
Abstract
Despite the increasing claims that structural complexity in agroforestry plantations is a key variable for enhancing bird biodiversity, few studies have experimentally manipulated the understory component of structural complexity. Here, we examined the impact of removing understory vegetation from oil palm plantations on the richness and abundance of birds in eastern Guatemala, testing if bird richness and abundance are positively affected by the presence of understory vegetation. Oil palm plantations with understory hold more bird richness and abundance than those lacking it. The experimental removal of this vegetation, equivalent to a reduction in complexity, decreases bird richness but not abudance. Our analysis shows clear effects of vegetation removal on bird richness, such that removal reduces richness and abundance to levels found in areas without understory. Hence, leaving or implementing structural complexity within plantations could satisfy the current need of making commodity production a cleaner industry, fulfilling both, the production of highly demanded commodities and the conservation of biodiversity in productive landscapes. Oil palm (Elaeis guineensis) is one of the most rapidly expanding crop in tropical regions, representing the major cause of loss of natural forests and of the decline of endangered species in several countries. Enhancing understory vegetation might reduce such negative effects. This is one of the few experimental tests about how a component of the vegetation and structural complexity of plantations affects birds.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Land surface required to grow commodities has increased substantially to support increasing demands from human populations. If current trends continue unabated, 109 ha of natural ecosystems would be converted to agriculture by 2050 (Tilman et al. 2001). This land conversion is a significant driver altering Earth’s ecosystems, including biodiversity loss (Donald 2004; Foley et al. 2005). Therefore, a challenge to be met is to maintain the capacity of the biosphere to provide goods and services while conserving biodiversity. Wildlife-friendly farming is a current approach to simultaneously promote biodiversity conservation and agricultural production (Fischer et al. 2008). Our work deals with this concept, espcially since protected areas will not suffice to ensure long-term conservation of biological diversity, thus making it a challege to attempt it also in productive and semi-natural areas (Hartley 2002).
Plantations support some native biodiversity, playing a subsidiary role in conservation providing an occasional habitat, even for endangered species (Estades and Temple 1999; Daily et al. 2001; Simonetti 2006). Agricultural systems support native fauna depending on structural characteristics such as the existence of understory or multiple vegetation strata within the plantation, including prevalence of epiphyte or the presence of leguminous crops, as they can provide food and shelter for native species (Greenberg et al. 1997; Grez et al. 2003; Lindenmayer and Hobbs 2004; Aratrakorn et al. 2006). The presence of fauna in croplands is also affected by the proximity to native forests surrounding the plantation, and the size of forest remnants adjacent or inside plantations (Estades and Temple 1999; Koh 2008a).
Among commodities, vegetable oils and oil seeds have an increasing demand in global markets, not only for food industry, but for the increasing biofuel demand as well. Compared to other oleaginous crops, oil palm (Elaeis guineensis) has a very high yield per area and is one of the most rapidly expanding crops in tropical regions (Clay 2004; Koh and Wilcove 2008). The global area for oil palm cultivation has more than tripled, increasing from 3.6 million ha in 1961 to 13.2 million ha in 2006, an expansion rate of 246,000 ha/year occurring mostly in southeast Asia, Africa and Latin America (FAO-Food and Agricultural Organization 2007; Koh and Wilcove 2008). In the main producing countries (e.g. Indonesia, Malaysia, Colombia), oil palm plantations are the major cause of loss of natural forests and of the consequent decline of endangered species, such as the Sumatran Orangutan (Pongo abelii; Donald 2004). In general, fauna assemblages supported by oil palm plantations are depauperate in comparison to natural forests (Donald 2004; Aratrakorn et al. 2006; Fitzherbert et al. 2008; Koh and Wilcove 2008). Consequently, the production of palm oil, the top-selling vegetable oil in the World, has had a poor environmental record (Donald 2004). Given its economic importance and ubiquity in the food and oleochemichal business, as well as its current and projected expansion as a biodiesel feedstock, there is both a potential and a need to develop better practices to reduce its environmental impacts, including maintaining biodiversity within the plantations (Stone 2007; Turner et al. 2008), with initiatives such as the Round Table on Sustainable Palm Oil (RSPO-Roundtable on Sustainable Palm Oil 2005).
Despite the current demographic and economic trends, information to advance management techniques for maintaining biodiversity within the plantations is scarce at best. In fact, in the last decades, less than 1% of publications on oil palm are related to biodiversity and species conservation, and little research has been done to quantify the impacts on biodiversity of different management systems in oil palm plantations (Donald 2004; Koh 2008a; Turner et al. 2008).
The presence of a well developed understory is advanced as an enhancer of bird species richness in oil palm plantations (Aratrakorn et al. 2006), structural complexity being relevant for bird species richness in coffee and cocoa plantations as well (Van Bael et al. 2007; Gordon et al. 2007). Nevertheless, current information is correlational, and experimental demonstration has yet to be undertaken. In this framework, we experimentally assessed the importance of undergrowth for a bird assemblage in an oil palm plantation in Guatemala, by evaluating how understory, as a component of the vegetation complexity and structural complexity of plantations affects birds. If the understory significantly enhances bird species richness and abundance, these ought to be larger in a plantation with developed understory compared to those exhibiting nil or poorly developed vegetation. Further, bird species and abundance should decrease if understory is removed from plantations exhibiting such vegetation. Then, if the understory is important for birds in oil palm plantations, then we expect that modifying the understory will significantly impact bird species richness and abundance. To evaluate these hypotheses, our experimental approach consisted in modifying the undergrowth and analyzing bird responses to changes in understory development. As species response might be coupled to migratory habits, dietary guild and extinction vulnerability status, we also analyzed bird response according to these features (Pimm et al. 1988). Experimentally testing if bird biodiversity is enhanced in oil palm plantations with well developed understory will provide a clue for achieving biodiversity conservation outside protected areas rendering croplands more environmentally friendly.
Methods
Study area
Our study site was an oil palm plantation located at El Estor, eastern Guatemala. The plantation is located between two protected areas (Bocas del Polochic Wildlife Refuge and Sierra de las Minas Biosphere Reserve). The oil palm plantations in this region reach ~6,000 ha, producing 32 metric tons of fruit and 8 metric tons of oil per hectare, one of the highest productivities for any palm plantation worldwide (INDESA -Inversiones de Desarrollo- 2007).
Bird surveys
Bird surveys were carried out during January and February 2008, in 15 randomly selected plots under three different conditions: 5 plots without understory, 4 plots with understory vegetation that would remain unaffected and 6 plots that would have their understory later cleared. Plots with understory were those in which the undergrowth was dense and extensive, and plots without understory were those in which undergrowth was little or absent (sensu Aratrakorn et al. 2006). By understory, we meant, mostly, tall grasses or shrubs, mostly Fabaceae and Euphorbiaceae. Following a standard procedure (Ralph et al. 1999), point counts had a fixed ratio of 25 m and were located at least 100 m apart; we surveyed birds between 6:00 and 10:00 a.m., registering all bird species observed (only sight identifications were used) during sampling periods of 5 minutes.
Experimental design
To evaluate if bird diversity was affected by the understory vegetation in the plantation, our experiment evaluated bird richness and abundance before and after removing the understory from randomly selected plots, and then comparing the bird assemblages in the plots that had been cleared with control plots where vegetation was left intact (both with and without understory).
To establish the baseline before the undersory manipulation, we surveyed the 15 plots on three consecutive mornings. After this assessment, we removed the understory from six randomly selected plots (experimental plots), cutting all the vegetation and leaving them similar to those plots lacking understory (no-understory plots). Other four plots remained with the understory unmanaged (control plots). After 10 days, we conducted a second survey on three consecutive days, in the same hours as in the baseline sampling. Number of species and individuals were analyzed with repeated measures Analysis of Variance, and we performed planned comparisons between plots with understory versus without understory, with understory versus with experimental understory, and with experimental understory versus without understory.
To evaluate if species losses occurr in a particular order according to the different understory conditions, we assessed the nestedness of bird assemblages. A nested pattern emerges if bird richness and composition at a given treatment is actually a smaller subgroup contained in a larger assemblage (Ulrich and Gotelli 2007). We also evaluated if distance to native forest affected bird richness and abundance, comparing near versus distant plots (less or more than 200 m from forest edge respectively).
Results
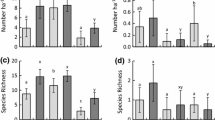
Twenty-three bird species from 14 families occurred at the oil palm plantation at El Estor. All but one inhabits oil palm plots with developed understory, and only 12 species are present in plots without understory (Table 1). Most species registered are resident (61%) and insectivores (48% of species). Regarding conservation status, 78% are regarded of Least Concern. Species never recorded in plots lacking undergrowth include two locally vulnerable species, Turdus assimilis and Malacoptila panamensis (Table 1; Eisermann and Avendaño 2006). Bird composition was not nested among experimental, control and no-understory plots (T = 20.4°, z = −0.54; p > 0.05), suggesting that within oil palm plantations species losses occur at random among plots of differing understory cover. The number of species expected in the pooled samples, given the empirical data, are close to what we observed in field (Mao Tau mean = 20 sd = 2.81), suggesting that in the sampled days we accumulated most of the avifauna that could be expected in the oil palm plantation.
Bird richness is 3.6 times higher in plots with understory than in those lacking it (F 2,12 = 10,0, p = 0.03; Fig. 1a). In the baseline count, the number of species recorded at experimental plots was similar to the control plots (p = 0.55), but 3.1 times higher than in plots without understory (p = 0.03). After undergrowth removal, the number of species in experimental plots decreased significantly by 41% (p = 0.01), but did not differ from those plots lacking understory (p = 0.09). Detectability of birds could be lower in plots with understory than in the cleared ones. If that were the case, we would have to have more bird records in plots without understory, but that did not happen, since even with an increased detectability, the cleared plots had less richness.

a Species numbers in all treatments before and after the manipulation of understory (Mean ± SE). The group “with exp” (with experimental understory) refers to the plots in which the undergrowth was removed. b Numbers of bird individuals in all treatments before and after the manipulation of understory (Mean ± SE). The group “with exp” (with experimental understory) refers to the plots in which the undergrowth was removed
Mean number of individuals was 3.2 times higher in plots with understory than in plots without understory. Differences are marginally significant though (F 2,12 = 3.4, p = 0.06; Fig. 1b). In the baseline count, the number of individuals in experimental plots was similar to control plots (p = 0.84), but marginlally higher (p = 0.06) than in the cleared plots. After undergrowth removal, the number of individuals in the experimental plots significantly decreased by 60% compared to control plots with understory (p = 0.02) but did not differ from plots without undersory (p = 0.93; Fig. 1b).
Distance from native forest affects neither bird species richness nor abundance. Mean number of species was similar among plots near (less than 200 m) and far away from the forest edge (over 200 m apart; t = 0.72; p = 0.48 and t = 0.90; p = 0.38), pre and post experiment, respectively. Similarly, bird abundance did not differ among plots near and far the forest edge prior to (t = 0.95; p = 0.35) or after the experimental removal of the understory (t = 0.53: p = 0.59). Plots were also indpendent among them (Mantel r = −0.58, p < 0.05). Therefore, both bird richness and abundance are higher in vegetated than in oil plam plots without understory and the number of species and individuals declined after the understory was removed in the experimental plots.
Discussion
The opportunity for setting aside new or enlarging existing protected areas—particularly in the tropics—is increasingly limited; beside the challenge to support current parks and reserves, a new quest is to achieve biodiversity conservation in productive landscapes (Hartley 2002). Turning croplands environmentally friendlier therefore is a step toward this goal. Oil palm plantations are reputed as harsh upon biodiversity, diminishing richness of several groups such as beetles, butterflies and birds (Chung et al. 2000; Koh 2008a). The existence of a well developed understory could reduce this impact, contributing to conserve some biodiversity (Aratrakorn et al. 2006). In fact, the experimental manipulation of the understory abundance demonstrates that enhancing such vegetation might have significant benefits for birds.
Understory vegetation beneath oil palms, as set forward by several authors (e.g. Aratrakorn et al. 2006) in fact promotes bird richness and abundance. Understory could be providing food resources, refuge and breeding sites for birds and other species, as occurs in other commercial plantations such as coffee, pine, eucalyptus, cacao and rubber. At these plantations, understory is also suggested as a significant source of food and shelter for species, thus enhancing biodiversity in these productive areas (Greenberg et al. 1997; Grez et al. 2003; Aratrakorn et al. 2006; Harvey and González-Villalobos 2007).
At El Estor, the undergrowth and the resources it might be providing, benefits even vulnerable species (T. assimilis and M. panamensis), two birds reported to live in primary forests only (Eisermann and Avendaño 2006). Guatemala resident birds appear to be more affected by the absence of understory than migrant species. Nine out of eleven missing species at understory devoided plots are resident ones, including the two vulnerable species, suggesting that they are more prone to become locally extinct than migratory species, contrary from what is usually assumed (Pimm et al. 1988).
Commercial plantations support only a fraction of the biodiversity that was once held in the natural ecosystems they replaced. Oil palm for instance supports about 10% of the original assemblage (see Donald 2004; Aratrakorn et al. 2006; Koh and Wilcove 2008). Adopting management procedures to enhance biodiversity within plantations may not be neither technically difficult nor financially costly as it might seem, as biodiversity within plantations could provide environmental services for the community and the industry owners (Turner et al. 2008). Current evidence indicates that agroforestry practices could offer a number of ecosystems services and environmental benefits, in addition to alleviating poverty (Jose 2009).
The dominance of insectivorous species (70% of species feed on insects) could be a key issue to promote the development of understory vegetation in oil palm plantations. Herbivorous insects are usually controlled by pesticides but currently there is a tendency to adopt biocontrol procedures. Several plant species are planted in order to attract insect predators and parasitoids in order to reduce insect abundance, precluding pest outbreaks (Koh 2008b). Allowing the spontaneous development of understory vegetation might also contribute to insect control as bird richness and abundance are enhanced in such plantations. Birds do reduce insect abundance (e.g. Koh 2008b), hence as with other plant species, insectivorous birds might contribute to the natural pest control, strengthening justifications for conserving biodiversity in this agricultural landscape (Koh 2008b).
Further, oil palm plantations with enhanced understory might even function as corridors between natural ecosystems. At El Estor, such management could favor the connectivity among bird populations at the two protected areas set apart by the plantations, hence integrating productive areas into current conservation efforts. Thus, leaving or building up understory vegetation at oil palm plantations ought to be encouraged. A missing point that must be urgently unraveled is whether such vegetation affects the productivity of plantations. Regarding these changes in production, experimentes conducted in oil palm, showed that there were no significant differences in yield between oil palm plots underplanted with cocoa and the controls (without any understory; Amoah et al. 1995).
Vegetation multilayers in agricultural systems appear to be very important for achieving biodiversity conservation in productive lands, and a feasible management measure particularly if it does not comprise their yield. Oil palm plantations can actually become environmentally friendlier, by leaving or promoting understory within them, combined with undertaking other practices previously identified (RSPO-Roundtable on Sustainable Palm Oil 2005; Koh and Wilcove 2007; Koh 2008a). This practice could be another step toward satisfying the current need of making commodity production a cleaner industry, aiding in the biosphere’s sustainability.
References
Amoah FM, Nuertey BN, Baidoo-Addo K, Oppong FK, Osei-Bonsu K, Asamoah TEO (1995) Underplanting oil palm with cocoa in Ghana. Agrofor Syst 30:289–299
Aratrakorn S, Thunhikorn S, Donald P (2006) Changes in bird communities following conversion of lowland forest to oil palm and rubber plantations in southern Thailand. Bird Conserv Int 16:71–82
Chung AYC, Eggelteon P, Speight MR, Hammond PM, Chey VK (2000) The diversity of beetle assemblages in different habitat types in Sabah, Malaysia. Bull Entomol Res 90:475–496
Clay JW (2004) World agriculture and the environment. A commodity-by-commodity guide to impacts and practices. Island Press, Washington
Daily G, Ehrlich PR, Sánchez-Azofeifa A (2001) Countryside biogeography: use of human-dominated habitats by the avifauna of southern Costa Rica. Ecol Appl 11:1–13
Donald P (2004) Biodiversity impacts of some agricultural commodity production systems. Conserv Biol 18:17–37
Eisermann K, Avendaño C (2006) Diversidad de aves en Guatemala, con una lista bibliográfica. In: Cano E (ed) Biodiversidad de Guatemala, vol 1. Universidad del Valle de Guatemala, Guatemala, pp 525–623
Estades CF, Temple SA (1999) Temperate-forest bird communities in a fragmented landscape dominated by exotic pine plantations. Ecol Appl 9:573–585
FAO-Food and Agricultural Organization (2007) FAOSTAT. Food and Agricultural Organization of the United Nations, Rome. Available at: http://faostat.fao.org
Fischer J, Brosi B, Daily GC, Ehrlich PR, Goldman R, Goldstein J, Lindenmayer DB, Manning AD, Mooney HA, Pejchar L, Ranganathan J, Tallis H (2008) Should agricultural policies encourage land sparing or wildlife-friendly farming? Front Ecol Environ 6:380–385
Fitzherbert EB, Struebig MJ, Morel A, Danielsen F, Brühl CA, Donald PF, Phalan B (2008) How will oil palm expansion affect biodiversity? Trends Ecol Evol 23:538–545
Foley JA, DeFries R, Asner GP, Barford C, Bonan G, Carpenter SR, Chapin S, Coe M, Daily G, Gibbs HK, Helkowski JH, Holloway T, Howard E, Kucharik CJ, Monfreda C, Patz JA, Prentice IC, Ramankutty N, Snyder PK (2005) Global consequences of land use. Science 309:570–574
Gordon C, Manson R, Sundberg J, Cruz-Angoón A (2007) Biodiversity, profitability, and vegetation structure in a Mexican coffee agroecosystem. Agric Ecosyst Environ 118:256–266
Greenberg R, Bichier P, Cruz-Angon A, Reitsma R (1997) Bird populations in shade and sun coffee plantations in central Guatemala. Conserv Biol 11:448–459
Grez AA, Moreno P, Elgueta M (2003) Coleopteros (Insecta: Coleoptera) epígeos asociados al bosque maulino y plantaciones de pino aledañas. Rev Chil Entomol 29:9–18
Hartley MJ (2002) Rationale and methods for conserving biodiversity in plantation forests. For Ecol Manag 155:81–95
Harvey C, González-Villalobos JA (2007) Agroforestry systems conserve species-rich but modified assemblages of tropical birds and bats. Biodivers Conserv 16:2257–2292
Howell SNG, Webb S (1995) A guide to the birds of Mexico and Northern Central America. Oxford Unversity Press, Oxford
INDESA-Inversiones de Desarrollo (2007) IX Aniversario Inversiones de Desarrollo. General technical report. INDESA, Guatemala
IUCN-International Union for Conservation of Nature (2007) Red list of threatened species. UICN, Geneve. Available at www.iucnredlist.org. Accesed 12 May 2008
Jose S (2009) Agroforestry for ecosystem services and environmental benefits: an overview. Agrofor Syst 76:1–10
Koh LP (2008a) Can oil palm be more hospitable for butterflies and birds? J Appl Ecol 45:1002–1009
Koh LP (2008b) Birds defend oil palms from herbivorous insects. Ecol Appl 18:821–825
Koh LP, Wilcove D (2007) Cashing in palm oil for conservation. Nature 448:993–994
Koh LP, Wilcove D (2008) Is oil palm really destroying biodiversity? Conser Lett 1:60–64
Lindenmayer DB, Hobbs RJ (2004) Fauna conservation in Australian plantation forests—a review. Biol Conserv 119:151–168
Pimm SL, Jones HL, Diamond J (1988) On the risk of extinction. Am Nat 132:757–785
Ralph CJ, Geupel GR, Pyle P, Martin TE, DeSante DF, Borja M (1999) Handbook of field methods for monitoring landbirds. General technical report. PSW-GTR-144. Forest Service, U.S. Department of Agriculture, Pacific Southwest Research Station, California
RSPO-Roundtable on Sustainable Palm Oil (2005) RSPO principles and criteria for sustainable palm oil production. Public Release Version. Kuala Lampur
Simonetti JA (2006) Conservación de biodiversidad en ambientes fragmentados: el caso del bosque maulino. In: Grez AA, Simonetti JA, Bustamante R (eds) Biodiversidad en ambientes fragmentados de Chile: patrones y procesos a diferentes escalas. Editorial Universitaria, Santiago, pp 213–229
Stone R (2007) Can palm oil plantations come clean? Science 317:1491
Tilman D, Farigone J, Wolff B, D′Antonio C, Dobson A, Howarth R, Schindler D, Schlesinger WH, Simberloff D, Swackhamer D (2001) Forecasting agriculturally driven global environmental change. Science 292:281–292
Turner E, Snaddon J, Fayle TM, Foser W (2008) Oil palm research in context. Plos One 3:e1572
Ulrich W, Gotelli N (2007) Null model analysis of species nestedness patterns. Ecology 88:1824–1831
Van Bael SA, Bichier P, Ochoa I, Greenberg R (2007) Bird diversity in cacao farms and forest fragments of western Panama. Biodivers Conserv 16:2245–2256
Acknowledgments
We thank H. García and O. Nuñez, (Fundación Defensores de la Naturaleza) for their support and interest in this research. M.T. Milla, A. Pérez, E. Paná and A. Caal for their valuable help in the field, and R. Erales (INDESA), for granting permits to work in the palm plantation. Thanks to AGCI (Agencia de Cooperación Internacional de Chile) Fellowship for supporting A.N.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Nájera, A., Simonetti, J.A. Can oil palm plantations become bird friendly?. Agroforest Syst 80, 203–209 (2010). https://doi.org/10.1007/s10457-010-9278-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-010-9278-y