cutting edge to ease installation. Before each set of measurements, the lysimeter cylinders were pushed into the soil and left for 4 days, during which vertical drainage was unimpeded. Undisturbed soil cores were removed by excavating the lysimeters to a depth of 20–30 mm below the base; excess soil was removed before sealing the base and the perforations in the lysimeter walls using waterproof tape to prevent exchange of water. Lysimeters were prepared between 0700 and 0800 h after overnight rain, or when precipitation ceased if rain fell during the day before being installed in pits (165 mm diameter × 90 mm deep) c. 1 m from where the soil cores were collected. The casing protruded 10 mm above the soil to prevent entry of runoff or extraneous soil.
Two lysimeters were installed in all replicates of the unpruned and root + shoot pruned sub-treatments of the four tree-based treatments and the sole crop on the upper terrace, and also in the cropping area on the lower terrace (Fig. 1), giving a total of 36 lysimeters. These were weighed twice daily (0700–0800 and 1700–1800 h) using a balance (0.1 g resolution, equivalent to 0.01 mm of water) powered by a generator. Soil in the lysimeters was replaced after each rainfall event. In the absence of rain, lysimeters were used for up to 7 days, as recommended by Daamen et al. (1993). Measurements were made between 14 March and 10 July 2004 during Periods i–iv shown in Fig. 2a to determine E s under both rainy and dry conditions. As E s from moist soil depends primarily on microclimatic conditions, total shortwave radiation was measured 0.5 m above the soil for all lysimeter locations (SKS1110 pyranometer, Skye Instruments, Llandrindod Wells, UK); incident solar radiation was obtained from the weather station.

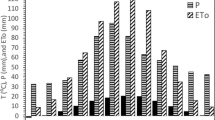
a Daily rainfall, maximum and minimum air temperature and b mean daily soil evaporation (E s ) during the 2004 short rains and subsequent dry season. Horizontal tramlines in a show periods when E s was measured and correspond to panels i, ii, iii and iv in (b). Error bars above each pair of treatment histograms show standard errors of the difference between treatment means (SED) for comparing the upper and lower terrace for specific land use treatments. The error bars for “SED land use” show SEDs for comparing E s between land use treatments on the upper (UT) and lower (LT) terrace sections. *, ** and *** denote P < 0.05, P < 0.01 and P < 0.001; NS no significant effect
Volumetric soil moisture content (θ w )
Neutron probes have been used to determine θ w for a wide range of soil types and land use systems. Their underlying theory, use and calibration are described by Bell (1987). Access tubes were installed in the upper, middle and lower terrace (Fig. 1) in the unpruned and root + shoot pruning sub-treatments of all four tree-based treatments and the sole crop control on the upper terrace (total of 81 tubes). The unpruned treatment represents current local practice and was expected to create the greatest competition for water with neighbouring crops, whereas the shoot + root pruning treatment was anticipated to be most effective in reducing competition. Three access tubes installed outside the experimental area on each of the upper, middle and lower terrace sections were used to calibrate the neutron probe (Wallingford Model IHII) against paired gravimetric measurements of θ w obtained using soil cores across the full range of θ w (Hillel 1998). Separate calibrations were constructed for the 0–30 cm and deeper soil horizons for each terrace position to account for the differing soil characteristics of topsoil and subsoil.
The base of the aluminium access tubes (5 cm diameter × 2 mm wall thickness) was sealed to prevent entry of ground water, while the top projected 5 cm above the soil and was covered by a metal cap to exclude rain and soil. Tubes were installed vertically after removing soil cores using an auger. Mean tube depth was 116.9, 139.7 and 174.6 cm on the upper, middle and lower terrace, reflecting the systematic variation in soil depth, and hence maximum rooting depth. Measurements were made at 15 cm intervals between 15 and 60 cm and at 20 cm intervals to the maximum depth of individual tubes. As neutron probe measurements at depths <15 cm may underestimate moisture content due to the loss of emitted radiation from the soil surface (Bell 1987), no consideration is given to the values obtained for the surface soil horizon in the “Discussion” section. θ w was measured at approximately weekly intervals between 29 November 2003 and 21 January 2004 and 13–26 March 2004 to give a total of 11 measurement dates.
Statistical analysis
The experimental design required split-plot analysis of variance as its unbalanced nature meant that the equivalent model had to be fitted using the residual maximum likelihood approach (REML), with estimated values and standard errors of the difference for treatment effects being taken from the fitted model (Siriri et al. 2010). Local variation in the performance of a cover crop of beans grown before the main experiment began was used as a covariate to remove any confounding influence on actual variation between treatments. Species and pruning regime represented the main and sub-treatments and the treatment structure was covariate + main treatment × sub-treatment; the covariate was yield from the cover crop. All analyses were carried out using Genstat 8 (Release 8.1) software. Standard errors of the difference between means (SED) and standard errors of the mean (SEM) are shown; significance was assumed at P ≤ 0.05.
Results
Climatic conditions
Daily maximum temperature was generally lower and minimum temperature much higher during the 2004 short rains (March–May) than during the dry season (June–August; Fig. 2a) due to the greater radiative exchange associated with limited cloud cover during the latter period. Mean daily maximum and minimum temperatures were respectively 24.2 and 11.7 °C. SD at 1500 h rarely reached 1.5 kPa during the rainy season, but often exceeded 2 kPa during the later stages of the dry season (data not shown). SD at 0800 h was generally <0.2 kPa during the rainy season and rarely reached 0.5 kPa, even in the dry season. The relatively low air temperature and SD values reflect the humid environment of tropical highland areas such as Kabale. Rainfall was well distributed during the rainy season, with seven events >20 mm (Fig. 2a). Little rain fell during the dry season.
Solar radiation
Figure 3a shows the mean diurnal timecourses for total short wave radiation measured above the tree canopy and 0.5 m above the soil on the upper terrace for the sole crop and all unpruned tree treatments in March 2004, when alnus and calliandra were 41 months old and sesbania was 35 months old. Mean below-canopy values for the tree-based systems ranged from 45 % of the above-canopy value (alnus) to 59 % (sesbania), and were much lower than in the sole bean crop (74 %; P < 0.001). Cumulative shortwave radiation under the tree canopies over the same period was between 29 % (sesbania) and 56 % (alnus) lower than in sole crops on the upper terrace (P < 0.001; Fig. 3b). There were no significant treatment differences in the cropping zone on the lower terrace, although values were 12–25 % lower in the tree-based treatments than when sole bean crops were grown on the upper terrace. The influence of terrace position was significant (P < 0.01) as cumulative radiation under the sole bean crop on the upper terrace was approximately double that in the alnus, calliandra and tree mixture treatments and significantly greater than that of the sole crop on the lower terrace. Although no significant treatment effects were detected on the lower terrace, cumulative shortwave radiation under the sole crop was lower than on the upper terrace, whereas the reverse applied for alnus, calliandra and the tree mixture, reflecting treatment differences in canopy structure and shading intensity.

a Mean diurnal timecourses for above- and below-canopy short wave radiation in the unpruned tree treatments and sole bean crop during March 2004 and b cumulative below-canopy short wave radiation. In a SED1 and SED2 respectively show standard errors of the difference between treatment means and the treatment × time interaction; in b SED1, SED2 and SED3 show values for comparing treatment means for the upper terrace, lower terrace and values for both terrace positions. *, ** and *** denote P < 0.05, P < 0.01 and P < 0.001; NS no significant effect
Soil evaporation (E s )
Measurements during a period when rainfall and the diurnal temperature range varied greatly showed that E s differed between land use treatments on the upper terrace and also in the cropping area on the lower terrace, particularly after high rainfall (Fig. 2a, b). During Period 1 (15–24 March 2004; Fig. 2bi), E s was greatest in the sole crop and lowest in the alnus treatment for both terrace positions (P < 0.001). When mean values for the upper and lower terrace are considered, 53 % of rainfall received by the sole crop was lost as E s , compared to 29–40 % in the tree-based treatments (P < 0.001). Absolute E s values were lower during Period 2 (14–21 May 2004; Fig. 2bii) but, when expressed as a proportion of rainfall, E s increased to 62 % in the sole crop and 39–53 % in the tree-based treatments. Values on the upper terrace were higher for the sole crop than in all other treatments (P < 0.001). Almost no rainfall occurred during Periods 3 and 4 (23–30 June 2004 and 7–14 July 2004; Fig. 2a), resulting in a further decline in E s (Fig. 2biii, iv). Soil evaporation was similar during both periods and did not differ between land use treatments.
The extent of the differences in E s between terrace positions varied between treatments. During Period 1, E s was greater in the cropping area on the lower terrace than on the upper terrace in the alnus, sesbania and tree mixture treatments (P < 0.01). A similar, but non-significant, trend was apparent for the calliandra and the sole crop treatments. During Period 2, the presence of trees on the upper terrace reduced E s by c. 50 % relative to the lower terrace (P < 0.001), but the difference between terrace positions was again not significant for the sole crop. During Period 3, E s was again lower on the upper terrace in the alnus and tree mixture treatments (P < 0.05). During Period 4, E s was lower on the upper terrace in the calliandra, sesbania and sole crop treatments (P < 0.01, P < 0.05 and P < 0.001), but did not differ significantly for the alnus and tree mixture treatments, in contrast to Period 3.
Volumetric water content (θ w )
Rainfall during the 7–9 day period preceding each measurement date varied between 0 and 102 mm, whereas mean daily temperature and relative humidity showed much smaller variation, ranging from 17.1 to 19.4 °C and 68 to 84 % (Table 1). Table 2 illustrates the increase in soil depth between the upper and lower terrace, which is typical of bench terraces in SW Uganda. On the upper terrace, θ w tended to be greater under sesbania than in the sole crop treatment for all soil depths, although this was significant only for the 30–45 cm horizon (P < 0.05). None of the other tree-based treatments showed a significant difference relative to the sole crop for any horizon. On the middle terrace, θ w was greater in the 60–80 and 80–100 horizons of the sesbania treatment than in the sole crop (P < 0.001). Values for calliandra and the tree mixture did not differ from the sole crop except in the 80–100 horizon, where θ w was greater in the calliandra and lower in the tree mixture treatment than in the sole crop (P < 0.05). On the lower terrace, θ w was greater in the alnus treatment than in the sole crop for all depths between 15 and 60 cm (P < 0.01–P < 0.001). Values for the other tree-based systems did not differ significantly from the sole crop.
The extent of the increase in mean θ w for the entire soil profile varied between treatments depending on terrace position (Table 2). Mean θ w was greatest in the alnus and sesbania treatments on the upper terrace (P < 0.001), and in the sesbania treatment on the middle terrace (P < 0.001), followed sequentially by the alnus, sole crop, calliandra and tree mixture treatments. On the lower terrace, θ w was again greatest for alnus (P < 0.001), but did not differ between the other treatments. Values for all terrace positions were 7–15 % greater for alnus than for the sole crop, and 14–18 % greater on the upper and middle terrace for sesbania (P < 0.001). Values for calliandra and the tree mixture were 3–15 % lower than in the sole crop.
Mean θ w values increased with depth (P < 0.01–P < 0.001) and were greater after ‘high’ rainfall (>20 mm; five events; Fig. 4b, d, f) than after ‘low’ rainfall (<20 mm; six events; Fig. 4a, c, e; P < 0.01). The influence of land use treatment was also greater after high rainfall for all terrace positions (P < 0.001). Values for θ w in the surface 60 cm of the profile were generally greatest for alnus and sesbania on the upper and middle terrace, and for alnus on the lower terrace when rainfall was >20 mm (P < 0.001; Fig. 4b, d, f), but not when rainfall was <20 mm (Fig. 4a, c, e). The results suggest that sesbania and, especially, alnus facilitate soil rewetting during periods of high rainfall; θ w tended to be lowest in the tree mixture and calliandra treatments.

Profiles of volumetric soil water content (θ w ) in the unpruned tree treatments and sole bean crop for the upper (a, b), middle (c, d) and lower (e, f) terrace following periods of low (a, c, e) or high (b, d, f) rainfall (<20 mm and >20 mm respectively). g Mean θ w values for the entire profile. In a–f, SED1, SED2 and SED3 respectively show values for comparing land use systems, soil depths and the land use system × treatment interaction. *, ** and *** denote P < 0.05, P < 0.01 and P < 0.001; NS no significant effect
Mean θ w values for the entire soil profile are summarised in Fig. 4g for all main treatments, terrace positions and sampling dates within each rainfall category. The significant treatment × rainfall interaction on the upper terrace (P < 0.05) indicates that the influence of rainfall varied among treatments; thus, θ w was lower in the alnus treatment than in the sesbania treatment when rainfall was <20 mm (P < 0.001), but was similar when rainfall was >20 mm. Soil water content was between 14 % (tree mixture) and 53 % (alnus) greater in the tree-based treatments when rainfall >20 mm, compared to 9 % in the sole crop.
No significant treatment × rainfall interaction was detected on the middle terrace, where mean θ w was greatest in the sesbania treatment after low rainfall (P < 0.001; Fig. 4g), but did not differ between the other tree-based treatments. When rainfall was >20 mm, θ w was greatest in the sesbania and alnus treatments (P < 0.001), in which values were 12–36 % greater than after low rainfall. There was again a significant treatment x rainfall interaction on the lower terrace (P < 0.05), where mean profile θ w differed little between treatments when rainfall was <20 mm but was much greater in the alnus treatment when rainfall was >20 mm (P < 0.001). Mean θ w in the tree-based treatments was between 6 % (tree mixture) and 50 % greater (alnus) after high rainfall compared to 10 % in the sole crop treatment.
The effect of root + shoot pruning the trees on the upper terrace on mean profile θ w within the cropping area on the lower terrace was greatest for the tree mixture (Table 3), in which θ w was increased by c. 43 % at distances of 2 and 6 m from the tree line compared to unpruned trees (P < 0.001). Pruning of alnus increased θ w by 14 % 2 m from the tree line (P < 0.05), but had no effect at 6 m; pruning of calliandra and sesbania had no detectable effect at either distance.
Discussion
Tree growth and potential competitive impact
Mean tree height and diameter at breast height (dbh) 36 months (alnus, calliandra) or 30 months after planting (sesbania) were respectively 7.1 m and 7.2 cm, 6.8 m and 5.3 cm, and 5.1 and 4.4 cm for unpruned alnus, sesbania and calliandra trees (Siriri et al. 2010). Tree height and dbh were significantly reduced by root + shoot pruning only in alnus, by 15 and 29 % respectively. The trees were therefore sufficiently well established to compete strongly with adjacent crops for above- and below-ground resources unless subjected to appropriate management practices such as root and/or shoot pruning. The presence of unpruned trees is likely to have influenced soil water balance in the adjacent cropping area both directly by abstracting soil moisture and indirectly through microclimatic modifications.
Soil evaporation (Es)
Trees influence E s through effects on microclimate and soil water content (Ong et al. 1996, 2006, 2007; Otengi et al. 2007; Lin 2010). The microclimatic factors most likely to be modified are solar radiation receipts at ground level and wind speed (Wallace 1996; Otengi et al. 2007). However, aerodynamic factors such as wind speed are less important in the relatively closed canopies provided by well-established improved tree fallows and rotational woodlots (Ritchie 1972; Wallace and Gregory 2002), with the result that solar radiation is the major factor governing first stage evaporation following rainfall. In the present study, irradiance measured 0.5 m above the soil was reduced by 29–56 % in the tree-based treatments relative to the sole crop treatment on the upper terrace, and by 12–25 % in the cropping zone on the lower terrace (Fig. 3). The results for the upper terrace are consistent with Jackson and Wallace (1999), who found that net radiation, the primary driver of soil surface evaporation, was reduced by ≤65 % in a linear agroforestry system at Machakos in semi-arid Kenya, where incident radiation is generally greater than in the humid African highlands examined here. The variation between tree-based systems reflects their differing canopy structures; thus, the denser canopy of alnus reduced below-canopy irradiance to a much greater extent than the more open canopy of sesbania (Fig. 3).
These differences were reflected by effects on E s (Fig. 2b), particularly after periods of high rainfall when this process depends primarily on the energy balance at the soil surface, whereas E s during periods of low rainfall (Fig. 2, Periods 2–4) is determined mainly by soil hydraulic properties (Hillel 1998). The ability of tree-based systems to retain soil moisture is potentially important in affecting E s ; thus, the increase in E s as a proportion of rainfall between Periods 1 and 2 was greatest for alnus (from 29 to 45 %), possibly because slower depletion of soil moisture during Period 1 following relatively high rainfall increased water supplies to maintain soil evaporation during the drier Period 2. Raussen et al. (1999) reported that soil hydraulic conductivity under alnus woodlots was more than twice that of continuous cropping systems, thereby enhancing infiltration of rainfall.
The mean reduction in E s in the tree-based treatments relative to sole crops during the rainy season (Periods 1 and 2), ranging between 20 % (tree mixture) and 36 % (alnus), was much greater than during the dry season (Periods 3 and 4) when E s was similar in all treatments (Fig. 2b). The reductions in E s in the tree-based systems during the rainy season are greater than reported for a linear agroforestry system containing Grevillea robusta in Kenya, where the mean decrease relative to sole maize was 16 % (Jackson and Wallace 1999). This contrast may have occurred because differences in planting arrangement (linear vs. block planting) influenced the intensity and extent of shading. A study of a mulched contour hedgerow system containing maize and cowpea (Vigna unguiculata L.) in semi-arid Kenya showed a reduction in E s of <9 % relative to bare soil (Kinama et al. 2005). Expressed as a percentage of rainfall, E s was nevertheless high under unmulched maize/senna (Senna spectabilis (DC.) H.S. Irwin and R.C. Barneby) and grass strip/maize intercrops (60 and 65 % respectively), suggesting that the improved fallow/rotational woodlot systems examined here were more effective in reducing E s than linear agroforestry systems. Moreover, spatial variation in E s in the present study appeared to be greater than in linear agroforestry systems as the microclimatic changes induced by growing trees on the upper terrace reduced E s by up to 30 % on the cropped lower terrace at distances of up to 6 m from the tree line during Period 1 (Fig. 2b). The similarity of the E s values obtained when sole crops were planted on both the upper and lower terrace suggests that variation in leaf area index associated with the fertility gradient was too small to affect soil evaporation, implying that the significant reduction in E s on the lower terrace in the tree-based treatments resulted from microclimatic modifications caused by the presence of trees on the upper terrace. The linear agroforestry system described by Jackson and Wallace (1999) represents an intermediate scenario as the trees strongly influenced E s at a distance of 0.3 m from the tree line, but not at 2.5 m.
Volumetric soil moisture content (θ w )
Factors influencing the capacity of tree-based systems to store water include infiltration, soil water evaporation, abstraction by trees and crops, organic matter content and textural characteristics (Ong et al. 2006, 2007). However, the limited literature regarding the importance of these factors for the species examined here precludes comparison with previous studies of soil water storage and microclimate in agroforestry systems, as most have focussed on deep-rooted tree species (Jackson et al. 2000; Lott et al. 2003; Radersma and Ong 2004; Lott et al. 2009) and have rarely examined the short-lived shrubs that are often used in fallows and rotational woodlots. The presence of alnus and sesbania increased θ w relative to the sole crop control treatment on the upper terrace, but a similar effect was not found for the calliandra and tree mixture treatments (Table 2). This observation reflects the trend for E s , which was lowest for alnus (Fig. 2b), perhaps because its dense canopy and more intense litterfall reduced both E s and runoff. The dense undergrowth in the sesbania treatment may have induced similar effects, increasing mean profile values for θ w . Similar trends were apparent on the middle terrace (Table 2), where mean profile θ w was greatest in the alnus and sesbania treatments, although there was no evidence of the further increase in θ w between the middle and lower terrace that had been anticipated in view of the greater soil depth at the latter location (Fig. 1; Table 2). This observation suggests that the ability of trees grown on the upper terrace to modify soil hydrological properties in ways that improve water storage on the upper and middle terrace does not extend to the lower terrace.
The observed beneficial influence of sesbania contrasts with reports that θ w in the topsoil was lower in improved fallows containing sesbania than under sole maize (Hartemink et al. 1996). This effect was attributed to aggressive water abstraction by sesbania, whereas water conservation by sesbania in the present study appeared to outweigh losses resulting from abstraction by roots. However, although E s was lower in the calliandra treatment than under sole crops (Fig. 2b), mean profile θ w values were similar in both treatments following periods of both high and low rainfall (Fig. 4g), suggesting that greater lateral extension of its roots into the cropping area compared to the other tree species examined created a more extensive spatial hydrological influence. Siriri et al. (2010) noted that calliandra depressed crop yield for distances of up to several metres from the tree line, supporting evidence that the roots of this species are highly versatile and may extend over considerable lateral and vertical distances (Hairiah et al. 1992). Inclusion of calliandra in the tree mixture may have been responsible for the significant reduction in θ w relative to the sole crop control treatment, supporting the supposition that, although E s was reduced (Fig. 2b), the water saved was absorbed by its extensive root system. Nevertheless, calliandra is widely used for fodder by farmers in Kabale despite its competitiveness with crops (Nyeko et al. 2004).
When rainfall was <20 mm during the week preceding measurements, mean profile θ w was greater in the tree-based treatments than in the sole crop control only for sesbania (Fig. 4g), perhaps because transpiration was lower than in the other tree species, or because its bushy growth habit reduced E s . When rainfall was >20 mm, mean θ w was greatest in the sesbania and, especially, the alnus treatments, perhaps because the mulching effect of the greater litter deposition increased infiltration. A further possibility is that the misty conditions encountered in the morning during the rainy season due to the low air temperature and high humidity allowed trees to trap additional moisture in the form of dew on their canopy, in a manner analogous to cloud forests; some species may be more effective than others in channelling this source of moisture to the soil due to differences in crown architecture.
The observation that root + shoot pruning of alnus and the tree mixture increased θ w on the lower terrace, but had no detectable effect in the calliandra and sesbania treatments (Table 3) reflects the finding that pruning the latter two species did not improve crop performance, although for different reasons (Siriri et al. 2010). The lack of any crop yield response to root + shoot pruning sesbania during five cropping seasons when beans or maize were grown substantiates the absence of any adverse effect of unpruned trees on θ w in the adjacent cropping area; indeed, the presence of unpruned sesbania on the upper terrace increased values for θ w in the cropping area, whereas unpruned calliandra trees competed strongly for water and reduced θ w on the lower terrace and the yield of bean and maize crops by c. 40 % over six cropping seasons during which rainfall varied greatly (Siriri et al. 2010). The pruning intensity used here may have been insufficient to control competition effectively; thus, a more extreme management regime than annual root pruning to 30 cm depth when the trees were young and 50 cm when they reached 3 years of age may be required for calliandra.
Conclusions
Previous studies of the effectiveness of root and/or shoot pruning in controlling competition by tree fallows grown on the upper section of bench terraces in Uganda showed that unpruned sesbania had little impact on crop yield on the lower terrace, whereas pruning of alnus and, especially, calliandra was vital to maintain crop yield (Siriri et al. 2010). The present study has revealed that some tree species have beneficial effects on E s and θ w in the adjacent cropping area and that the effects of pruning on crop yield were associated with reductions in E s and increases in θ w . The presence of sesbania or alnus on the upper terrace increased θ w on both the upper and lower terrace, whereas calliandra tended to reduce θ w for all terrace positions; the presence of trees greatly reduced E s compared to sole crops following periods of high rainfall. Root + shoot pruning of the tree mixture and alnus increased θ w in the cropping area compared to unpruned trees, but had no significant effect in the other tree-based treatments. Sesbania and alnus may be incorporated into smallholdings without compromising water supplies to adjacent crops, but the extensive lateral rooting of calliandra deprived adjacent crops of water even though E s was reduced.
The use of tree fallows on the upper section of bench terraces is recommended for soil fertility improvement and production of valuable tree products on steep hillslopes in East Africa, and such approaches may be more widely applicable throughout the semi-arid and sub-humid tropics. The key is to identify appropriate species and management practices that enhance crop yield on the lower terrace while enabling tree products to be harvested from the upper terrace.
References
Bayala J, Ouedraogo SJ, Teklehaimanot Z (2008) Rejuvenating trees in agroforestry systems for better fruit production using crown pruning. Agrofor Syst 72:187–194
Bell JP (1987) Neutron probe practice. Report 19, 3rd edn. Institute of Hydrology, Wallingford, 29 p
Chandrashekara UM (2007) Effects of pruning on radial growth and biomass increment of trees growing in homegardens of Kerala, India. Agrofor Syst 69:231–237
Cooper PJM, Leakey RRB, Rao MR, Reynolds L (1996) Agroforestry and the mitigation of land degradation in the humid and sub-humid tropics of Africa. Exp Agric 32:235–290
Daamen CC, Simmonds LP, Wallace JS, Laryea KK, Sivakumar MVK (1993) Use of microlysimeters to measure evaporation from sandy soils. Agric For Meteorol 65:159–173
Fischer G, Shah M, Tubiello FN, van Velthuizen H (2005) Socio-economic and climate change impacts on agriculture: an integrated assessment, 1990–2080. Phil Trans Roy Soc B 360:2067–2083
Gregory PJ, Ingram JSI (2000) Global change and food and forest production: future scientific challenges. Agric Ecosyst Environ 82:3–14
Hairiah K, van Noordwijk M, Santosa B, Syekhfani MS (1992) Biomass production and root distribution of eight trees and their potential for hedgerow intercropping on an ulitisol in Southern Sumatra. Agrivita 15:54–68
Hartemink AE, Buresh RJ, Jama B, Janssen BH (1996) Soil nitrate and water dynamics in Sesbania fallow, weed fallows and maize. Soil Sci Soc Am J 60:568–574
Hillel D (1998) Environmental soil physics. Academic Press, San Diego 771 p
Jackson NA, Wallace JS (1999) Soil evaporation: measurements in an agroforestry system in Kenya. Agric For Meteorol 94:203–215
Jackson NA, Wallace JS, Ong CK (2000) Tree pruning as a means of controlling water use in an agroforestry system in Kenya. For Ecol Manag 126:133–148
Jones M, Sinclair FL, Grime VL (1998) Effect of tree species and crown pruning on root length and soil water content in semi-arid agroforestry. Plant Soil 201:197–207
Kinama JM, Stigter CJ, Ong CK, Ng’ang’a JK, Gichuki FN (2005) Evaporation from soils below sparse crops in contour hedgerow agroforestry in semi-arid Kenya. Agric For Meteorol 130:149–162
Lin BB (2010) The role of agroforestry in reducing water loss through soil evaporation and crop transpiration in coffee agroecosystems. Agric For Meteorol 150:510–518
Lott JE, Khan AAH, Black CR, Ong CK (2003) Water use in a Grevillea robusta-maize overstorey agroforestry system in semi-arid Kenya. For Ecol Manag 180:45–59
Lott JE, Ong CK, Black CR (2009) Understorey microclimate and crop performance in a Grevillea robusta-based agroforestry system in semi-arid Kenya. Agric For Meteorol 149:1140–1151
Nyeko P, Stewart J, Franzel F, Barkland P (2004) Farmers’ experiences in the management and utilization of Calliandra calothrysus, a fodder shrub in Uganda. Agricultural Research and Extension Network (AgREN) Publication 140
Ong CK, Black CR, Marshall FM, Corlett JE (1996) Principles of resource capture and utilization of light and water. In: Ong CK, Huxley PA (eds) Tree-crop interactions in agroforestry systems: a physiological approach. CAB International, Wallingford, pp 73–158
Ong CK, Black CR, Muthuri CW (2006) Modifying forests and agroforestry for improved water productivity in the semi-arid tropics. CAB Rev Perspect Agric Vet Sci Nutr Nat Resour 65:1–19
Ong CK, Anyango S, Muthuri CW, Black CR (2007) Water use and water productivity of agroforestry systems in the semi-arid tropics. Ann Arid Zone 46:84–255
Otengi SBB, Stigter CJ, Ng’anga JK, Liniger H (2007) Soil moisture and its consequences in a six year old hedged agroforestry demonstration plot in semi-arid Kenya for two successive contrasting seasons. Afr J Agric Res 2:89–104
Radersma S, Ong CK (2004) Spatial distribution of root length density and soil water in linear agroforestry systems in sub-humid Kenya: implications for agroforestry models. For Ecol Manag 188:77–89
Raussen T, Siriri D, Ong C (1999) Trapping water, producing wood and improving yields through rotational woodlots on degraded parts of bench terraces in Uganda. East Afr Agric For J 65:85–93
Ritchie JT (1972) Model for predicting evaporation from a row crop with incomplete cover. Water Resour Res 8:1204–1213
Rosenzweig C, Strzepek KM, Major DC, Iglesias A, Yates DN, McCluskey A, Hillel D (2004) Water resources for agriculture in a changing climate: international case studies. Glob Environ Change 14:345–360
Sanchez PA, Shepherd KD, Soule MJ, Place FM, Buresh RJ (1997) Soil fertility replenishment in Africa: an investment in natural resource capital. In: Buresh RJ, Sanchez PA, Calhoun PG (eds) Replenishing soil fertility in Africa. Special Publication 51. Soil Science Society of America, Madison, pp 1–46
Schroth G (1999) A review of below-ground interactions in agroforestry, focusing on mechanisms and management options. Agrofor Syst 43:5–34
Siriri D, Bekunda MA (2004) Soil fertility management in Uganda: the potential of agroforestry. In: Proceedings of Second National Agroforestry Workshop, 10–14 Sep 2001, Mukono. ICRAF, Nairobi, pp 29–31
Siriri D, Raussen T (2003) Agronomic and economic potential of improved fallows on scoured terrace benches in the humid highlands of Southwestern Uganda. Agric Ecosyst Environ 95:359–369
Siriri D, Ong CK, Wilson J, Boffa JM, Black CR (2010) Tree species and pruning regime affect crop yield on bench terraces in SW Uganda. Agrofor Syst 78:65–77
Sun H, Tang Y, Xie J (2008) Contour hedgerow intercropping in the mountains of China: a review. Agrofor Syst 73:65–76
Swinkels RA, Franzel S, Shepherd KD, Ohlsson E, Ndufa JK (1997) The economics of short rotation improved fallows: evidence from areas of high population in western Kenya. Agric Syst 55:99–121
TIST (2008) Planting trees and improving agriculture for better lives. http://www.tist.org/
Villalobos FJ, Fereres E (1990) Evaporation measurements beneath corn, cotton and sunflower canopies. Agron J 82:1153–1159
Wajja-Musukwe T-N, Wilson J, Sprent JI, Ong CK, Deans JD, Okorio J (2008) Tree growth and management of Ugandan agroforestry systems: effects of root pruning on tree growth and crop yield. Tree Physiol 28:233–242
Wallace JS (1996) The water balance of mixed tree-crop systems. In: Ong CK, Huxley PA (eds) Tree-crop interactions: a physiological approach. CAB International, Wallingford, pp 189–233
Wallace JS, Gregory PJ (2002) Water resources and their use in food production systems. Aquat Sci 64:363–375
Wallace JS, Jackson NA, Ong CK (1999) Modelling soil evaporation in an agroforestry system in Kenya. Agric For Meteorol 94:189–202
Yamoah CF, Agboola AA, Mulongoy K (1986) Soil properties as affected by the use of leguminous shrubs for alley cropping in maize. Agric Ecosyst Environ 18:167–177
Acknowledgments
We thank the International Foundation for Science and USAID for funding and Posiano Nteziryayo for trial management and data collection.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Siriri, D., Wilson, J., Coe, R. et al. Trees improve water storage and reduce soil evaporation in agroforestry systems on bench terraces in SW Uganda. Agroforest Syst 87, 45–58 (2013). https://doi.org/10.1007/s10457-012-9520-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-012-9520-x