
Abstract
In Western Africa, interactions between trees and agricultural crops are a key element in determining parkland management in an agricultural environment that is rapidly changing. Eggplant (Solanum melongena), chilli pepper (Capsicum annuum), taro (Colocasia esculenta) and pearl millet (Pennisetum glaucum) were tested for their shade tolerance under Parkia biglobosa trees in south-central Burkina Faso using a split-plot design. Soil characteristics, chlorophyll fluorescence and crop growth and yield were measured to quantify the effect of P. biglobosa on the crops and their environment. The experiment ran during 2 years. P. biglobosa suppressed the vegetative growth and yield of pearl millet in both years. Eggplant and chilli pepper were severely injured by the rains and produced fruits only during the first year. Eggplant yields were suppressed by trees to between one third and one tenth of the yield in the control plots. However, chilli pepper yields increased by up to 150% when grown under the tree canopy compared to the control. In both years, the vegetative growth and yield of taro was higher when grown in the shade than outside the tree canopy.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Production of annual crops in sub-Saharan Africa is often practiced in parkland agroforestry systems where scattered trees create a permanent open upper storey (Nair 1993). These trees are rarely planted and were often left standing when the original woodlands were cleared (Gijsbers et al. 1994). Parkia biglobosa (Jacq.) Benth is one of the dominant multipurpose species of the West African parklands (Boffa 1999). This species grows best where annual rainfall is between 600 and 1200 mm (Boffa 1999), is found in savannah zones, and has a natural distribution ranging from 5°N to 15°N and 16°W to 32°E (Hopkins and White 1984). P. biglobosa, known locally as néré and in English as the African locust bean, is a member of the leguminosæ but shows no evidence of nodulation and nitrogen fixation (Tomlinson et al. 1995). It is a deciduous tree which grows to heights of 7–20 m and has a wide crown extending outwards from the bole by up to 10 m (FAO 1988). Its extensive root system may extend up to 60 m from the trunk (Jonsson 1995) and the lateral roots may cover an area eight times greater than that of the crown (Tomlinson et al. 1995). One of its most important products is its 15–25 cm long pods (FAO 1988). The seeds have a hard testa and can be fermented to produce “soumbala”, a black and strong smelling proteinaceous food consumed as a spicy seasoning for local dishes (Boffa 1999; Teklehainmanot 2004). The making and selling of “soumbala” is one of the few activities widely practiced by women to provide cash income (Gausset et al. 2005).
Several authors have reported a decrease in tree densities during the past few decades in some agricultural areas of the Sahel region and low regeneration in parklands (Lericollais 1989; Gijsbers et al. 1994). Interactions between trees and associated crops appear likely to influence farmers’ management of tree densities in parklands, although a range of socio-economic, biological and policy factors may also influence management (Boffa 1999; Boffa et al. 2000). The major crops associated with P. biglobosa are sorghum (Sorghum bicolor) and pearl millet (Pennisetum glaucum) (Boffa et al. 2000). Most studies on the effect of néré on cereal crops have shown that production is reduced when crops are grown under trees compared to open fields (Kessler 1992; Kater et al. 1992; Wilson et al. 1998). This depressive influence of trees was attributed to reduced light intensity and increased atmospheric humidity and soil water content. While most previous studies have concentrated on common cereal crops, the available information on the interaction between parkland species and other annual crops is scarce. However, many other species, for example large leafed vegetables and root crops, have been suggested as being suitable alternative under storey crops (Kater et al. 1992; Teklehainmanot 2004) due to their relative shade tolerance and high soil fertility and moisture requirements (Kessler 1992; Kater et al. 1992). The objective of this paper is to assess the effects of P. biglobosa on the growth and yield of P. glaucum and three food crop species, Solanum melongena (eggplant), Capsicum annuum (chilli pepper) and the tuber crop Colocasia esculenta (taro). We hypothesized that the three latter crops would have shade-tolerant properties and perform better under P. biglobosa than P. glaucum.
Materials and methods
Study site
The study was carried out over a two year period (2006–2007) in the parklands of Nobéré (11°30′N and 1°50′W), a town of ca. 2500 inhabitants located 113 km south of Ouagadougou, Burkina Faso, at an altitude of 300 m.a.s.l. The parklands of Nobéré are typical of sub-Saharan Africa as they are dominated by P. biglobosa and Vitellaria paradoxa. The rainfall is unimodal with an average annual total of 500–900 mm. Annual rainfall for 2006 and 2007 was, respectively, 843 and 946 mm, and was greatest between June and September (Fig. 1).

Rainfall during 2006 and 2007 at Nobéré in Burkina Faso, West Africa
Trees
Six mature P. biglobosa trees were selected for the experiment in 2006 (Table 1), but only five of these could be used in 2007, due to lack of agreement with the tree owner. Selection was based on their position relative to other trees to ensure that no nearby trees could shade them. The trees were growing in a 1 km2 zone situated 6.4 km West of Nobéré. The fields under the trees had been left fallow for 2 years, before which they had been under maize, sorghum, millet or cotton cultivation. The site was flat, and the soil had no sharp texture change across the site.
Crops
Four food crop species were included in the experiment based on interviews intended to identify plants considered as shade tolerant by ten farmers in Nobéré. The following crops were particularly mentioned as shade tolerant; the number of respondents mentioning specific crops is shown in parenthesis: Solanum melongena (eggplant) (5), Capsicum annuum (chilli pepper) (7) and Colocasia esculenta (taro) (7). These crops were selected for study. Pennisetum glaucum (pearl millet) was also selected as a control for comparison with the other crops.
Eggplant and chilli pepper were sown in May (2006 and 2007) in a plant nursery in Nobéré, and transplanted to the field at the beginning of July 2006 and mid-July 2007. Pearl millet seeds and taro tubers were sown directly in the field in July in both years.
Table 2 gives details of the cultivation practices used. All treatments to which crops were subjected corresponded to local cultivation practices. Cultivation practices were less intensive during 2007 than 2006.
Taro seedlings emerged ca. 14 days after sowing, while pearl millet seedlings emerged ca. 7 days after sowing. Pesticide (Deltamethrin at approximately 25 g a.i/ha) was applied to eggplants, at 2 week intervals between the end of July and mid-September. Weeding was done by hand for all crops.
Experimental design
Measurements of crop performance and nutrient status were made under the crown of all selected trees. The area under each tree was subdivided into three concentric zones:
- Zone A:
-
from the trunk to half of the crown projection radius;
- Zone B:
-
from half of the crown projection radius to the edge of the crown projection;
- Zone C:
-
from the edge of the crown projection to a distance of 3 m
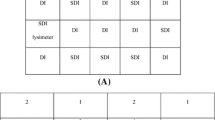
Unshaded control plots (8 × 8 m) were planted at least 40 m from the edge of the crown of all sampled trees and any other trees. The area under the tree crowns and within the control plots was further divided into quarters to allow crops to be grown in a split-plot design (Fig. 2).

Schematic representation of the experimental design around the tree trunk at Nobéré in Burkina Faso, West Africa. A square plot (8 × 8 m) remote from the trees was included as an unshaded control
Crop growth and yield
The following measurements were made in each concentric zone for all crops: weekly counts of (1) leaf number per plant and (2) plant height from the ground to the highest leaf for four randomly selected plants for each concentric zone and crop species; and (3) total weight (fresh) of harvested fruit or pearl millet panicles for all plants in 2.25 m2 quadrats in each concentric zone. Yield data were not collected for chilli pepper and eggplant in 2007 because fruit production was negligible. The lower yields in 2007 could be attributed to the high rainfall amounts received in August which led to a slower growth for pearl millet and taro, as well as flower shedding for chilli pepper and eggplant, along with fruit rot for eggplant.
Soil properties
Soil samples were taken at depths of 0–10, 10–20, 20–30, 30–40 and 40–50 cm in October 2006 under each tree canopy for each concentric zone. Due to budget limitations, samples were pooled for specific concentric zones, giving a total of 20 samples for analysis, i.e. one for each soil depth in each concentric zone and control plots. The samples were air-dried prior to chemical analyses at the Bureau National des Sols (BUNASOLS) laboratories in Ouagadougou. Soil texture was determined using the Bouyoucos method (Day 1965). Total soil N was measured by the Kjedahl method and total P was determined by acid mineralization (Houba et al. 1985). Total C and organic matter levels were analysed using the Walkley–Black method (Allison 1965) and Ca and Mg levels were determined by silver thiourea extractions (Pleysier and Juo 1980). pH (water and KCl) was measured with a pH-meter equipped with a glass electrode (WTW InoLab, Weilheim, Germany) in slurries formed from a ratio of 1 g soil to 2.5 ml water (Peech, 1965).
Chlorophyll fluorescence
Measurements of F0, the initial fluorescence of dark adapted leaves, and FM, the maximum fluorescence of light adapted leaves were made using a Mini-PAM with leaf clip holders (Heinz Walz GmBH, Effeltrich, Germany). In 2006, three plants for each concentric zone and control plot were randomly selected for three randomly chosen trees, giving a total of 36 samples for each crop. Measurements were made on three occasions during the growing season (31 August, 6 October and 14 October 2006) between 0900 and 1100 h local time. In 2007, two plants per concentric zone and control plot were randomly selected for two randomly chosen trees, giving a total of 18 plant samples for each crop. Measurements were made at 0900, 1000, 1100 and 1700 h local time on 21 September 2007. All measurements were made following dark adaptation for 45 min and the maximum quantum yield of PSII photochemistry ΦP0 was calculated as (Roháček 2002):
where FV is the variable fluorescence (FV = FM − F0).
Statistical analysis
Data were analyzed using SAS version 9.1 software. Differences between treatments for all variables examined were tested using ANOVA General Linear Model (GLM) regression with Tukey’s corrections for comparison of means. Data for the different crops and years were analyzed separately and all analyses were performed using plot mean values. The linear models were:
where Y ijk represents the mean value for plant height, leaf number, or yield for each crop, μ is the general mean, a i is the fixed effect of the cardinal direction in which the crop was grown, β j is the fixed effect of the “zone” in which the crop was grown, d(k) is the random effect of the tree, given that d(k) and ε ijks are normal with mean 0 and variance σ2 and σ 2tree and all d(k) and ε ijks values are independent. In all cases, the fixed effect of cardinal direction was found to be statistically insignificant.
The following model was used for the chlorophyll fluorescence measurements:
where Y ijk is the mean value for chlorophyll fluorescence, μ is the general mean, φ i is the fixed effect of crop, β j is the fixed effect of the “zone” in which the crop was grown, θ ij is the fixed effect of the interaction between crop and zone, dk is the random effect of tree, given that d(k) and ε ijks are normal with mean 0 and variance σ2 and σ 2tree and all d(k) and ε ijks values are independent.
Model validation was carried out using residual plots and statistical significance was set at P < 0.05.
Results
Growth and yield varied considerably between the two seasons. Whereas 2006 could be characterized as a normal year, 2007 experienced heavy rainfall in August which caused flooding in some parts of the village. This was reflected by crop yields, which were much higher in 2006 than in 2007.
Crop height and leaf number
The different growth conditions in the 2 years were also reflected by plant heights for pearl millet and eggplant and leaf numbers per plant for all crops measured ca. 110 days after sowing (DAS), as the values for both variables were much smaller in 2007 than in 2006 (Table 3). The differences in height and leaf number between pearl millet growing close to and further away from tree boles were statistically significant. In 2006, mean plant height in zone A was 50 cm less than in control plots (P < 0.0001), while in 2007, the tallest plants were in zone C and the control plot, being 46 cm taller than in zone A (P = 0.0002). Differences in leaf number between zones were not significant (Table 3). The opposite trend was found for taro as mean plant height was 14 cm (2006) and 16 cm (2007) greater in zone A than in the control plots (P < 0.0001). Leaf number for taro plants growing closest to the trunk was 15% greater than outside the tree canopy in 2006 (P < 0.0001).
At ca. 110 days after transplanting, mean plant height for chilli pepper plants in zone A was 8 cm (2006) and 14 cm (2007) greater than in the control treatment (P < 0.0001). Leaf number for control plants was 107% greater than in zone A in 2006 (P < 0.0001). However, the opposite trend was found in 2007 when plants in zone A had 200% more leaves than control plants (P = 0.0008). Eggplant height also varied between zones, although the trend differed between years. In 2006, mean height in zone A was 13 cm less than in control plots (P < 0.0001), whereas in 2007, mean plant height in zone A was 11 cm greater than in control plots (P < 0.0001). In 2006, mean leaf number in the control plots was 102% greater than in zone A (P < 0.0001), whereas in 2007, leaf numbers in zones B and C were only half that of control plants (P = 0.0011).
Crop yield
In both years, yields for pearl millet were significantly higher in control plots and zone C than in zones situated closer to the trunk (Table 3). In the zone closest to the trunks, yield was less than a third of that for control plants and was ca. 10% of the control value in 2007 (P < 0.0001).
The yields of chilli pepper and eggplants were almost zero in 2007 and were not recorded. However, in 2006, eggplant resembled pearl millet in that the total weight of fruits harvested close to the trees was lower than in the control plots (P = 0.0001). By contrast, a considerable increase in fruit production was obtained when chilli pepper was grown as an under storey crop in 2006 (Table 3). Fruit yield in zones A and B was higher than in zone C and the control plots (P = 0.0001). Similarly, taro produced higher yields in zones A and B in both years and values close to the trunk were approximately double those outside the tree canopy (P = 0.0049 in 2006 and P = 0.0040 in 2007).
Chlorophyll fluorescence
For all crops, the maximum quantum yield of PSII photochemistry (Fv/Fm = ΦP0) was significantly higher under the tree canopies than in the open (Fig. 3). A similar trend was observed in 2007 for taro (P < 0.01), eggplant (P < 0.01) and millet (P < 0.01) but not for chilli pepper, for which the differences were not significant (data not presented). The difference in maximum quantum yield between the four crops was also significant (P < 0.0001), with eggplant having the highest average value (0.54), followed by chilli pepper (0.49), taro (0.48) and pearl millet (0.41). The differences in ΦP0 between chilli pepper, taro and millet were not significant. Measurements taken at different times on the same day in 2007 showed no significant effect of time on chlorophyll fluorescence for any of the crops.

Maximum quantum yield of PSII photochemistry (Fv/Fm) of crops in 2006 as a function of the concentric zones around individual trees and in control plots at Nobéré in Burkina Faso, West Africa. Error bars represent single standard errors of the mean; values with different letters in the same graph are significantly different (P < 0.05)
Soil chemical properties and soil organic matter
The presence of P. biglobosa seemed to influence soil properties (Fig. 4). Soil organic matter content (B), total soil N(C) and total P(D) content all tended to be higher in the topsoil of Zones A and B than in Zone C and the control plots. Total P content in deeper horizons (25–50 cm) and total K (E) content did not vary between concentric zones. Soil pH (A) became more acidic with increasing distance from the tree trunks. However, firm conclusions are difficult to draw as these analyses were not replicated due to financial constraints.

Soil pH (water), soil organic matter (%), soil total N (%), total P, and total K as a function of soil depth for each concentric zone at Nobéré in Burkina Faso, West Africa
Discussion and conclusion
In the Sahelian countries trees in fields are generally associated with higher soil fertility, higher soil water content and lower soil temperatures under the trees (see Breman and Kessler 1995 and Boffa 1999 for reviews). However, under semi-arid tropical conditions the effect of trees on associated crops is often negative. C4 plants, especially, including many tropical cereal crops, tend to have poor performance under tree shade (Kessler 1992, Boffa et al. 2000, Bayala et al. 2002, Ong et al. 2002), despite their ability to perform well in high temperatures (e.g. Pearcy et al. 1981, Ehleringer and Pearcy 1983). Even though millet yields in the Sahel are limited by a lack of nutrients (especially P and N, e.g. Fofana et al. 2008), the favorable soil fertility is not reflected by higher yields. In our study, the productivity of pearl millet was negatively affected when grown under P. biglobosa, as yields were decreased by up to 68% in 2006 and 89% in 2007 relative to control plants. This is comparable to results obtained by Maïga (1987), Kessler (1992) and Sanou et al. (2011) (64, 70 and 46%, respectively).
Interactions between C3 crops and trees in the Sahel have been less intensively studied. Photosynthetic responses of C3 plants to low irradiation may not necessarily be different from C4 plants (Ehleringer and Pearcy 1983, Pearcy and Calkin 1983), but C3 plants display a wider variation in their responses to sun exposure and high temperatures. In our study, we observed contrasting results between the three C3 crop types. Eggplant productivity decreased by up to 91% when grown close to trees, while chilli pepper yields in zones A and B were, respectively, 88 and 150% higher than in control plots in 2006. Similar to chilli pepper, taro yields obtained under the tree canopies were one to three times greater than in the control plots, consistent with results obtained by Ceasar (1980), Valenzuela et al. (1991) and Sanou et al. (2011), who reported that tuber dry weight in taro increased with decreasing light intensity.
Eggplant is believed to be of African origin but has undergone domestication in Asia (Daunay et al. 2001). Its presumed wild relative, S. incanum L., is a shrub found in dry to moist savannas in Africa (e.g. Gianoli and Hannunen 2000). Although shaded eggplant leaves show acclimation to low light conditions (Rosati et al. 2001), Uzun (2007) found decreasing yields at low light levels. The yield of tomato, a species closely related to eggplant, was also reported to decrease with decreasing light intensity (Dorais et al. 1991; Cockshull et al. 1992), and whether eggplant is truly a shade tolerant species is still widely debated. Chilli pepper originates from semi-deciduous forests in Mesoamerica, which is also where the first domestication took place (Aguilar-Melendez et al. 2009). We have not found references to the impact of shade on chilli pepper yields, but in a study on bell pepper (Capsicum annuum var. annuum) Roberts and Anderson (1994) found that yields increased when plants were moderately shaded. Jaimez and Rada (2006) grew the related sweet pepper (Capsicum chinense) in 40% shade in a humid tropical region and concluded that this species can be grown in partial shade. Finally, taro is believed to be of tropical rainforest origin and was probably first domesticated in Papua New Guinea (Denham et al. 2003). Its shade tolerance is therefore not surprising. Johnston and Onwueme (1998) concluded that, among five major tropical tuber crops, cocoyam (Xanthosoma sagittifolium) and taro were particularly well adapted to shade. They also found that leaves of taro grown in shade were larger and thinner than when cultivated in full sun (Onwueme and Johnston 2000); in our study, taro grown under trees produced an greater number of leaves. It thus seems that C3 plant reactions to shade, to some degree, reflect their role in natural ecosystems.
A major factor determining the impact of trees on associated crops appears to be competition for resources between crops and trees. Bayala et al. (2008) found that pruning of Parkia biglobosa and Vitellaria paradoxa increased yields of the cereals underneath considerably, indicating that release from competition would boost yields. A central question is whether above-ground or below-ground competition is the cause for the depressed yields. In another study from Burkina Faso, millet yields were positively correlated with light availability under the tree (Sanou et al. 2011). Unfortunately, analysis of the impact of trees on crops is complicated by covariation between environmental variables under the tree (e.g. Breman and Kessler 1995). Other negative influences of trees on associated crops may include competition for nutrients and water (Ong et al. 2002), though competition during the rainy season may be to the advantage of the crops as they develop a more intensive root system in the upper soil layers than the trees (Breman and Kessler 1995). Modeling indicates that a lack of light could, indeed, be responsible for reduced yields under trees (Bayala et al. 2007), but it is probable that different factors will be limiting in different environments (cf. Breman and Kessler 1995).
Plants adapted to shade usually have a low light compensation point, making them photosynthetically efficient in shade, but they tend to be less efficient when exposed to higher light levels (Kubiske and Pregitzer 1996). Shade adapted plants may even be harmed by high light levels and photoinhibition (Demmig-Adams and Adams 1992). Therefore we hypothesized that shade tolerant plants would be more photoinhibited than light demanding crops, and tested this by chlorophyll fluorescence measurements. The Fv/Fm ratio is correlated with the quantum yield of photochemistry in photosystem II (PSII) (Hormann et al. 1994), and in the dark-adapted state it is interpreted as the maximum photochemical yield of PSII (Baker 2008). For non-stressed plants, the ratio is relatively stable at 0.83 for a wide range of species (Björkman and Demmig 1987). In our study, average values were significantly below this value, indicating that plants in all zones were stressed (Baker 2008). The results also showed that all crop species examined tended to react in the same way, having relatively high Fv/Fm values under the canopy and low values outside. This indicates that plants were less photochemically stressed under the tree canopy than outside, and suggests a reduction in the potential activity of PSII and a decrease in the conversion efficiency of incident light outside the tree canopy. Despite being a C4 plant adapted to high irradiance and temperature, pearl millet reacted similarly to the other species and had a low Fv/Fm outside trees. Masojídek et al. (1991) also found a decrease in Fv/Fm in pearl millet plants subjected to high irradiance, particularly when the plants were drought stressed. It is possible that nutrient availability, high temperatures and water stress may limit photosynthesis outside the canopy, resulting in a similar photoinhibition of all crops (Demmig-Adams and Adams 1992; Vandenbelt and Williams 1992; Tomlinson et al. 1995; Boffa et al. 2000; Lott et al. 2009). This could be resolved by variation of fertilisation and irrigation in new tree/crop interaction experiments.
Our results indicate that taro may be suitable for cultivation under P. biglobosa, and that the micro-environment under P. biglobosa trees is suitable for producing high yields by certain adapted crops. Another crop with potential for cultivation under P. biglobosa may be chilli pepper, but it is too early to draw even preliminary conclusions based on yields from one season only. In any case, further testing of both species at other sites is desirable, in particular because of the large variation in yields from year to year. However, decisions on tree and crop combinations and planting density in parklands are seldom based on biological productivity itself, but are also influenced by farmers’ perceptions (Boffa et al. 2000). Based on results from this experiment, the use of eggplant as an under storey crop at Nobéré is questionable due to the observed yield decrease under tree canopy. Eggplant is, however, widely grown in the study area, both under trees and in open areas, and the fact that it has a very well established market is an important factor in making it an attractive crop for local farmers. Nonetheless, we cannot explain why farmers perceive it as drought tolerant.
Mertz et al. (2001) showed that the consumption of cultivated vegetables in southeastern Burkina Faso is based on a narrow range of species. This should also result in a limited array of crop choice for farmers. Extension services in Burkina Faso do not incorporate vegetables in their programs to a significant extent (Mertz et al. 2001) but, based on results presented here, we argue that promotion of crops such as taro for cultivation under P. biglobosa could increase agricultural production and thus improve the livelihoods of the local population.
References
Aguilar-Melendez A, Morrell PL, Roose ML, Kim SC (2009) Genetic diversity and structure in semiwild and domesticated chillies (Capsicum annuum; Solanaceae) from Mexico. Syst Phytogeogr 96:1190–1202
Allison LE (1965) Organic carbon. In: Black CA, Ivan DD, White JL, Clark SE (eds) Methods of soil analysis: chemical and microbiological properties. American Society of Agronomy, Wageningen, pp 1367–1375
Baker NR (2008) Chlorophyll fluorescence: a probe of photosynthesis in vivo. Ann Rev Plant Biol 59:89–113
Bayala J, Teklehaimanot Z, Ouedraogo SJ (2002) Millet production under pruned tree crowns in a parkland system in Burkina Faso. Agrofor Syst 54:203–214
Bayala J, van Noordwijk M, Lusiana B, Ni’matul K, Teklehaimanot Z, Ouedraogo SJ (2007) Separating the tree-soil-crop interactions in agroforestry parkland systems in Saponé (Burkina Faso) using WaNuLCAS. Adv Agrofor 4:296–308
Bayala J, Ouedraogo SJ, Teklehaimanot Z (2008) Rejuvenating indigenous trees in agroforestry parkland systems for better fruit production using crown pruning. Agrofor Syst 72:187–194
Björkman O, Demmig B (1987) Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 170:489–504
Boffa JM (1999) Agroforestry parklands in sub-Saharan Africa. FAO Conservation Guide, Italy, p 230
Boffa JM, Taonda SJB, Dickey JB, Knudson DM (2000) Field scale influence of karite (Vitellaria paradoxa) on sorghum production in the Sudan zone of Burkina Faso. Agrofor Syst 49:153–175
Breman H, Kessler JJ (1995) Woody plants in agro-ecosystems of semi-arid regions: with an emphasis on the Sahelian countries. Advanced series in agricultural sciences. Springer-Verlag, Berlin
Ceasar K (1980) Growth and development of Xanthosoma and Colocasia under different light and water supply conditions. Field Crop Res 3:235–244
Cockshull KE, Graves CJ, Cave CRJ (1992) The influence of shading on yield of glasshouse tomatoes. J Hort Sci 67:11–24
Daunay M-C, Lester RN, Ano G (2001) Eggplant. In: Charrier A, Jacquot M, Hamon S, Nicolas D (eds) Tropical plant breeding. CIRAD, Montpellier, pp 200–225
Day PR (1965) Particle fractionation and particle size analysis. In: Black CA, Ivan DD, White JL, Clark SE (eds) Methods of soils analysis: physical and mineralogical properties, including statistics, measurements and sampling. American Society of Agronomy, Wageningen, pp 545–566
Demmig-Adams B, Adams WW (1992) Photoprotection and other responses of plants to high light stress. Ann Rev Plant Physiol Plant Mol Biol 43:599–626
Denham TP, Haberle SG, Lentfer C, Fullagar R, Field J, Therin M, Porch N, Winsborough B (2003) Origins of agriculture at Kuk swamp in the highlands of New Guinea. Science 301:189–193
Dorais M, Andre G, Trudel MJ (1991) Annual greenhouse tomato production under sequential intercropping system using supplemental light. Scient Hort 45:225–234
Ehleringer J, Pearcy RW (1983) Variation in quantum yield for CO2 Uptake among C3 and C4 plants. Plant Physiol 73:555–559
FAO (1988) Non-timber uses of selected arid zone trees and shrubs in Africa. FAO Conservation Guide no. 19, Italy
Fofana B, Wopereis MCS, Bationo A, Breman H, Mando A (2008) Millet nutrient use efficiency as affected by natural soil fertility, mineral fertilizer use and rainfall in the West African Sahel. Nutr Cycl Agroecosyst 81:25–36
Gausset Q, Yago-Ouattara EL, Belem B (2005) Gender and trees in Péni, South-Western Burkina Faso. Dan J Geogr 105:67–76
Gianoli E, Hannunen S (2000) Plasticity of leaf traits and insect herbivory in Solanum incanum L (Solanaceae) in Nguruman, SW Kenya. Afri J Ecol 38:183–187
Gijsbers HJM, Kessler JJ, Knevel MK (1994) Dynamics and natural regeneration of woody species in farmed parklands in the Sahel region (Province of Passoré) Burkina Faso. For Ecol Manage 64:1–12
Hopkins HC, White F (1984) The ecology and chorology of Parkia in Africa. B Jard Bot Nat Belg 54:235–266
Hormann H, Neubauer C, Schreiber U (1994) On the relationship between chlorophyll fluorescence quenching and the quantum yield of electron transport in isolated thylakoids. Photosyn Res 40:93–106
Houba VJG, Van der Lee JJ, Walinga I (1985) Soils analysis. American Society of Agronomy, Wageningen, p 8
Jaimez RE, Rada F (2006) Flowering and fruit production dynamics of sweet pepper (Capsicum chinense Jacq.) under different shade conditions in a humid tropical region. J Sustain Agric 27:97–108
Johnston M, Onwueme IC (1998) Effect of shade on photosynthetic pigments in the tropical root crops: yam, taro, tannia, cassava and sweet potato. Exp Agric 34:301–312
Jonsson K (1995) Agroforestry in dry savannah areas in Africa: interactions between trees, soils and crops. PhD Dissertation, Swedish University of Agricultural Science, Umeå, Sweden
Kater LJM, Kante S, Budelman A (1992) Karité (Vitellaria paradoxa) and náé (Parkia biglobosa) associated with crops in south Mali. Agrofor Syst 18:89–105
Kessler JJ (1992) The influence of karite (Vitellaria paradoxa) and náé (Parkia biglobosa) trees on sorghum production in Burkina Faso. Agrofor Syst 17:97–118
Kubiske ME, Pregitzer KS (1996) Effects of elevated CO2 and light availability on the photosynthetic light response of trees of contrasting shade tolerance. Tree Physiol 16:351–358
Lericollais A (1989) La mort des arbres à Sob en pays Serer (Sénégal). In: Antheaume B, Blanc-Pamard C, Chaléard JL, Dubresson A, Lassailly-Jacob V, Marchal J-Y, Pillet-Schwarz AM, Pourtier R, Raison J-P, Sevin O, Pinton F (eds) Tropiques Lieux et liens: Florilège offert à Paul Pelissier et Gilles Sautter. ORSTOM, Paris, pp 87–197
Lott JE, Ong CK, Black CR (2009) Understorey microclimate and crop performance in a Grevillea robusta-based agroforestry system in semi-arid Kenya. Agric For Meteorol 149:1140–1151
Maïga A (1987) L’arbre dans les systèmes agroforestiers traditionnels dans la province de Bazéga. CNRST/IRBET, Ouagadougou, Burkina Faso
Masojídek J, Trivedi S, Halshaw L, Alexiou A, Hall DO (1991) The synergistic effect of drought and light stresses in sorghum and pearl millet. Plant Physiol 96:198–207
Mertz O, Lykke AM, Reenberg A (2001) Importance and seasonality of vegetable consumption and marketing in Burkina Faso. Econ Bot 55:279–289
Nair PKR (1993) An introduction to agroforestry. Kluwer Academic Publishers, Dordrecht
Ong CK, Wilson J, Deans JD, Mulayta J, Raussen T, Wajja-Musukwe N (2002) Tree-crop interactions: manipulation of water use and root function. Agric Water Manage 53:171–186
Onwueme IC, Johnston M (2000) Influence of shade on stomatal density, leaf size and other leaf characteristics in the major tropical root crops, tannia, sweet potato, yam, cassava and taro. Exp Agr 36:509–516
Pearcy RW, Calkin HW (1983) Carbon dioxide exchange of C3 and C4 tree species in the understory of a Hawaiian forest. Oecologia 58:26–32
Pearcy RW, Tumosa N, Williams K (1981) Relationships between growth, photosynthesis and competitive interactions for a C3 and C4 plant. Oecologia 48:371–376
Peech M (1965) Chemical and microbiological properties. In: Black CA, Ivan DD, White JL, Clark SE (eds) Methods of soils analysis. American Society of Agronomy, Wageningen, pp 915–926
Pleysier JL, Juo ASR (1980) A single extraction method using silver-thiourea for measuring exchangeable cations and effective CEC. Soil Sci 129:205–211
Roberts BW, Anderson JA (1994) Canopy shade and soil mulch affect yield and solar injury of bell pepper. Hortscience 29:258–260
Roháček K (2002) Chlorophyll fluorescence parameters: the definitions, photosynthetic meaning, and mutual relationships. Photosynthetica 40:13–29
Rosati A, Badeck FW, Dejong TM (2001) Estimating canopy light interception and absorption using leaf mass per unit leaf area in Solanum melongena. Ann Bot 88:101–109
Sanou J, Bayala J, Teklehaimanot Z, Bazié P (2011) Effect of shading by baobab (Andasonia digitata) and néré (Parkia biglobosa) on yields of millet (Pennisetum glaucum) and taro (Colocasia esculenta) in parkland systems in Burkina Faso, West Africa. Agrofor Syst. doi:10.1007/s10457-011-9405-4
Teklehainmanot Z (2004) Exploiting the potential of indigenous agroforestry trees: Parkia biglobosa and Vitellaria paradoxa in sub-Saharan Africa. Agrofor Syst 61:207–220
Tomlinson H, Teklehaimanot Z, Traore A, Olapade E (1995) Soil amelioration and root symbioses of Parkia biglobosa (Jacq.) Benth. in West Africa. Agrofor Syst 30:145–159
Uzun S (2007) Effect of light and temperature on the phenology and maturation of the fruit of eggplant (Solanum melongena) grown in greenhouses. N Z J Crop Hortic Sci 35:51–59
Valenzuela HR, O’Hair SK, Schaffer B (1991) Shading, growth and dry-matter partitioning of cocoyam (Xanthosoma sagittifolium (L.) Schott). J Am Soc Hortic Sci 116:1117–1121
Vandenbelt RJ, Williams JH (1992) The effect of soil surface temperature on the growth of millet in relation to the effect of Faidherbia albida trees. Agric For Meteorol 60:93–100
Wilson TO, Brook RM, Tomlinson HF (1998) Interactions between néré (Parkia biglobosa) and under-planted sorghum in parkland systems in Burkina Faso. Exp Agric 35:85–98
Acknowledgments
This study was funded by the European Community’s sixth framework program through the “Sahelian Fruit Trees” project (contract no. INCO-DEV 015465). We wish to thank the farmers of Nobéré for their permission and participation in the field study. We also wish to thank Yonli Hermann and Guibré K. Roger for field assistance. We are grateful for our anonymous reviewers’ helpful criticism and suggestions to an early version of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Pouliot, M., Bayala, J. & Ræbild, A. Testing the shade tolerance of selected crops under Parkia biglobosa (Jacq.) Benth. in an agroforestry parkland in Burkina Faso, West Africa. Agroforest Syst 85, 477–488 (2012). https://doi.org/10.1007/s10457-011-9411-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-011-9411-6