
Abstract
Fallowing can improve crop yields as a result of improved soil fertility and nutrient status. The objective of this work was to determine the effects of fallows and pruning regimes in coppicing fallows on soil moisture and maize yields under conventional tillage (CT) and no tillage (NT). Fallows that were evaluated were coppicing Acacia angustissima, non coppicing Sesbania sesban, natural fallow (NF) and continuous maize. In 2000/2001 season, maize yields were significantly different (P < 0.05) among treatments and were; 1.8, 1.2, 0.7 and 0.5 tonnes per hectare (t ha−1) under CT, while under NT yields were 1.3, 0.8, 0.7 and 0.2 t ha−1 for A. angustissima, maize, S. sesban and NF plots respectively. In 2001/2002 season, yields decreased in the order S. sesban > continuous maize > NF > A. angustissima, for both CT and NT. The 2-week pruning regime had significantly higher maize yields when compared to the 1 and 3 week pruning regime during the 2002/2003 cropping season. For the three seasons, CT had significantly higher yields than NT. A. angustissima had significantly higher mean available water at suctions <33 kPa for the 0–25 cm depth when compared to other fallow treatments. The bulk of the available water (47–80%) was retained at suction <33 kPa for all treatments and depths. There were no treatment differences in water retention at suctions >33 kPa for all treatments. It was concluded that improved fallowing increased yields when compared to NF. However, in coppicing fallows competition for water can result in reduced yields when there is rainfall deficiency, thus the need for pruning to manage the competition.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Most soils in the smallholder farming areas of Zimbabwe, tropical Africa and many other parts of the world have very low soil fertility, low crop yields and this has resulted in increased food insecurity. Most of these smallholder farmers are resource poor and they cannot afford high inputs of inorganic or organic nutrient supplements. Improved fallowing is one of many technologies which are currently being used for soil fertility improvement in low input agricultural systems in the semi arid to arid tropics (Kwesiga et al. 1999; Mafongoya et al. 2006). Improved fallows have been adopted by farmers Southern Africa in countries such as Zambia, Malawi (Kwesiga et al. 1999; Mafongoya et al. 2006), semi arid Kenya (Ong et al. 2006, 2007) and in humid tropics (Siriri et al. 2010). Improved fallows can increase nutrient input through nitrogen fixation (Sun et al. 2008; Mafongoya and Dzowela 1999; Chikowo et al. 2004; Mapfumo et al. 2005; Jose et al. 2004), they built up of soil organic matter (Nyamadzawo et al. 2008b; Lal 1989; Kwesiga and Beniest 1998) and they improve infiltration rates of soils (Young 1997; Nyamadzawo et al. 2008a).
Soil fertility benefits from fallowing are derived mainly from the use of annual, biannual or perennial nitrogen fixing trees or legumes which are either planted in rotation (e.g. improved fallows) or together with crops (e.g. alley or intercropping). Nitrogen from improved fallows can be made available to the subsequent crop when the crop residues decompose. Research has demonstrated that crop N uptake is the prime determinant of grain yield under low fertility conditions (Muchow and Sinclair 1995), thus improved legume tree fallows give significant increases in maize yields through enhanced N inputs (e.g. Mafongoya and Dzowela 1999; Mapfumo et al. 2005). Trees also act as “safety nets” and nutrient pumps as they help in closing the nutrient cycle by taking up nutrients leached into deeper layers to the surface (Jose et al. 2004). Legume trees have deeper rooting systems, and they break up hardened layers and help increase infiltration rates (Nyamadzawo et al. 2008a) and also help with the hydraulic lift which can bring water to the surface from deeper layers (Jose et al. 2004).
The disadvantage of using annuals as improved fallows is that they need to be re-planted regularly after each cropping phase, thus they are labour intensive and challenging to establish. The use of perennials legume fallows such as A. angustissima can be a solution as they do not need regular re-planting and they add huge amounts of biomass (Chikowo et al. 2004). One challenge of using coppicing fallows is that they can compete with crops for moisture and nutrients during the cropping phase, especially in low rainfall areas or during seasons when there is limited rainfall (Ong et al. 2006, 2007). Competition for water may cause moisture stress and this can result in reduced crop yields especially in drought years (Rao et al. 1992; Nyamadzawo 2004; Sun et al. 2008; Govindarajan et al. 1996). Competitive interactions involving water seem to be the most influential driving force of productivity (Jose et al. 2004). In another study, Jose et al. (2000a) reported that growing season water uptake in maize plants was reduced by up to 31.4% when tree roots were present and yields were reduced 35 and 33% for maize and black walnut and maize and red oak temperate cropping alley respectively. Similar observations were also made by Siriri et al. (2010) who reported reduce yields on a maize crop adjacent to unpruned calliandra, alnus and S. sesban relative to sole maize due to competition for water in the humid tropics, in Uganda. For a successful maize crop–tree fallow intercrop, there is need to manage coppices to limit their competitive impact (Schroth 1999), for example through pruning (Ong et al. 2007).
During the cropping phase of maize fallow rotation most smallholder farmers in Zimbabwe and southern Africa at large use conventional tillage (CT) to prepare the land. CT which involves using ox or donkey drawn moldboard plough is practiced because it improves the soil tilth and for weed control. CT has been associated with high rates of soil loss and physical degradation (Elwell 1989; Mrabet 2002; Nyamadzawo et al. 2009). Grandy and Robertson (2006) observed that years of soil regeneration can be lost after a single tillage event. Conservation tillage practices which minimize soil disturbance and promotes retention of crop residues on the soil surface are viable options for increasing water use efficiency because they reduce erosion (Giller et al. 2011) and increase water infiltration and storage in soil (Nyagumbo 2002; Mzezewa and van Rensburg 2011). Conservation tillage systems such as no tillage (NT) also maintains soil aggregation and higher SOC levels when compared with CT (Zotarelli et al. 2005; Grandy and Robertson 2006; Chivenge et al. 2007).
However, the two tillage practices have been found to affect yields differently. Lal (1976) and Moyo (1997) found that NT increased yields relative to CT. Some workers have found that NT had no effect on yields e.g. Carter et al. (1990), while Vogel et al. (1995) found that CT had a higher yield than NT. However, Hussein et al. (1999) found that CT resulted in higher yields during the early years, but yields decreased in successive years, relative to NT.
Thus, this study will evaluate the effects of different fallows and tillage on soil moisture and maize yields. This study will also evaluate the effects of different pruning regimes in coppicing fallows on soil moisture and maize yields. It was hypothesized that fallows improved soil moisture and maize yields and that frequent pruning in coppicing fallows reduces competition for water, hence increasing yields. The objective of this work was to determine the residual effects of coppicing and non coppicing fallows on maize yields in plots subjected to CT and NT and to evaluate the effects of pruning regimes in coppicing A. angustissima on soil water and maize yields.
Materials and methods
Study site
ICRAF Domboshawa Research site
The experimental site was at the Domboshawa Training Centre (DTC) which is about 30 km north of Harare, at approximately 19°35′S, 31°14′E and at 1,474 m altitude (Mafongoya and Dzowela 1999). The soils are classified as alfisols (USDA) or lixisols (FAO). The soil texture is sandy clay loam, with 22% clay and 71% sand. Table 1 summaries soil properties at the ICRAF site at DTC. Domboshawa receives unimodal rainfall; the mean annual rainfall is 750 mm, usually received from November to April. Figure 1 show the rainfall received at DTC during the period of the study.

Rainfall distribution at Domboshawa during the 2000/2001, 2001/2002 and 2002/2003 seasons
The study was conducted in a randomised block design experiment, replicated three times. The main treatments were fallows of Acacia angustissima (a coppicing fallow), Sesbania sesban, natural fallow (NF) and continuous maize (Zea mays) which served as the control. The experiment was initiated in the 1991–1992 season, to compare planted fallows of different duration. Fallows were established in a phased manner on 12 × 9 m plots, separated from each other by a distance of 2 m. Three-year fallows were first established in 1991–1992, 2-year in 1992–1993, and 1-year fallows in 1993–1994 season. This fallow phase was followed by a cropping phase during which all plots were cultivated by ox-drawn plough. After the end of 4 years of cropping, a new 2-year fallow phase was reinstated in November 1998 and the fallow period ended in October 2000 (Nyamadzawo et al. 2009). A continuous maize treatment was included as a control at the initiation of the experiment in 1992 and was maintained throughout the study period.
At the end of the fallow period in October 2000, the woody biomass was removed from the plots while leaf litter, fresh leaves and twigs were left in the plots. Total above ground litter additions after 2 years of fallowing were 10 and 5.7 Mg ha−1, for A. angustissima and S. sesban, respectively (Chikowo et al. 2004). Biomass from NF was burnt before ploughing from the control of weeds and pests and for easy of ploughing as is the normal practice in the smallholder farming sector. During the 2-year cropping phase (November 2000–April 2002) A. angustissima produced an additional average 1.5 Mg ha−1 annually through coppicing and this was cut and left in the plots.
In the third season, which started in December 2002–April 2003, only A. angustissima was used to evaluate the effects of pruning regimes as it was the only coppicing legume tree in the experiment. This experiment was initiated after realizing that the pruning regime that we used in the first two seasons (pruning once a month) contributed to moisture stress in A. angustissima plots and as a result the maize yields were depressed. In each block of the experiment there were three plots of A. angustissima and each was randomly picked and randomly assigned a pruning frequency, which were weekly, biweekly or 3 week intervals. Coppices from A. angustissima were cut, weighed and left in the plots after the removal of a subsample that was taken to the laboratory for the determination of moisture content.
Plot management
Plots were divided into two sub-treatments of CT and NT at fallow termination in 2000. Tillage involved ploughing using an ox-drawn mouldboard plough to a depth of 15–20 cm. In NT plots the soil was only disturbed during opening of planting holes using a hand hoe before planting maize. Maize was planted using hand hoes at an inter-row space of 90 cm and an in-row space of 30 cm at the beginning of each rain season (November/December 2000, 2001 and 2002). Plots were weeded using hoes twice during the growing season and this disturbed the top 0–5 cm depth.
Fertilizer was applied at the following rates; nitrogen (N) = 60 kg ha−1, phosphorus (P) = 15 kg ha−1 and potassium (K) = 10 kg ha−1, these were half the generally recommended fertilizer application rates. The maize varieties planted were hybrid Seed Co. (SC) 501 in the first season, hybrid SC 513 in the second and the third season.
Pruning frequencies measurements
From 2000 to 2003 maize was planted between the 1st and the 5th of December. In the first 2 years pruning was carried out once a month. However, in the third season (2002–2003) different pruning regimes were evaluated after noticing depressed crop yields in plots that had coppicing fallows. The first pruning was carried out 1 week after maize had germinated (1 week pruning regime), then after 2 and 3 weeks respectively for the 2 and 3 weeks pruning regimes. Coppicing fallows were pruned on the 16th, 23rd, 30th of December 2002; the 6th, 13th, 21st, 27th of January 2003; and the 10th, 17th and 25th of February 2003. A. angustissima coppices were cut from an area of 4 m2, then bagged and taken to the laboratory where it was oven dried and the dry weight recorded. A small representative sample was collected, dried and stored in khaki bags, while the rest was returned to the field.
Soil moisture analysis
Sampling for soil moisture coincided with pruning of coppices. Soil samples for moisture determination were collected from the 0–20, 20–40, 40–60, 60–80 and 80–100 cm depths using augers. Soil moisture content was determined gravimetrically in the laboratory soon after sampling. The volumetric water content was obtained after multiplying the gravimetric water content by the average bulk density. Bulk density was measured using samples that were collected using cores. Bulk density was calculated by dividing the mass of soil by the volume.
Soil water retention
Samples for water retention were collected at fallow termination, in October 2000 and October 2002. Samples were collected from the following depths: 0–5, 10–15, 20–25 and 35–40 cm using metal cores. Soil cores were saturated with water and equilibrated at the following suctions; 0, 5 and 10 using tension tables and 33, 200 and 1500 kPa using pressure plates. Soil moisture retention was calculated at different suctions and the results were used to plot soil water retention curves.
Maize yields
Grain and stover yields were measured from an 8.1 m2 net plot. The maize and residues were harvested at the end of the cropping season after the crop had reached physiological maturity and had dried. Grain was corrected for moisture content to 12.5% moisture level. Data was analysed by carrying out ANOVA using the split plot design for grain and stover yields, soil moisture content and water retention using Genstat statistical package (GENSTAT 2003). Least significance differences (LSD) were used to separate treatment means where there was significant difference.
Results
Maize yields in different fallows systems
During the 2000–2001 season there was above normal rainfall and there were no soil moisture deficit, while in 2001–2002 season there was a rainfall deficit in the last half of the season (Fig. 1). In the first season (2000/2001), fallow treatments showed significant differences (P < 0.05) in maize yields. Maize yields decreased in the following order; A. angustissima > continuous maize > S. sesban > NF (Fig. 2a). Among the fallow treatments, nitrogen fixing improved fallows of A. angustissima (coppicing fallow) and S. sesban (non coppicing fallow) out yielded NF. CT performed better than NT in A. angustissima, continuous maize and NF. For CT plots average maize yields were 1.8, 1.2, 0.7 and 0. 5 t ha−1 for A. angustissima, continuous maize, S. sesban and NF respectively, while under NT maize yields were 1.3, 0.8, 0.8 and 0.2 t ha−1 for A. angustissima, S. sesban, continuous maize and NF respectively (Fig. 2a). Under NT S. sesban had better yields that S. sesban under CT, however the yields were not statistically different. In the first year (2000/2001), stover yields were significantly higher in A. angustissima plots, which also gave the highest yields and lowest in NF plots, which gave the lowest maize yields. CT had higher biomass yields relative to NT and biomass yields decreased in the same order as grain yields.

a Maize yields in 2000–2001 seasons under CT and NT at half the recommended fertilizer rates. Vertical bar LSD, LSD at P < 0.05, = 217. NF natural fallow. b Grain yields in the 2001–2002 season under CT and NT practices at half the recommended fertilizer rates. Vertical bar LSD, LSD at P < 0.05, = 407. NF natural fallow
In the second cropping year (2001/2002), S. sesban had significantly higher grain yields for both CT and NT. Grain yield was highest in the CT plots relative to the NT plots (Fig. 2b). For CT maize yields were 1.6, 1.25, 1.2 and 0.5 t ha−1 for S. sesban, continuous maize, NF and A. angustissima respectively. For NT plots maize yields were 1.5, 0.6, 0.25 and 0.3 t ha−1 for S. sesban, continuous maize, NF and A. angustissima respectively (Fig. 2b). Stover yields were highest under CT when compared to NT. The order of decreasing stover yields for both CT and NT was S. sesban = NF > continuous maize > A. angustissima. Both grain and stover yields from A. angustissima were low because in the second season as there was a mid-season drought which caused moisture stress in the maize crop.
Maize yields under different pruning regimes in coppicing fallows
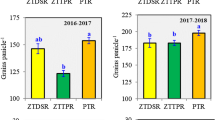
In the third year, 2002–2003, the average maize yields in A. angustissima were 2.0 t ha−1. The mean yields were 1.93, 2.14 and 1.95 t ha−1 for 1, 2 and 3 weeks pruning regime respectively. CT with fertilizer had significantly higher average yields (2.5 t ha−1) when compared to NT which had average yields of 2.0 t ha−1. CT with no fertiliser (2.1 t ha−1) had similar maize yields with NT with fertilizer (2 t ha−1), while NT with no fertiliser had the lowest maize yields (1.5 t ha−1).
Soil moisture distribution
Soil moisture distribution was related to the amount of rainfall received (Fig. 1). In the first season there was no soil moisture deficit, soil moisture was >10% during the season, except for December 2000 (Fig. 3). A summary of the soil moisture distribution of soil moisture during the first season is shown in Fig. 3. However, in the second season the average monthly soil moisture was low, the average soil moisture in the 0–20 cm depth was <10% and this was worse in the last half of the season. Soil moisture profiles for the second season are shown in Fig. 4. In the second season the mid season drought occurred at a period which coincided with 8 weeks after planting, during this period most of the maize roots were concentrated in the 0–60 cm depth (Chikowo et al. 2004). As a result maize roots were not able to extract limited amounts of water within the 0–60 cm depth, or the water beyond 60 cm depth and this resulted in water stress in the maize crop.

Soil moisture distribution for December 2000, January 2001, February 2001 and March 2001 during the 2000–2001 rain season. CT conventional tillage, NT no tillage, NF natural fallow

Soil moisture distribution for December 2001, January 2002, February 2002 and March 2002 during the 2001–2002 rain season. CT conventional tillage, NT no tillage, NF natural fallow
Soil water retention in improved fallows
In October 2000, there was a significant differences in water retention among treatments between 5 and 1,500 kPa, with A. angustissima having higher available water when compared to the other treatments and the same trend was shown in the 0–5, 10–15 and 20–25 cm depths (Fig. 5a–d). There were no significant differences among treatments in available water at the 35–40 cm depth and at suction >33 kPa. Most of the available water was held between 5 and 33 kPa. For the 5–33 kPa suctions, A. angustissima had significantly higher available water (Table 2).

Water retention curves for a 0–5 cm, b 10–15 cm, c 20–25 cm and d 35–40 cm depths at DTC for October 2000. The vertical bar is the LSD at P < 0.05
In October 2002, available water at all suctions had decreased when compared to 2000 for all treatments and there were significant differences in available water between the 2 years. A. angustissima had significantly higher water retention in the 0–5 cm depth and NF had higher water retention for the 10–15 and 20–25 cm depths (Fig. 6a–c). For both October 2000 and 2002 there were significant differences in water retention at suctions <33 kPa, but no treatment differences in water retention at suctions >33 kPa for all treatments (Figs. 5, 6). Available water retained at suction <33 kPa was 77–80, 75–79, 63–68 and 47–66% of the total available water or the 0–5, 10–15, 20–25 and 35–40 cm depths respectively (Table 2).

Water retention curves for a 0–5 cm, b 10–15 cm, c 20–25 cm and d 35–40 cm depths at DTC for October 2002. The vertical bar is the LSD at P < 0.05
For both October 2000 and 2002, available water decreased as soil depth increased. There were no treatment differences in water retention at soil depths >25 cm and at suction >33 kPa (Figs. 5, 6). Soil water retention decreased with depth, possibly because soil organic matter decreased as depth increased. There were no significant changes in clay content with depth. There were no significant tillage effects on water retention for both October 2000 and 2002.
Effects of pruning regimes on biomass and soil moisture distribution
In the third season, pruning had an effect on biomass productivity and soil moisture levels. The mean seasonal biomass yields were 3.7, 5.3 and 6.4 Mg ha−1 for the 1, 2 and 3 weeks pruning regime respectively. Total biomass productivity was significantly higher (P < 0.05) under 3 weeks pruning regime than in 1 and 2 weeks pruning regimes. There were no significant tillage effects on biomass productivity for the 1 week pruning regime, however for the 2 and 3 week pruning regimes there were significant differences between NT and CT, with NT have higher biomass productivity when compared to CT.
Soil moisture was significantly different (P < 0.05) between pruning regimes. For CT the average volumetric soil moisture levels in the 1 week cutting frequency were significantly higher (18.5%) when compared to the 2 and 3-week pruning regimes which had volumetric soil moisture of 15.1 and 14.6% respectively. For NT there were no significant differences in the average soil moisture among the one and 2 week pruning regimes (17.5%), but these two pruning regimes were different from the 3 week pruning regime which had a volumetric moisture content of 15.5%. There were only significant treatment differences for the 2 week pruning regime.
Discussion
Grain and biomass yield
The grain and biomass yields were highest in A. angustissima in the first year of cropping. The same A. angustissima plots had the highest biomass residues from the fallowing years. In addition A. angustissima generated coppices and this resulted in continuous biomass addition during the cropping. Enhanced nitrogen (N) levels from the accumulated biomass could have resulted in increased crop yields (Muchow and Sinclair 1995; Mafongoya and Dzowela 1999). In addition to the high biomass and N addition, there was adequate rainfall during the first post fallow cropping season (2000/2001).
Unlike in the temperate regions were agroforestry systems are managed with high input of inorganic or organic nutrient supplements (Jose et al. 2000b), in the tropical Africa most smallholder farmers cannot afford organic and inorganic nutrient supplement, thus they rely solely on the enhanced fertility from N fixing legume trees. Grain yields are usually low and they respond inorganic N applications. Studies in from the tropics (Chikowo et al. 2004) and the temperate regions (Jose et al. 2000b) showed that although N and P are added to the subsequent crops from the decomposing tree litter (leaves and roots), a considerable amount of fertilizer input is still needed to maintain productivity.
During the first cropping season the maize crop in S. sesban plots did not perform well because it was infested by cut-worms, this reduced biomass and grain yields. Both grain and biomass yields were lowest in NF, because it was not nitrogen fixing though it is good at improving soil physical properties such as infiltration rates (Nyamadzawo et al. 2007). In addition, low soil N and increased immobilization in NF may have resulted in reduced available N for plant uptake, poor crop establishment and low yields.
In the second year, which had a mid season drought, grain and biomass yields were highest in S. sesban. This was probably because there was reduction in nutrient uptake during the first year as the crop failed due to cut worm infestation and therefore residual fertility was higher. Secondly most of the plant’s litter and leaves from S. sesban had decomposed and there were available nutrients for plant uptake. Studies by Mafongoya and Dzowela (1999) showed that S. sesban residues were of better quality when compared to A. angustissima. The crop was able to use these nutrients to establish a good crop stand (Chikowo et al. 2004). Maize grown in plots where there was S. sesban, a non coppicing fallow, did not compete for water with coppices as was the case in A. angustissima plots. In continuous maize plots low yields were attributed to continuous monoculture cropping which resulted in reduced soil nutrients, reduced infiltration rates and soil structural degradation (Nyamadzawo et al. 2007) and also low plant available water (Table 2).
In coppicing A. angustissima, maize yields were very low in the second season of the study (2001/2002). A mid-season drought resulted in serious competition for water between the coppicing fallow and the maize crop. Work by Chikowo et al. (2004) at the same site showed that most maize roots were concentrated in the 0–60 cm depth, 8 weeks after planting, a period which coincided with the mid season drought. Soil moisture content was lower in the 0–60 cm depth and this resulted in severe moisture stress in the crop. The study by Chikowo et al. (2004) also showed that maize has a low root length density, which is concentrated mainly in the 0–20 cm depth; as a result water uptake at depths greater than 20 cm was low. Similar observation were also made by several other researchers in the temperate zone (Lehmannn et al. 1998; Jose et al. 2000a; Wanvestraut et al. 2004) who reported observing the greatest concentration of tree-root density within the top 30 cm of soil, the region which was predominantly explored by crop root systems and this zone had a more prominent variation in soil moisture. Wanvestraut et al. (2004) reported that the upper 0–30 cm soil layer alone contained 52% of the total fine root biomass.
The use of coppicing fallows can result in serious competition for moisture when there is rainfall deficiency or in semi arid areas and the result is depressed crop yields (Rao et al. 1992). In this study although coppices from A. angustissima fallows were cut once a month to reduce competition between coppices and crop during the first two seasons, this did not reduce moisture stress in the maize crop during the second season. Similar results have been reported from a maize–leucaena intercrop (Leucaena leucocephala (Lam.) de Wit.) in Kenya (Govindarajan et al. 1996). Miller and Pallardy (2001) also reported that competition for water to reduce crop yields by 22–27% in silver maple and maize, while Jose et al. (2000b) reported a 35 and 33% reduction in maize yields for black walnut and red oak systems, respectively in temperate region of the United States. Frequent pruning was found to reduce apical dominance of the root system, so that they become branched and superficial (Hairiah et al. 1992), thus increasing competition for water, as increased numbers of roots scavenge for water in the top soil layer.
Water retention was highest in fallows relative to continuous maize at suctions <33 kPa. At lower suction water retention is structure controlled, while at higher suctions it is texture controlled (Hillel 1982). Therefore the differences in water retention at lower suction are a reflection of structural differences between fallows and continuous maize (Table 2). At lower suctions water retention is also affected by the amount of SOC. The differences in SOC levels may have resulted in greater water holding capacity in fallows than in continuous maize (Young 1997). In the top 0–15 cm depth, between 75 and 80% of the total available water was retained at suction <33 kPa. This showed the important role of SOC in water retention. Water retention was highest in A. angustissima, which also had the highest amounts of SOC. The SOC levels were highest in A. angustissima and lowest in continuous maize (Nyamadzawo et al. 2008b). In A. angustissima a 3.8 g kg−1 increase in SOC over the NF resulted in a difference of 23% in water holding capacity. The decrease in available water with increasing depth reflected a decrease in SOC with depth (Nyamadzawo et al. 2008b). There were no differences in texture in the different plots and this explains why there were no significant differences in water retention at higher suctions. Water retention was not significantly different between CT and NT and this agrees with Lindwall et al. (1984), who found no significant differences in water retention between NT and CT after carrying out a 5-year study on water conservation in two tillage systems in a maize cropping system. Although A. angustissima had the highest plant available water when compared to the other fallow systems that were evaluated (Table 2), increased transpiration from coppices of A. angustissima used up most of the available water and very little was left for the maize crop, resulting in low maize yields.
Effects of tillage on grain and stover yield
CT had higher yields relative to NT during the two seasons. This may be due to increased mineralisation in CT relative to NT which allowed maize under CT plots to establish a good stand quickly. Results from N mineralisation studies at the same site have shown faster mineralisation in CT than in NT (Chikowo et al. 2004). In a 5-year study of water conservation by Lindwall et al. (1984), water conservation was greater in CT than in NT. Other studies have shown that the existence of better pore continuity to lower soil depth under NT can result in greater water loss by evaporation (Phillips 1981). Schillinger and Bolton (1993) have also shown that residues in NT reduced evaporation during frequent rainfall periods, but evaporation in NT was higher during long dry spells due to none disturbance of capillary flow resulting in reduced crop yields. Giller et al. (2011) suggested that as a result of lower maize yields under conservation agriculture many smallholder farmers have been reluctant to adopt conservation agriculture.
The use of fertilizer in crop production systems is an economic investment. In a study in Kentucky, Phillips (1981) reported that low N rates of fertilizer produced lower yields under NT than under CT. However, the same study reported that when rates were increased NT yielded more than CT. Campbell et al. (1998) found that in wheat NT yielded more at N rates greater than 75 kg ha−1, however at rates lower than 75 kg ha−1 CT yielded more than NT. High yields in CT at low N levels could be due to enhanced N mineralisation after tillage. Chikowo et al. (2004) reported faster N mineralisation and had better yields under CT at the same study site. At Domboshawa, N was applied at a rate of 60 kg ha−1 and this could partly explain why NT had lower yields than CT. House et al. (1984) found out that under NT yields were low in early years and this was probably due to reduction in soil organic matter and N mineralisation and increased immobilization of fertilizers in NT than in CT. Although fallows under NT had better soil structure (Nyamadzawo et al. 2007) these benefits did not translate to improved maize yields. This showed that for the soils at the ICRAF Domboshawa Research site, soil fertility was a more limiting factor than soil structure.
Effects of different pruning regimes on maize yields
One disadvantage of using coppicing fallows is that they compete with crops for moisture and nutrients during the cropping phase, especially in low rainfall areas or during seasons when there is limited rainfall (Ong et al. 2006, 2007). Competition for water may cause moisture and nutrient stress and this can result in reduced crop growth and yields especially in drought years (Rao et al. 1992; Nyamadzawo 2004; Sun et al. 2008). Similar results of reduced crop yields were also reported in a Leucaena-millet (Pennisetum glaucum (L.)) inter-cropping (Ong et al. 1991), maize–leucaena intercrop (Govindarajan et al. 1996), while Siriri et al. (2010) reported a reduction in maize yields on maize crop adjacent to unpruned calliandra, alnus and sesbania relative to sole maize. Legume trees such as A. angustissima have an extensive root system that can out-compete annual crops such as maize which have roots mainly in 0–100 cm depth (Chikowo et al. 2004).
Having a higher fine root biomass may favor trees when they are mixed with crops as in an alley cropping system (Jose et al. 2000b). For a successful maize crop–coppicing fallow intercrop, there is need to manage to limit their competitive impact (Schroth 1999), for example through pruning (Ong 2007).
Maize yields were significantly higher under the 2 week pruning frequency when compared to the 1 and 3 week pruning frequencies during the 2002–2003 season. Increased the interval between pruning resulted in increased competition for water and possibly nutrients between the fallow trees and the crop and this was shown by lower yields in the 3 week pruning frequency. Siriri et al. (2010) reported that crop yields may decreased probably due to increased competition and declining soil fertility. Increased interval between prunings may also reduce the quality of biomass pruned, although quality and mineralisation studies were not carried out.
Although there was less biomass accumulation under the 1 week pruning regime when compared to the 2 and 3 weeks pruning regimes, the 1 week pruning regimes had greater soil moisture content. Jackson et al. (2000) reported that severe shoot pruning can reduce water use by trees, improving recharge of the crop rooting zone in semi-arid areas. Frequent pruning can also result in low shoot re-growth and increased apical dominance of roots. Jones et al. (1998) found that shoot pruning of Prosopis juliflora in semi-arid Nigeria reduced competition for below-ground and above-ground resources, but also reduced root length density.
However, Hairiah et al. (1992) found that frequent cutting reduced apical dominance of the root system, such that they become branched and superficial resulting in increased competition for water, as the increased numbers of roots scavenge for water in the soil in the tropics. Jose et al. (2000b) also reported that when tree roots were pruned in either ‘trench’ or ‘barrier’ treatments, a higher root biomass was observed and this implied increased root proliferation along the disturbed soil profile. In this study no clear explanation could be given for high yields under the 2 week pruning regime when compared to the 1 or 3 week regimes during the 2002–2003 season. Thus, there is need for more studies to evaluate the effect of different pruning regimes on crop yields.
Conclusions
Improved fallowing with A. angustissima and S. sesban increased maize yields when compared to NF, non nitrogen fixing fallow and continuous maize. Coppicing fallows enhanced maize yields only when there is no competition for moisture. Even though A. angustissima had the greatest plant available water, severe competition for water with the maize crop during periods of moisture deficiencies resulted in reduced maize yields, thus the need to frequently prune to reduce competition with crops. In non coppicing improved fallows such as S. sesban, maize yields were higher even during periods of moisture stress and are recommended for low rainfall areas or in areas when there are frequent rainfall deficiencies. NFs had low yields throughout the study period and this because NF provided little soil fertility improvement.
References
Campbell CA, Thomas AG, Brederbeck VO, MacConkey BC, Selles F, Spurr D, Zertner RP (1998) Converting from NT to pre-seeding tillage: influence on weeds, spring wheat grain yields and nitrogen, soil quality. Soil Tillage Res 46:175–185
Carter MR, White RP, Andrew RG (1990) Reduction of secondary tillage in mouldboard ploughed system for silage corn and spring cereals in medium textured soils. Can J Soil Sci 69:150–156
Chikowo R, Mapfumo P, Nyamugafata P, Giller KE (2004) Woody legume fallow productivity, biological N2 fixation and residual benefits to two successive maize crops in Zimbabwe. Plant Soils 262:303–315
Chivenge PP, Murwira HK, Giller KE, Mapfumo P, Six J (2007) Long term impact of reduced tillage and residue management on soil carbon stabilization: implications for conservation agriculture on contrasting soils. Soil Tillage Res 94:328–337
Elwell HA (1989) Soil structure under conservation tillage. In: Conservation tillage. Commercial Grain Producers Association handbook, Harare
GENSTAT (2003) GESTAT statistical package, 8th edn. VSN International, Oxford
Giller KE, Corbeels M, Nyamangara J, Triomphe B, Affholder F, Scopel E, Tittonell P (2011) A research agenda to explore the role of conservation agriculture in African smallholder farming systems. Field Crops Res. doi:10.1016/J.Fcr.2011.04.010
Govindarajan M, Rao MR, Mathuva MN, Nair PKR (1996) Soil–water and root dynamics under hedgerow intercropping in semiarid Kenya. Agron J 88:513–520
Grandy AS, Robertson GP (2006) Aggregation and organic matter protection following tillage of a previously uncultivated. Soil Sci Soc Am J 70:1398–1406
Hairiah K, van Noordiwick M, Santoso B, Syekhfani MS (1992) Biomass production and root distribution of eight trees and their potential for hedgerow intercropping on an ultisol in Southern Sumatra. Agrivita 15:54–68
Hillel D (1982) Introduction to soil physics, 2nd edn. Academic Press, Orlando
House GF, Stinner BR, Crosselt I, Odim EP, Langdale GW (1984) Nitrogen cycling in conventionally tilled and no till agroecosystems. J Soil Water Conserv 39:194–200
Hussein I, Olson KR, Elbar SA (1999) Impacts of tillage and no tillage on production of maize and soyabean on an eroded Illinois silt loam soil. Soil Tillage Res 52:37–49
Jackson NA, Wallace JS, Ong CK (2000) Tree pruning as a means of controlling water use in an agroforestry system in Kenya. For Ecol Manag 126:133–148
Jones M, Sinclair FL, Grime VL (1998) Effect of tree species and crown pruning on root length and soil water content in semi-arid agroforestry. Plant Soil 201:197–207
Jose S, Gillespie AR, Seifert JR, Mengel DB, Pope PE (2000a) Defining competition vectors in a temperate alley cropping system in the mid-western USA. 3. Competition for water. Agrofor Syst 48:41–59
Jose S, Gillespie AR, Seifert JR, Mengel DB, Pope PE (2000b) Defining competition vectors in a temperate alley cropping system in the mid-western USA. 3. Competition for nitrogen and litter decomposition dynamics. Agrofor Syst 48:61–77
Jose S, Gillespie AR, Pallardy SG (2004) Interspecific interactions in temperate agroforestry. Agrofor Syst 61:237–255
Kwesiga F, Beniest J (1998) Sesbania sesban for Eastern Zambia—an extension guideline. ICRAF, Nairobi
Kwesiga FR, Franzel S, Place F, Phiri D, Simwanza CP (1999) Sesbania sesban improved fallow in eastern Zambia: their conception, development and farmer enthusiasm. Agrofor Syst 47:49–66
Lal R (1976) No-tillage effects on soil properties under different crops in western Nigeria. Soil Sci Soc Am J 40:762–768
Lal R (1989) Agroforestry systems and soil management of a tropical alfisol. IV. Effects of soil physical and soil mechanical properties. Agrofor Syst 8:197–215
Lehmann J, Peter I, Steglich C, Gebauer G, Huwe B, Zech W (1998) Belowground interactions in dryland agroforestry. For Ecol Manag 111:157–169
Lindwall CW, Sawatzky B, Jensen T (1984) Zero tillage in Southern Alberta. In: Proceedings of Alta Soil Science Workshop, Faculty of Extension, University of Alberta, Edmonton, Alta, pp 128–136
Mafongoya PL, Dzowela BH (1999) Biomass production of tree fallows and their residual effect on maize in Zimbabwe. Agrofor Syst 47:139–151
Mafongoya PL, Kuntashula E, Sileshi G (2006) Managing soil fertility and nutrient cycles through fertilizer trees in southern Africa. In: Uphoff N, Ball AS, Fernandes E, Herren H, Husson O, Liang M, Palm C, Pretty J, Sanchez P, Sanginga N, Thies J (eds) Biological approaches to sustainable soil systems. Taylor & Francis, New York, pp 274–289
Mapfumo P, Mutambanengwe F, Giller KE, Mpepereki S (2005) Tapping indigenous herbaceous legumes for soil fertility management by resource-poor farmers in Zimbabwe. Agric Ecosyst Environ 109:221–233
Miller AW, Pallardy SG (2001) Resource competition across the crop–tree interface in a maize–silver maple temperate alley cropping stand in Missouri. Agrofor Syst 53:247–259
Moyo A (1997) Annual report on soil loss erosion assessment of different tillage systems; results of Makoholi on station trials 1993/94. Conservation Tillage Project, Institute of Agricultural Engineering, Harare
Mrabet R (2002) Stratification of soil aggregation and organic matter under conservation tillage systems in Africa. Soil Tillage Res 66(2):119–128
Muchow RC, Sinclair TR (1995) Effect of nitrogen supply on maize yield: II. Field and model analysis. Agron J 87:642–648
Mzezewa J, Van Rensburg LD (2011) Effects of tillage on runoff from bare clayey soil on a semi-arid ecotope in the Limpopo Province of South Africa. Water SA 37(2):165–172
Nyagumbo I (2002) Effects of three tillage systems on seasonal water budget and drainage of two Zimbabwean soils under maize. DPhil thesis, Department of Soils Science and Agricultural Engineering, University of Zimbabwe
Nyamadzawo G (2004) The effects of improved fallows and tillage management on soil and water conservation in a sandy soil. MPhil thesis, Department of Soil Science and Agricultural Engineering, Faculty of Agriculture, University of Zimbabwe
Nyamadzawo G, Nyamugafata P, Chikowo R, Giller KE (2007) Improved legume tree fallows and tillage effects on structural stability and infiltration rates of a kaolinitic sandy soil from central Zimbabwe. Soil Tillage Res 96(1–2):182–194
Nyamadzawo G, Nyamugafata P, Chikowo P, Giller KE (2008a) Residual effects of fallows on infiltration rates and hydraulic conductivities in a kaolinitic soil subjected to conventional tillage (CT) and no tillage (NT). Agrofor Syst 72:161–168
Nyamadzawo G, Chikowo R, Nyamugafata P, Nyamangara J, Giller KE (2008b) Soil organic carbon dynamics of improved fallow–maize rotation systems under conventional and no tillage in central Zimbabwe. Nutr Cycl Agroecosyst 81:85–93
Nyamadzawo G, Nyamangara J, Nyamugafata P, Mzulu A (2009) Soil microbial biomass and mineralization of aggregate protected carbon in fallow–maize systems under conventional and no-tillage in central Zimbabwe. Soil Tillage Res 102:151–157
Ong CK, Corlett JE, Singh RP, Black CR (1991) Above and belowground interactions in agroforestry systems. For Ecol Manage 45:45–57
Ong CK, Black CR, Muthuri CW (2006) Modifying forestry and agroforestry to increase water productivity in the semi-arid tropics. CAB reviews: perspectives in agriculture, veterinary science. Nutr Nat Resour 1(65):1–19
Ong CK, Anyango S, Muthuri CW, Black CR (2007) Water use and water productivity of agroforestry systems in the semi-arid tropics. Ann Arid Zone 46:255–284
Phillips RE (1981) Soil moisture. In: No tillage research. Research reports and reviews. University of Kentucky, Lexington, pp 23–42
Rao MR, Ong CK, Pathak P, Sharma MM (1992) Productivity of annual cropping and agroforestry systems on shallow alfisol in semi arid India. Agrofor Syst 15:51–64
Schillinger WF, Bolton FE (1993) Fallow water storage in tilled vs untilled soils in Pacific North West. J Prod Agric 6:267–269
Schroth G (1999) A review of below-ground interactions in agroforestry, focusing on mechanisms and management options. Agrofor Syst 43:5–34
Siriri D, Ong CK, Wilson J, Boffa JM, Black CR (2010) Tree species and pruning regime affect crop yield on bench terraces in SW Uganda. Agrofor Syst 78:65–77
Sun H, Tang Y, Xie J (2008) Contour hedgerow intercropping in the mountains of China: a review. Agrofor Syst 73:65–76
Vogel H, Nyagumbo I, Olsen K (1995) Effects of tied ridging and mulch ripping on water conservation in maize production on sand veld. Conservation Tillage Project, Institute of Agricultural Engineering, Harare
Wanvestraut R, Jose S, Nair PKR, Brecke BJ (2004) Competition for water in a pecan-cotton alley cropping system in the southern United States. Agrofor Syst 60:167–179
Young A (1997) Agroforestry for soil management. C.A.B. International in association with ICRAF, UK
Zotarelli L, Alves BJR, Urquiaga S, Torres E, Dos Santos HP, Paustain K, Boddey RM, Six J (2005) Impact of tillage and crop rotation on aggregate-associated carbon in two oxisols. Soil Sci Soc Am J 69:482–491
Acknowledgments
We thank ICRAF for the facilities at Domboshawa experimental site. We are grateful for funding given to the Department of Soil Science and Agricultural Engineering, University of Zimbabwe by the European Union (IMPALA Project No. ICA4-CT2000-30011) and ICRAF’s ANAFE Scholarship fund.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Nyamadzawo, G., Nyamugafata, P., Wuta, M. et al. Maize yields under coppicing and non coppicing fallows in a fallow–maize rotation system in central Zimbabwe. Agroforest Syst 84, 273–286 (2012). https://doi.org/10.1007/s10457-011-9453-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-011-9453-9