
Abstract
Fungi are the cause of numerous plant diseases. Leading plant pathogens include various species of the genera Curvularia and Bipolaris. In this study of 21 airborne isolates, seven species with pathogenic potential for rice crops were identified (Curvularia aeria, Curvularia clavata, Curvularia pallescens, Curvularia trifolii, Bipolaris australiensis, Bipolaris hawaiiensis and Bipolaris sorghicola). For all isolates, optimum temperatures for mycelial growth and germination of conidia were determined over the 10–40 °C range. All strains were mesophilic, and optimum temperatures for germination of conidia lay within the range favourable for colony growth. In addition to their practical application in protecting the rice crop, these findings are of ecological interest in that they improve awareness of the aeromycological biodiversity of the study area.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Since rice (Oryza sativa L.) is a staple cereal and a major component of the daily diet in Cuba (Barrios and Pérez 2005), it is essential to minimise damage to rice crops. The chief fungal pathogens affecting rice are dispersed via the airborne route; characterisation of the aerial ecosystem in rice-growing areas is therefore a valuable tool for integrated rice disease management (Almaguer et al. 2011).
A number of fungi have been identified as rice pathogens (Estrada and Sandoval 2004); leading pathogens include various Curvularia and Bipolaris species (Cardona and González 2008), such as Curvularia aeria, C. affinis, C. brachyospora, C. clavata, C. eragrostridis, C. lunata, C. pallescens, C. trifolii, Bipolaris australiensis, B. hawaiiensis, B. oryzae and B. sorokiniana, as well as species belonging to other genera including Drechslera spp, Exserohilum spp. and Alternaria spp. (Cárdenas et al. 2003). These fungi can directly affect the growing grain, giving rise to so-called “pecky grain” or “kernel spotting,” and may also affect other parts of the plant, thus facilitating the transmission and spread of the pathogen to the whole crop. Adverse effects include impaired seed germination, reduced seedling size and vigour, lower number of grains per head, and the formation of broken and chalky grains (Barrios and Pérez 2005).
Airborne Curvularia and Bipolaris spores have been recorded in various tropical and subtropical ecosystems (Picco and Rodolfi 2002; Picco et al. 2004; Sharma 2010). In Cuba, Curvularia is one of the main fungal rice pathogens, causing seedling blight, panicle blight, leaf spot and seed spot (Neninger et al. 2003; Estrada and Sandoval 2004).
Prior to the development of fungal disease in a crop, spores must be released, dispersed and deposited on the substrate; later, if environmental conditions are favourable, spores will germinate in the affected areas, forming infective structures (Agrios 2005). Temperature plays a key role in the germination of conidia and in subsequent mycelial growth, and thus influences both the onset and the severity of plant diseases. In many species of filamentous fungi, high temperatures (>25 °C) stimulate enzyme activity and favour the development of germ tubes and infective structures (Freire et al. 1998; Webster and Weber 2007). This is one reason why pecky grain is one of the most widespread fungal rice diseases in Cuba.
This study of airborne isolates identified potentially pathogenic species belonging to the genera Curvularia and Bipolaris, and determined optimum temperatures for colony growth and germination of conidia.
2 Materials and methods
2.1 Biological material
Sampling for airborne fungal spores was carried out in a paddy field in Bauta (Cuba) in 2007, using a Surface Air System (SAS) air sampler. A total of 21 isolates were selected, five belonging to the genus Bipolaris (B1 to B5) and sixteen to the genus Curvularia (C1 to C16). These strains are part of the fungal culture collection maintained by the University of Havana Biology Faculty (CCFB), storage under mineral oil.
2.2 Identification of isolates
Isolates of the two genera were identified according to criteria suggested by Ellis (1971), Sivanesan (1987) and Mena (2004). Species differentiation was based on colony morphology and analysis of morphobiometric characteristics including distribution of conidia on conidiophores, size, shape, colour, texture and number of septae. In all cases, observations were made on fresh preparations of young and mature areas of colonies (Rashid 2001; Cardona and González 2008; Motlagh and Kaviani 2008).
2.3 Optimum temperature for mycelial growth
A single 4-mm plug cut from the young area of culture dishes that had been incubated for 4 days at 30 °C was placed upside down in the centre of a new 90-mm Potato Dextrose Agar (PDA) plate. Plates were incubated for 7 days at a range of temperatures (10, 20, 25, 28, 30, 32, 35, 37 and 40 °C); three replicate dishes were prepared for each treatment. Growth was quantified by measuring colony diameters at 7 days (Taylor and Khan 2010).
2.4 Optimum temperature for germination of conidia
The 21 isolates were cultured in tubes containing Potato Dextrose Agar (PDA) and incubated at 30 °C for 7 days. Conidia were then harvested, suspended in 5-ml saline with 0.05 % Tween 80®, vortexed and resuspended in sterile distilled water. Suspensions were then incubated at different temperature (10, 20, 25, 28, 30, 32, 35, 37 and 40 °C) for 24 h. Percentage germination was determined by counting total spores and germinated spores in ten visual fields of 50 μl of each suspension. A conidium was considered to have germinated if it had a germ tube at least as long as the smallest diameter of the conidium (Yeo et al. 2003).
2.5 Statistical analysis
Data for colony radial growth at different temperatures were tested for normality (Kolmogorov–Smirnov) and equality of variance (Bartlett). A one-way ANOVA was used to test for significant differences in radial growth for the same strain at different temperatures, and means were compared using a Tukey test. Differences in conidial germination rates for the same strain at different temperatures were examined by means of the percentage comparison test. All tests were carried out using the STATISTICA v. 6.0 software package for Windows.
3 Results
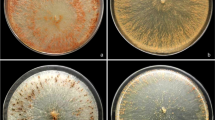
Four Curvularia and three Bipolaris species were identified. Table 1 shows morphological characteristics for Curvularia trifolii (Kauffman) Boedijn (7 isolates), C. aeria (Bat. J. A. Lima and C. T. Vasconc) (4 isolates), Curvularia clavata B.L. Jain (2 isolates) and Curvularia pallescens Boedijn (3 isolates). All isolates displayed smooth, mostly three-septate conidia; the central septum was not thickened or situated in the medial portion of the conidium. Table 2 shows morphological characteristics for the three Bipolaris species identified: B. australiensis (M. B. Ellis) Tsuda and Ueyama (2 isolates), Bipolaris hawaiiensis (M. B. Ellis) Uchida & Ueyama (2 isolates) and Bipolaris sorghicola (Lefebvre and Sherwin) Alcorn (1 isolates). All species had smooth conidia.
Colony radial growth for Curvularia strains at different temperatures is shown in Fig. 1. The optimum temperature ranges for growth of C. trifolii, C. aeria and C. pallescens were 25–35 °C, 20–35 °C and 25–30 °C, respectively. C. clavata strain C7 displayed significantly greater growth at 28 °C than at the other temperatures tested, whilst strain C15 of the same species attained optimum growth at between 25 and 35 °C. None of the colonies exhibited radial growth at 40 °C, except C. aeria strains C12 and C16, and to a lesser extent C. clavata strains C7 and C15, although growth in the latter (3–7 mm) was significantly lower than at the other temperatures.

Growth species of the genus Curvularia in PDA medium at different temperatures
For the two B. australiensis strains, growth was significantly greater at 30 °C than at the other temperatures tested (Fig. 2), whilst the optimum temperature range for the growth of B. hawaiiensis strains was 30–35 °C for B1 and 25–32 °C for B4. Bipolaris sorghicola strain B5 displayed significantly greater growth at 28 °C than at the other temperatures tested.

Growth species of the genus Bipolaris in PDA medium at different temperatures
Conidial germination rates for Curvularia strains at different temperatures are shown in Table 3. For all strains, the highest germination rate was observed at 28 °C, rates being statistically higher than those recorded at the other temperatures tested; the only exceptions were the three strains of C. trifolii (C3, C4 and C13) which displayed optimum germination temperatures of 37, 30 and 32 °C, respectively.
Conidial germination rates for B. hawaiiensis and for B. australiensis strain B3 also peaked at 28 °C, whilst the optimum temperature for germination in B. australiensis strain B2 was 30 °C. The maximum germination rate in B. sorghicola was recorded at 20 °C (Table 4).
4 Discussion
There have not been many aerobiological studies linked to rice growing in Cuba, and no reports have hitherto been published on potential airborne pathogens in rice agroecosystems, even though it is clear from the literature that conidial dispersal takes place largely through the airborne route and that these fungal species are major pathogens for cash crops.
Of the Curvularia species identified in this study, the most abundant was C. trifolii (7 isolates), which is commonly found on a wide range of plants and in various ecosystems; in Cuba, it has been isolated on the leaves of Clusia rosea, the seeds of Rottboellia cochiniensis, the leaves and seedlings of Saccharum oficinarum and the leaves of Zea mays (Mena 2004). Neninger et al. (2003) also report C. trifolii, C. pallescens and C. clavata as part of the pathogenic mycobiota in rice crops.
Curvularia aeria (4 isolates), a plurivorous species, has been isolated in the air and on various plants in the tropics, including several gramineae; in rice, it causes seed discolouration. Curvularia pallescens (3 isolates) is a cosmopolitan fungus classified as a saprobe and minor pathogen on various gramineous hosts, including several cereals (Mena 2004). Estrada and Sandoval (2004) first reported C. trifolii, C. aeria and C. pallescens as the cause of leaf spot and seedling blight in rice in Cuba, whilst Huang et al. (2005) have identified C. trifolii and C. pallescens as the causal agent of leaf spot in China.
The main hosts of C. clavata (2 isolates) are Oryza, Saccharum, Sorghum, Tripogon and Zea; it has been reported as the cause of various diseases in Oryza sativa and Sorghum spp (Niaz and Dawar 2009). Gutiérrez and Manzzanti (2002) identified C. clavata and C. aeria as causes of pecky grain in Argentina.
The Bipolaris species identified in the present study have also been reported as plant pathogens by a number of authors. Bipolaris australiensis (2 isolates) has been associated with pecky grain (Gutiérrez and Manzzanti 2002), and has also been isolated in other gramineae (Jones and Harrison 2004), in a wide range of dicotyledons, in the soil and in the air. It is the cause of leaf spot in Penisetum glaucum and leaf blight in Cymbopogon winterianus and is common in seeds (Mena 2004).
Bipolaris hawaiiensis (2 isolates) is a plurivorous species causing blight in the seeds and seedlings of a number of crops (e.g. Oryza sativa, Zea mays, Saccharum officinarum), although Pratt (2005) does not consider it a major rice pathogen. In Cuba, it is reported to cause seedling wilt in S. officinarum at nursery stage and is often encountered on the leaves both of this species and of other gramineae (Mena 2004). Both B. australiensis and B. hawaiiensis have been identified by Cárdenas et al. (2003) on rice grains in Cuban paddy fields.
Finally, although B. sorghicola (1 isolate) has not hitherto been identified as a rice pathogen, it is a major cause of disease in sorghum (Islam et al. 2009) and maize (Farr et al. 2005), and could well spread to other crops.
Analysis of optimum growth temperatures for the Curvularia and Bipolaris strains identified disclosed that all strains were mesophilic in terms of Cooney and Emerson’s classification criteria (1964): they did not display optimum growth at high temperatures (40 °C), and grew at below 18 °C (Salar and Aneja 2007). Whilst strains of both genera exhibited capacity for growth between 10 and 37 °C, the optimum temperature for the in vitro growth of Curvularia ranged from 20 to 35 °C, whilst for Bipolaris the optimum range was from 25 to 35 °C; similar optimum growth temperature ranges have been reported by Hassikou et al. (2003) and Freire et al. (1998).
Falloon (1975) notes that temperatures around 30 °C favour the in vitro growth of C. trifolii and that this is the optimum temperature for the development of seed spot in gramineae. Olufolaji (1985) also reports that the optimum temperature for the growth of mesophilic C. pallescens strains ranges from 25 to 28 °C. Differences between C. clavata strains C7 and C15 may reflect the fact that the physiological behaviour of each strain can differ considerably as a function of the timing of isolate collection, ecosystem characteristics and the source of the inoculum. Here, all strains were collected in the atmosphere of an agroecosystem; as a result, the inoculum source was both varied and indeterminate. Moreover, isolates were collected at different times of year, which may account for variations in the behaviour of some of the species studied.
Ojeda and Subero (2006) suggest that differences in the growth of Bipolaris species may also be due to the use of inocula from different sources. In a study of B. sacchari strains, Elliotta and Rayamajhib (2008) noted maximum growth rates over a range of temperatures similar to that observed here.
It should be stressed that the optimum growth temperatures recorded for the species studied are reached in the study area over most of the year; this favours the development of these pathogens (Agrios 2005), which thus pose a potential risk for rice crops both in the field and during post-harvest storage (Deshmukh and Gawai 2008).
The risk is heightened by the fact that, as the results of this study showed, the optimum temperature for conidial germination in most of the species studied was 28 °C, thus falling well within the range of optimum temperatures for colony growth. Within the range of optimum growth temperatures for Curvularia strains (20–25 °C), conidial germination rates average around 28 %; at a temperature of 28 °C, however, germination rates increase to 62 %. Similarly, the mean germination rate for Bipolaris conidia in the optimum temperature range for colony growth (25–35 °C) is 40 %; germination rates rise to 59 % at 28 °C.
Once the rice has been harvested, spores from these fungal pathogens may remain on grains, giving rise to infective structures favouring the development of post-harvest diseases, particularly since in Cuba rice is generally stored at ambient temperature (Neninger et al. 2003).
A review of the literature suggests that this is the first study of airborne Bipolaris and Curvularia isolates; the results are thus likely to be of particular value. The species studied are taxonomically and ecologically related and are major crop pathogens (Pratt 2005; Picco et al. 2004), so their identification and determination of the optimal conditions for their development can be a useful tool to implement phytosanitary measures against these pests.
5 Conclusions
Determining the optimum growth temperature of the species of the genera Curvularia and Bipolaris allowed the classification of all strains tested as mesophilic. The optimum temperature for germination of conidia of all strains was within the range favoured the growth, which is a potential risk in the development of post-harvest diseases depending on the conditions storage.
References
Agrios, G. N. (2005). Plant pathology (5th ed., pp. 386–500). Burlington, MA: Elsevier.
Almaguer, M., Rojas, T. I., Dobal, V., Batista, A., Rives, N., Aira, M. J., Hernández, A. N., & Hernández, A. (2011). Aerobiological dynamics of potentially pathogenic fungi in a rice agroecosystem in La Habana, Cuba. Aerobiologia. doi:10.1007/s10453-011-9222-2.
Barrios, L. M., & Pérez, I. O. (2005). Nuevos registros de hongos en semillas de Oryza sativa en Cuba. Manejo Integrado de Plagas y Agroecología, 75, 64–67.
Cárdenas, R. M., González, L., Parra, Y., Rivero, D., & Cruz, A. (2003). Influencia del manchado del grano de arroz (Oryza sativa L.) en la variedad J-104. nocividad y géneros de hongos presentes. Revista de Protección Vegetal, 18(2), 124–128.
Cardona, R., & González, M. (2008). Caracterización y patogenicidad de hongos del complejo Helmintosporium asociados al cultivo del arroz. Bioagro, 20, 141–145.
Cooney, D. G., & Emerson, R. (1964). Thermophilic Fungi. An account of their biology, activities and classification. San Francisco: Freeman and Company.
Deshmukh, A., & Gawai, D. (2008). Effect of temperature on incidence of storage fungi of some oil seeds. Journal of Pure and Applied Microbiology, 2(1), 261–264.
Elliotta, B., & Rayamajhib, N. (2008). Lower temperature during the dark cycle affects disease development on Lygodium microphyllum (old world climbing fern) by Bipolaris sacchari. Biological Control, 45, 56–63.
Ellis, M. B. (1971). Dematiaceous hyphomycetes. England: Commonwealth Mycological Institute. CAB International.
Estrada, G., & Sandoval, I. (2004). Patogenicidad de especies de Curvularia en arroz. Fitosanidad, 8, 23–26.
Falloon, R. E. (1975). Curvularia trifolii as a high-temperature turfgrass pathogen. New Zealand Journal of Agricultural Research, 19, 243–248.
Farr, D., Bills, G., & Chamuris, G. (2005). Fungi on plants and products in the United States. St. Paul, Minnesota: APS Press.
Freire, S. V. P., Paiva, L. M., Lima, E. A. L. A., & Maia, L. C. (1998). Morphological, cytological, and cultural aspects of Curvularia pallescens. Revista de Microbiologia, 29(3), 56–59.
Gutiérrez, S., & Manzzanti, M. A. (2002). Hongos asociados a granos manchados de Arroz. Arroz, 50, 22–25.
Hassikou, R., Hassikou, K., Ouazzanni, A., Badoc, A., & Doura, A. (2003). Biologie, physiologie et pouvoir pathogène de quelques isolats de Curvularia lunata agent de la curvulariose du riz. Bulletin de la Société Pharmaceutique de Bordeaux, 142, 25–44.
Huang, J., Zheng, L., & Hsiang, T. (2005). First report of leaf spot caused by Curvularia verruculosa on Cynodon sp. in Hubei. China Plant Pathology, 5, 253–257.
Islam, S. M. M., Masum, M. M. I., & Fakir, M. G. A. (2009). Prevalence of seed-borne fungi in sorghum of different locations of Bangladesh. Scientific Research and Essay, 4(3), 175–179.
Jones, A., & Harrison, R. (2004). The effects of meteorological factors on atmospheric bioaerosol concentrations—A review. Science of the Total Environment, 326, 151–180.
Mena, J. (2004). Taxonomía del complejo Bipolaris, Curvularia, Drechslera y Exserohilum en Cuba. Phd Thesis, Instituto de Ecología y Sistemática, CITMA. La Habana, Cuba.
Motlagh, M., & Kaviani, B. (2008). Characterization of new Bipolaris spp: The agent causal of rice brown spot disease in the North of Iran. International Journal of Agriculture and Biology, 10, 638–642.
Neninger, H., Hidalgo, E., Barrios, L. M., & Puedo, M. (2003). Hongos presentes en semillas de arroz (Oryza sativa L) en Cuba. Fitosanidad, 7(3), 7–11.
Niaz, I., & Dawar, S. (2009). Detection of seed borne mycoflora in maize (Zea mays L.). Pakistan Journal of Botany, 41(1), 443–451.
Ojeda, A., & Subero, L. (2006). Crecimiento y esporulación de Bipolaris oryzae (Breda de Hann) Shoem en diferentes medios de cultivo, condiciones de luz y temperatura. Mycopathologia, 13(1), 19–24.
Olufolaji, B. (1985). Comparative studies of the effect of Helminthosporium maydis and Curvularia pallescens infection on total nitrogen of maize leaves. Cryptogamie Mycology, 6(3), 97–200.
Picco, A. M., Lorenzi, E., Rodino, D., Rodolfi, M., & Biloni, M. (2004). Airspores detection or Pyricularia grisea (Cooke) Sacc. and Bipolaris spp. A three years monitoring in different rice fields in Northern Italy. Aerobiologia, 47(2), 133–137.
Picco, A. M., & Rodolfi, M. (2002). Pyricularia grisea and Bipolaris oryzae: A preliminary study on the occurrence of airborne spores in a rice field. Aerobiologia, 18(2), 163–167.
Pratt, R. G. (2005). Variation in occurrence of dematiaceous hyphomycetes on forage bermudagrass over years, sampling times, and locations. Phytopathology, 95(10), 1183–1190.
Rashid, M. (2001). Detection of Curvularia species on boro rice seeds of Dinajpur. Journal of Biological Sciences, 7, 591–592.
Salar, R. K., & Aneja, K. R. (2007). Thermophilic Fungi: Taxonomy and biogeography. Journal of Agricultural Technology, 3(1), 77–107.
Sharma, K. (2010). Seasonal variation of aeromycoflora over Ocimum sanctum plant with special reference to winter season. Journal of Phytology, 2(8), 1–5.
Sivanesan, A. (1987). Graminicolous species of Bipolaris, Curvularia, Drechslera, Exserohilum, and their teleomorphs. Mycological Papers, 158, 1–261.
Taylor, B. M., & Khan, A. (2010). Germination, radial growth and virulence of Metarhizium anisopliae and Paecilomyces fumoso-roseus on Bemisia Tabaci Gennadius (Homoptera: Aleyrodidae). Pakistan Entomologist, 32(2), 148–154.
Webster, J., & Weber, R. W. S. (2007). Introduction to Fungi. Cambridge: Cambridge University Press.
Yeo, H., Pell, J., Clark, S., & Pye, B. (2003). Laboratory evaluation of temperature effects on the germination and growth of entomopathogenic fungi and on their pathogenicity to two aphid species. Pest Management Science, 59, 156–165.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Almaguer, M., Rojas, T.I., Dobal, V. et al. Effect of temperature on growth and germination of conidia in Curvularia and Bipolaris species isolated from the air. Aerobiologia 29, 13–20 (2013). https://doi.org/10.1007/s10453-012-9257-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10453-012-9257-z