
Abstract
The aim of this paper was to summarize the current knowledge on how physical methods can reduce or control internal P release from sediments in lakes and reservoirs. Particular emphasis is given to the role of internal phosphorus load in fueling cyanobacterial blooms which are predicted to increase in frequency and intensity in response to climate change and eutrophication. We present selective case studies (both successful and unsuccessful) to assess the applicability and efficiency of major physical approaches used for decades to reduce internal loading in different systems of various morphology. In particular, we concentrate on where and when (1) hypolimnetic aeration/oxygenation, (2) hypolimnetic withdrawal and (3) sediment dredging are likely to reduce cyanobacterial blooms and whether these methods have an adverse impact on other organisms. We conclude that each method has its strength and weakness depending on the system considered. Sufficient knowledge of all lake nutrient sources and their dynamics together with detailed lake and sediment characteristics is an essential prerequisite for choosing an appropriate control method. We also report that many experiences demonstrated that a combination of restoration methods is often more successful than a single method.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
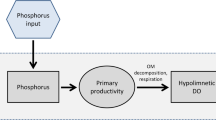
Cyanobacterial blooms are becoming more frequent and more intense as well as more widespread (Pearl and Huisman 2008; O’Neil et al. 2012) and create many adverse effects on ecosystems and human health including toxin exposures (Codd et al. 2005). Although the environmental factors (i.e., nutrients, light and temperature) controlling their onset and proliferation have been known for many decades, methods to control their development have been moderately successful. As phosphorus (P) is typically the limiting nutrient for productivity in lakes, mechanisms affecting P bioavailability have been well studied (Vollenweider 1975; Reynolds 1984; Wetzel 2001) although nitrogen plays a role in some shallow lakes (Sagrario et al. 2005; Pearl et al. 2011). While nutrients from catchments are partly in particulate form that settles in the water column and are not directly used by phytoplankton until released from particles, internal loads in lakes are predominantly in dissolved form directly available for algal growth. From this point of view, sediments are key components for considering in-lake measures to reduce phosphorus availability for cyanobacterial growth from internal load.
Preventive methods consisting of limiting nutrient inputs from external loading are without any discussion the long-term solution. However, they are often complemented by in-lake methods in order to accelerate the recovery process. Among them, physical, chemical and biological methods to reduce internal loading have been implemented in many waterbodies with relative success. Chemical methods (such as increase sediment sorption capacity by adding iron, aluminum or calcium) and biological methods (such as biomanipulation) will be discussed in separate papers in this special issue. In this paper, we review the applicability and relative success of major physical approaches used for decades to reduce internal loading in different systems of various morphology from representative case studies. While there are several excellent existing reviews dealing with factors influencing internal phosphorus loading in lakes (i.e., Søndergaard et al. 2001; Boström et al. 1988; Nürnberg 2009) or methods to control P release (Cooke et al. 2005), the main aim of this paper is to summarize the current knowledge on where and when physical methods can be used to control cyanobacterial blooms that are likely to increase in frequency and intensity in response to climate change and eutrophication.
We first discuss the direct and indirect links between internal loading and cyanobacterial blooms. After describing the working mechanism of each restoration method, we assess their relative successes and failures in controlling cyanobacterial blooms and try to understand the underlying reason. We then review the current knowledge on the ecological impact of each restoration method. Finally, we give some recommendations for choosing the appropriate physical control method.
Direct and indirect links between sediment phosphorus release and cyanobacterial growth
In shallow lakes, there is a direct link between the phosphorus release at the sediment water interface and the euphotic zone where photosynthesis takes place, as the water column remains vertically well mixed under the influence of wind-induced mixing (Søndergaard et al. 2003; Pettersson et al. 2010). In such systems, internal load often represents the main summer P load (Steinman et al. 2009; Nürnberg et al. 2012), which has a strong influence on algal bioavailability. Nürnberg et al. (2012) quantified internal phosphorus load over 26 years in a shallow mesotrophic lake and estimated that it was larger than external loads during years of high water temperature and low water load. Similarly, Steinman et al. (2009) conducted a study in a small eutrophic lake in Michigan and showed that in summer internal loading contributed 80 % of the total load. The increase in TP load in summer is often an indication of internal load. Interestingly, Istvánovics et al. 2002 reported on a positive feedback between high photosynthesis, elevated pH and P desorption from the resuspended sediments in the shallow Lake Balaton, highlighting that the cyanobacterium Cylindrospermopsis raciborskii was capable of generating high internal P release. Cyanobacteria are likely to dominate over other phytoplankton groups in these systems if the temperature is high (Pearl and Huisman 2008; Rigosi et al. 2014) and if the turbidity is also high. Indeed, high nutrient concentrations and high turbidity are often associated with cyanobacterial blooms in shallow systems, referred as one of the alternative stable state (Scheffer et al. 1997; Ibelings et al. 2007). High turbidity in shallow lakes is often associated with wind-driven sediment resuspension of both organic (algal biomass) and inorganic matter (suspended solid particles). Phlips et al. (1997), however, pointed out that if turbidity from inorganic particles becomes too high from wind resuspension, cyanobacteria productivity also becomes light limited.
In deep stratified lakes, a similar direct link may occur in the shallow parts (littoral zone) of the lake, which are well mixed and illuminated (Callieri et al. 2014) and where P release from sediments is in contact with the epilimnion (Søndergaard et al. 2005). Indirect link on the other hand occurs in the deeper parts (pelagic zone) of stratified lakes by mixing across the thermocline. Internal loading transfer to the epilimnion in stratified lakes occurs through turbulent vertical flux across the thermocline by mixing events, which entrain bottom waters through thermocline deepening (Soranno et al. 1997; Pannard et al. 2007; Pettersson et al. 2010), or by internal waves (MacIntyre et al. 1999; Bormans et al. 2004). Reported studies that quantify this entrainment are rare. Soranno et al. (1997) quantified the entrainment from hypolimnetic P to the budget of epilimnetic P in the deep eutrophic lake Mendota, USA, over a 2-year period. They showed that the entrainment potential depended on the mixing potential and the nutrient load potential. While Chl a concentration was similar in both years, cyanobacteria dominated the algal biomass with 85 and 96 %, respectively, over the 2 years. P entrained in the epilimnion from internal load represented up to 78 % of yearly total TP load. Entrainment events were related to storms and concentrated in early and late summer when the stratification was weaker in accordance with other studies (Pannard et al. 2011). Bormans et al. (2004) explained the persistence of a cyanobacterial bloom dominated by Cylindrospermopsis raciborskii from this entrainment of hypolimnetic high nutrients from wind-induced internal waves. Similarly, Pannard et al. (2011) demonstrated the close relationship between the metalimnetic Planktothrix agardhii bloom in a small Canadian lake and the entrainment of the nutrient-rich hypolimnion associated with internal waves.
In deep lakes in autumn and winter, mixing by convection redistributes phosphorus throughout the entire water column. Hence, there is generally a time lag between the high internal load generated predominantly in summer and its transfer into the epilimnion, which predominantly occurs in autumn. Interestingly, Sherman et al. (2000) have reported in a deep Australian reservoir a high correlation between the phosphorus concentration in the hypolimnion and the mean annual biomass of the cyanobacterial, suggesting a strong dependency on internal loading for cyanobacterial growth. Hence, considering internal loading from entrainment may help explain the observed variability in epilimnetic TP dynamics in stratified lakes. As opposed to other phytoplankton groups, cyanobacteria through the regulation of their buoyancy are well adapted to grow in strongly stratified lakes and reservoirs, gas vesicles allowing them to float and proliferate in the well-illuminated surface layers while other phytoplankton groups settle below the thermocline (Huisman et al. 1999; Bormans and Condie 1998). Cyanobacteria still require nutrients, which are more limiting in the surface layer in the summer than in the fall following overturn. Cyanobacteria are therefore the primary phytoplankton group to benefit from internal loading.
Hence, in both shallow and deep systems, the reduction of internal loading is likely to reduce the biomass of cyanobacterial blooms.
Expected impact of climate change on internal load
Climate change is now widely recognized to be one of the most distinct threats to aquatic ecosystems. The impacts of climate change on internal phosphorus loading have only been studied quite recently with particular attention to the effects of year-to-year variability in climate on the internal loads (Pettersson et al. 2010; Nürnberg et al. 2012). Jeppesen et al. (2007) discussed how climate change, in particular warming and drought, might influence the shallow lakes response to nutrient loading reduction. They suggested that the most probable outcome is that shallow European lakes will reach a more turbid state and in the south will become more saline as well. Expected increase in temperature and strengthening of the stratification leading to shallower surface mixed layer depth will likely increase the surface area of the sediments in contact with anoxic waters, hence enhancing phosphorus release from sediments (Schauser and Chorus 2009). Moreover, temperature increase will also lead to greater bacterial mineralization, which also should increase phosphorus release (Søndergaard et al. 2003). It is therefore expected that internal loading will be enhanced in response to climate change, and it is likely that cyanobacteria will be the first group to benefit from these increases as they are adapted to warm, turbid as well as stratified conditions (Rigosi et al. 2014; Mantzouki et al. 2016).
Control methods of P release by physical approaches
P release from sediments is influenced by many factors and processes that depend on the characteristics of the sediments and the water column, including redox potential, concentrations of Fe, Al and Ca, sediment organic matter and its reactivity, pH, temperature, resuspension, diffusion, microbial and bioturbation activities. Although internal loads are difficult to quantify due to the complexity of the many physical, chemical and biological processes involved, restoration methods to control them have been used for several decades. We now present the main physical techniques used to reduce P release from sediments and discuss selective representative case studies, which explicitly report on the restoration methods responses on cyanobacteria biomass.
Hypolimnetic aeration and oxygenation
Working mechanism
Hypolimnetic aeration or oxygenation is a technique used since the seventies to combat water quality problems associated with anoxia. Its primary goal is to raise or maintain dissolved oxygen (DO) at the bottom of lakes or reservoirs to suppress sediment release of reduced species (i.e., Fe, Mn, P and ammonia) by creating oxic conditions at the sediment–water interface. The technique intends to avoid complete destratification or warming the hypolimnion. In the particular case of controlling cyanobacterial blooms, the basic principle of hypolimnetic aeration/oxygenation system is to reduce phosphorus concentrations in the hypolimnion with minimum mixing across the metalimnion to avoid entrainment of nutrient-rich bottom waters into the epilimnion.
Aeration can be achieved by compressors that introduce air in the bottom of the lake through perforated tubes. An overview of various designs for hypolimnetic aerators is available in Cooke et al. (2005). They are grouped into three categories: (1) mechanical agitation, (2) injection of pure oxygen and (3) injection of air through a full lift or partial lift design or through a down-flow injection design. The hypolimnetic aeration may not operate satisfactorily if the water body is too shallow even if stratification exists, as the density gradient may not be sufficient to resist thermocline erosion while the hypolimnion is mixing. Procedures for sizing hypolimnetic aerators are described in detail by Lorenzen and Fast (1977), Ashley (1985) and Little (1995). The costs associated with this method are not very often reported in the literature. In Lake Amisk, capital costs were $30 k and operating cost were about $49 k per year (Prepas and Burke, 1997). Due to the need of the electric power, the operating costs of this method are relatively high. Moreover, aerators are usually put in spring and run during the whole summer season until autumn. Costs of oxygen injection estimated by Hickey and Gibbs (2009) were around $2.5 k/ha/year, while Cooke et al. (2005) reported overall costs for an average of 15 lakes in the USA of $3 k/ha/year.
Aeration systems are also used to completely mix the water column. In that configuration, although increasing O2 concentration in the hypolimnion is also an objective to reduce internal P loading, the complete mixing has other impacts (on light climate and buoyancy regulation) that affect cyanobacterial growth by other means than just reducing internal P loading. Hence, a separate article by Visser et al. (2016) in this special issue is specifically dedicated to artificial mixing as a method to control cyanobacterial blooms.
Successful and unsuccessful examples
Many examples of hypolimnetic aeration applications in lakes and reservoirs worldwide have been reported in the literature, and extensive reviews include Beutel and Horne (1999), Cooke et al. (2005) and Singleton and Little (2006). Here, we select studies that explicitly report on phytoplankton and cyanobacterial blooms responses to hypolimnetic aeration/oxygenation (Table 1). Moore et al. (2012) reported on a successful application of long-term hypolimnetic aeration on phytoplankton and cyanobacteria dynamics in Newman’s Lake (USA), a relatively shallow (z max = 9 m) dimictic eutrophic lake, where aeration was implemented since 1992 to control phosphorus recycling from the anoxic lake sediments. They stated that the system was capable of maintaining adequate oxygen to reduce internal loading but needed to stay in operation year round. They reported a phytoplankton community dominated by diatoms (Aulacoseira) with minimum cyanobacteria during operations and found a good correlation between phytoplankton biomass and TP concentrations with significant decrease over time. Cyanobacteria only contributed significantly to the biomass before 2000 when TP was higher than 40 μg/L. However, they also mentioned that external loads were not well monitored, and therefore, it was difficult to assess the efficiency of the hypolimnetic oxygenation. Toffolon et al. (2013) reported on a long-term (10 years) study of side stream pumping in Lake Serraia, Italy (z max = 18 m) that although increased DO was observed during operations, an increase in hypolimnetic temperature resulted from probable entrainment of hypolimnetic waters into the epilimnion. In response, phytoplankton biomass decreased and cyanobacteria were replaced by chlorophytes. This was explained by the overall reduction of P availability likely due to the reduction of the overall amount of the nutrients released from the sediments. Hypolimnetic pure oxygen was added year round during 1988–1993 to improve the water quality of Amisk Lake, a moderate-sized deep eutrophic lake in Alberta Canada (z max = 34 m) (Prepas and Burke 1997). Pretreatment hypolimnetic TP concentrations in summer reached 120 μg/L and Chl a 36 μg/L with the dominance of the cyanobacteria genera Aphanizomenon and Anabaena. During operations, DO remained above 5 mg/L in the hypolimnion and summer TP concentrations dropped to less than 50 % of pretreatments concentrations. This was achieved without disturbing the stratification but warming of the hypolimnion reached up to 3.6 °C. Epilimnetic TP decreased to 87 % of pretreatment mean summer values down to 27 μg/L and Chl a to 45 % down to 7 μg/L. Total phytoplankton biomass during summer plankton community shifted from cyanobacteria to a mixed assemblage of diatoms and cyanobacteria (Webb et al. 1997). The use of hypolimnion oxygen delayed the stratification onset and established a diatom population to persist longer and removed P from the water column so that less TP in the epilimnion became available for cyanobacterial growth. The low P release in Amisk Lake and similar hardwater lakes was due to low Fe:P ratio resulting from the lack of iron from the precipitation of Fe by sulfides as ferrous sulfides. Indeed, the iron to P molar ratio in the hypolimnion is 0.03, whereas a ratio of 2 is required for the adsorption of P to iron in oxic water (Manning et al. 1994). On the contrary, Burgi and Stadelman (2002) reported on unsuccessful responses with long-term phytoplankton composition changes in Lake Sembach, Germany, before and after reduction of external and internal loads implemented in 1984 when the lake was highly eutrophic. Improved oxic conditions decreased the remobilization of P from sediments and TP in the epilimnion during spring overturn decreased to 42 μg/L. General trends have shown decrease in dinoflagellates and chlorophytes and increase in cyanobacteria and diatoms with increase in carnivorous crustaceans and decline of herbivorous crustaceans. Zooplankton also migrated to deeper layers lowering the grazing pressure on algae. Surprisingly, Gächter and Müller (2003) concluded based on 10 years of data from the same lake that increased DO concentration from aeration did not reduce the P release from sediments during summer. They showed that irrespective of oxic conditions in the hypolimnion the sediment/water interface remained anoxic due to unchanged high sedimentation rates of organic matter, which could explain why oxygenation did not increase the P retention capacity of the sediment. The chemical characteristics of the sediments (very high P, vivianite) also contributed to prevent restoration success. Similarly, Schauser and Chorus (2009) reported no effect on phosphorus reduction and phytoplankton biomass in Lake Tegel, Germany, following several years of hypolimnetic aeration. They concluded that the improvement was entirely due to the reduction of external loading. In the same lake, (Lindenschmidt and Hamblin 1997) had demonstrated in a modeling study that the aerators used were in fact destratifying the water column and therefore enhancing the phosphorus flux to the epilimnion. In a comprehensive report, Chorus and Schauser (2011) highlighted that Lake Tegel P from sediments originated from the mineralization of recently sedimented organic matter acted as a sink annually but seasonally as a source and that redox-sensitive adsorption to Fe played a negligible role. Finally, partial and full lift hypolimnetic aeration were used in Medical Lake, USA (z max = 18 m), to improve water quality after a return to eutrophic conditions (Soltero et al. 1994). While both systems in place for 3 years during the growing season from spring to fall reduced TP concentrations in the hypolimnion to one-third of pretreatment values (with max of 50 μg/L), they also increased hypolimnetic temperature and had no effect on Chl a concentrations with averaged values around 5 μg/L, or on phytoplankton biomass. They attributed this lack of response to an undersized system in light of the increase biological oxygen demand (BOD) during aeration.
In summary, hypolimnetic oxygenation and aeration have generally increase DO in the hypolimnion even if it has increased the BOD. However, the reduction of P loading has not always reached expectations partly due to a lack of iron to bind all the P released. When P released was reduced, cyanobacteria biomass was always reduced and more so when hypolimnetic aeration/oxygenation did not entrain hypolimnetic waters into the epilimnion. Unsuccessful cyanobacterial responses reported have come from (1) aerators inadequately sized to account for increased BOD demand or such that increased diffusion of P into the epilimnion resulted in an enhanced cyanobacterial growth, (2) lack of iron to bind released P, (3) lack of external load control and (4) lack of sufficient time of operation. If these limitations are overcome, hypolimnetic aeration should reduce hypolimnetic phosphorus accumulation and internal cycling and ultimately reduce the development of cyanobacterial blooms.
Ecological impacts on other organisms
Adverse biological effects resulting from aeration have also been reported. The supersaturation of hypolimnetic water with N2 that might lead to a gas bubble disease in fish is a possible problem in some cases (Kortmann et al. 1994). Some biological beneficial effects also result from aeration. In particular, aeration allows zooplankton to swim deeper into the lake where they can hide from predators in the dark bottom during the day (McComas 2002). Additionally, the expanded aerobic environment should enhance growth and expansion of coldwater fish. In Lake St Mary, Canada, ecological benefits of hypolimnetic aeration included expansion of fish habitat from increased oxygen concentrations, increased visibility and enhanced density of zooplankton (Rieberger 1992). In Amisk Lake, crustaceous zooplankton increased in the deep waters, densities and biomass of benthic macroinvertebrates increased and Cisco expanded their distribution to deeper depths than untreated basins (Webb et al. 1997). To our knowledge, no studies have been reported on other biological organisms including bacteria.
Hypolimnetic withdrawal
Working mechanism
This technique is often used in reservoirs discharging water for power generation or which are dammed near the surface and allow bottom release waters. Although hypolimnetic withdrawal is generally used to control the temperature of the outflow from reservoirs through outlets at different depths, it can also be used in lakes as a control strategy for limiting cyanobacterial blooms through reduced phosphorus internal loading. Hypolimnetic withdrawal relies on the density stratification, which enhances the stability of the water column. It involves the removal of phosphorus from the hypolimnion with the water removal or occasionally its treatment before been returned to the lake. The decrease in phosphorus concentration is then thought to limit the growth of cyanobacteria, particularly in lakes or reservoirs where the internal phosphorus load dominates. This method ensures that bottom water residence time is reduced, as well as the availability of nutrients to the epilimnion by limited vertical mixing. However, hypolimnetic withdrawal has a tendency to store heat as it expands the epilimnion heated by solar radiation (Kennedy 1999) and as such decreases the thermal stability of the water column which can promote vertical entrainment of nutrients in the epilimnion (Effler et al. 1986). The amount of water withdrawn is limited to prevent excessive water-level variations when high-quality inflow is not available. Timing of hypolimnetic withdraw is best to coincide with higher summer internal P loads and before the onset of destratification in the fall.
There are several types of systems based on either passive siphoning (relying on the pressure head between the lake and the pipe outlet) or active pumping, with withdrawal depth very close to the bottom in the deepest area. This restoration method is mostly applied to small surface area lakes and to deep lakes (usually at least seasonally stratified) (Nürnberg 2007). The advantage of this method is relatively low capital cost and potentially long-term effectiveness. Costs estimated by Hickey and Gibbs (2009) range around $400 k and $10 k per treatment. Similar costs have been reported for systems in US lakes (i.e., $420 k for Lake Ballinger, $310 k for Lake Devil and $282 k for Pine Lake) by Cooke et al. (2005). Relatively low costs of operating and maintenance are an advantage of this method.
Successful and unsuccessful examples
Successful hypolimnetic withdrawal installation has been documented in the USA, Canada, Germany, Finland, Poland and Australia (Nürnberg et al. 1987; Dunalska et al. 2007; Keto et al. 2004; Cooke et al. 2005; Sherman et al. 2001). Maximum hypolimnetic TP concentration as well as epilimnetic TP concentration decreased in almost all the studied lakes. The longer the withdrawal was operating, the greater was the change in epilimnetic TP concentrations. Schauser and Chorus (2007) reviewed the performances of both external and internal control measures in two Berlin lakes. In Schlachtensee, an Olszewski pipe was operated yearly for a few weeks since the eighties to withdrawal the P-enriched hypolimnion water at the end of the stagnation period. The comparison between the P content in the lake, the external P load and the P withdrawn from the hypolimnion showed that the hypolimnion withdrawal had in most years only a minor effect on the total P budget of the lake (Schauser and Chorus 2009) and on the phytoplankton and cyanobacteria biomass (Chorus and Schauser 2011). The redox-sensitive P desorption from the sediments was found to be the main source of P in the hypolimnion. However, the strongly stable summer thermal stratification largely kept the desorbed P in the hypolimnion with minimal transfer to the epilimnion and therefore not directly accessible for phytoplankton growth. They reported a shift from cyanobacteria dominance to insignificant contribution when summer epilimnetic TP < 25 μg/L. Interestingly, they also clearly demonstrated that phytoplankton biomass decreases only below a threshold of TP = 50 μg/L when phytoplankton switched from light limitation to phosphorus limitation. Lehman et al. 2009 reported on the use of hypolimnetic withdrawal as a control method for cyanobacteria. They presented the growth response of Aphanizomenon flos-aquae and Microcystis aeruginosa in Ford Lake, Michigan, USA, following three very short term (1 week), hypolimnetic withdrawals, where they replenished oxygen by forced advection and destabilized the water column. Ford Lake is a eutrophic lake (z max = 11 m) supporting warm water fisheries, and lake manipulation was intended to lengthen the spring diatoms dominance and limit summer cyanobacteria growth. Typically Aphanizomenon flos-aquae dominate from July to early August when it is replaced by Microcystis aeruginosa through to September. Their results show that withdrawal of bottom water (at 11 m depth) led to higher NO3, an extended diatom bloom and the elimination of the Aphanizomenon bloom. It did not, however, decrease Microcystis biovolume. Nürnberg (2007) provided a comprehensive review of general patterns of lakes responses to hypolimnetic withdrawal applications (in 40 European and 8 North American lakes). The evaluation in terms of phosphorus concentration compared before and during treatments revealed a decrease in both hypolimnetic and epilimnetic summer TP with reduced differences between the concentrations in both layers. The evaluation of Chl a concentration changes was less often reported, and only 6 lakes presented explicit results. Among those, only 3 showed reduced biomass (associated with much reduced TP in the epilimnion), two unchanged (with moderate changes in epilimnion TP and still values higher than 50 μg/L) and one increase (associated with no TP change in epilimnion). Gächter (1976) suggested that the interruption of the metalimnetic flux of nutrients under hypolimnetic withdrawal was responsible for the disappearance of Planktothrix rubescens in Lake Mauensee, Switzerland. A similar decline of Planktothrix rubescens was also observed in a Slovenian lake as a result of reduced metalimnetic P flux from hypolimnetic withdrawal (1980–1982) together with reduction of external loads from sewage diversions (1982–1985) (Vrhovsek et al. 1985). Therefore, all these case studies suggest that the metalimnetic P flux is responsible for the intensity of the Planktothrix rubescens blooms observed.
In summary, hypolimnetic withdrawal has generally decreased the internal P loading at the sediment water interface but not always controlled cyanobacterial blooms (Table 1). In each case study, understanding the reasons of failure (and success) is crucial. Unsuccessful cyanobacterial responses presented here have resulted from (1) lack of external control, (2) TP concentrations above threshold for P limitation, (3) decreases in water column stability leading to bottom nutrient-rich waters to be entrained into the epilimnion.
Adverse effects on water quality and other organisms
There are several drawbacks to hypolimnetic withdrawal. Problems of large amplitude internal waves in reservoirs causing water quality problems downstream have been reported. In particular, a well-documented incident occurred in a Sydney water supply reservoir in 1998 where interface excursions of 35 meters created a 70-day supply shutdown following a water quality problem of Cryptosporidium contamination following large rainfall runoffs (Cox et al. 2003). Negative biological effects might also take place downstream due to the discharge of water with a lower temperature, higher nutrients, ammonia, hydrogen sulfide or other toxic compounds. To avoid this negative impact, mixing with epilimnetic water might be employed (Cooke et al. 2005). These adverse effects can also be attenuated by additional chemical phosphorus precipitation in the outlet (Chorus and Mur 1999). If low DO is expected in the outlet, then aeration equipment could be installed. Drawbacks include strong odor caused by hydrogen sulfite, lake water-level drop, if water treated, increase in temperature which may destabilize the water column causing mixing with surface water and transport of phosphorus from the hypolimnion to surface waters.
Methods that directly treat the source of the internal nutrients loads (i.e., the sediments) have also been widely used. We report on a non-chemical method of sediment treatments: sediment dredging.
Sediment dredging
Working mechanism
The primary aim of sediment dredging is to remove the surface sediment layer rich in toxic pollutants (i.e., heavy metals (Garcia-Orellana et al. 2011), or organic pollutants (Martinez and Hornbuckle 2011; Weston et al. 2002), thus decreasing their release from the sediments to the water column. In the last two decades, sediment dredging has also been used to reduce nutrient and cyanobacterial inoculum (Zhong et al. 2008; Kleeberg and Kohl 1999).
Successful and unsuccessful examples
Examples of both successful and unsuccessful control of cyanobacteria by sediment dredging have been reported in the literature and are summarized in Table 1. A typical example of successful restoration took place in Lake Trummen (Vaxjo, Sweden) with cyanobacterial control by sediment dredging. Until 1920, the lake was oligotrophic, but the increase in domestic sewage and industrial wastewater inputs resulted in progressive eutrophication followed by formation of cyanobacterial blooms, fish kills and oxygen demand (Eiseltova 1994). During the years of 1957–1958, the external sources of nutrients were restricted, but without expected effects with respect to water quality. Thus, Lake Trummen was dredged in 1970–1971. After this radical action, the cyanobacterial blooms disappeared and were replaced by a species-rich plankton community. The ecosystem as a whole recovered to a functional unit, characterized by a balance between production and decomposition, and Lake Trummen became available for a variety of recreation activities (Bjork et al. 2010).
On the contrary, unsuccessful remediation action took place in Vajgar fishpond, Czech Republic (Pokorny and Hauser 2002). Sediment removal from the Vajgar pond (44 ha) started in 1991 and finished in 1992. During this period, a total amount of 330.000 m3 of sediment was removed (whole cost was approximately $850 k). While during the first year after the removal of sediments, phytoplankton dynamics showed some signs of improvement; 2 years later, phytoplankton structure and dynamics returned to a state as that before the year of 1991. Indeed, although the internal load of Vajgar pond was reduced by the sediment removal, the external loading remained high and there was still regular import of phosphorus and phytoplankton from the intensely managed fishponds in the upper pond catchment (Pokorny and Hauser 2002). Kleeberg and Kohl (1999) reported up to 99 % reduction of P fluxes from dredged cores of 50 cm compared to intact cores under both aerobic and anaerobic conditions from a shallow eutrophic lake Groβer Muggelsee, Germany. In this lake, despite efforts of external load reductions, large cyanobacterial blooms of Limnothrix redekei, Planktothrix agardhii and Aphanizomenon flos-aquae appear every summer. The reduction of P release in the cores was not due to breakdown of organic matter with depth or reduced TP concentrations but to progressed diagenesis. Hence, they suggested that as P fluxes from sediments were mainly determined by P supply via sedimentation, a reduction of external P loading would be more efficient than dredging.
Lake Taihu is a very shallow hypereutrophic lake in China (zmean = 2 m) that frequently exhibits serious cyanobacterial blooms in summer (Pu et al. 1998; Pearl et al. 2015). Although external loads have been considerably reduced in recent years, eutrophication in Lake Taihu has not decreased as internal loading is delaying the effect (Jiang et al., 2008). The influence of sediment dredging of the top 30 cm of sediments from Lake Taihu, China, on the release of phosphorus was tested in a laboratory incubation experiment (Zhong et al. 2008). The sediment cores of the dredged sediments at monthly interval between December 2005 and June 2006 had a lower release potential (by 50 %) compared to the controls over a period of 6 months at temperatures close to in situ values. The porosity of the dredged cores was lower due to compaction with depth, and organic matter content was also lower. The authors suggested that dredging was a possible measure to control cyanobacterial blooms in Lake Taihu after external loading was efficiently reduced. Van der Does et al. (1992) also reported on the successful restoration of shallow lake Geerplas, the Netherlands, by dredging. The lake was characterized by a high internal loading, a reduction of external loading alone not sufficient for recovery. A removal by suction of the mud layer (from 1 to 100 cm) was achieved together with the positioning of screens to reduce horizontal mud transport during two attempts in 1989 and 1990–1991. Phosphorus concentrations in the lake decreased to 0.1 mg/L. A simultaneous reduction in 1990 of external loading from 0.38 down to 0.13 P m−2 y−1 did, however, hide the real cause of improvement.
Often studies on dredging have reported responses associated with combined restoration methods. The restoration of shallow Lake Cockshoot Broad, UK, started in the 1980’s. First, the lake was isolated from the sewage effluent contaminated River Bure in 1982, followed by sediment dredging. Sediment removal was combined with other in-lake methods such as local fish community removal, which resulted in chlorophyll-a concentration decreases and aquatic plants re-colonization (Moss et al. 1996). Sediment dredging was combined with a Phoslock® addition in an enclosure experiment in the Netherlands and resulted in reduced cyanobacteria, microcystin and soluble reactive phosphorus concentrations (Lurling and Faassen 2012).
In summary, dredging is a costly restoration measure. In addition to the cost of dredging at the site, dredging also raises the problem of sediment disposal and treatment, both aspects which are also costly and impact on the environment (Peterson 1981). Treated sediments can, however, be used as fertilizers, and the benefit gained from sales after treatment can slightly offset the overall cost. Disposal sites often require hard to obtained permits, especially in lowlands, and require chemical and toxicity testing of the disposed material (Cooke et al. 2005). Dredging should be selected only if internal loading is the dominant nutrient source, if sedimentation rates are low and if the depth associated with high concentration of labile P is known. All the case studies reporting unsuccessful control of cyanobacteria by dredging came from observations that external loading was not concomitantly controlled. Most successful cyanobacterial responses after dredging have been associated with other combined restoration methods (fish removal, inactivation by Phoslock, external control).
Adverse effects on other organisms
The environmental impact of dredging on fauna and flora in sediments is discussed frequently. Basically, disturbance of sediments may act against living organism in two ways: (a) through the physical changes (changes in gravel and organic matter content, direct removal of organisms, etc.) and (b) physicochemical changes since the disturbance of contaminated sediments may remobilize sediment-bound contaminants into the water column where they potentially impact a wide range of species (Burton and Johnston 2010). If a lake is dredged completely, 2–3 years may be required to reestablish benthic fauna (Cooke et al. 2005). Dredged cores have reduced significantly the benthic macrofauna diversity and density (Lewis et al. 2001). Changes in gravel content on the sensitivity of fauna communities have been reported by Cooper et al. (2011) who showed that the sensitivity of macrofaunal communities may increase as both the proportion of gravel increased and the level of natural physical disturbance decrease. Of course, the effects may vary with different communities colonizing different sites. For example, the study of Bolam et al. (2011) investigated in a field experiment the vertical migratory capability of temperate macroinvertebrate species. They found that the survival following sediment burial varied significantly across species. Although the release of toxic substances from resuspended sediment may occur, toxicological responses in the field appear to be rare. On the other hand, the accumulation of contaminants into the living systems is likely even for several months after dredging and disposal (Roberts 2012).
Conclusions and recommendations
The mechanisms responsible for the release of phosphorus from sediments are lake specific, but the dominant process in shallow wind exposed lakes is often resuspended while in iron-rich deeper lakes it is redox-sensitive release. Although accurate quantification of internal phosphorus loads is difficult to determine due to the many factors and processes, which are involved, numerous restoration techniques have been developed and tested.
The selected case studies reporting on both successful and unsuccessful responses to different physical restoration methods, summarized in Table 1, have demonstrated that an essential prerequisite for long-term success is the detailed understanding of the dominant processes at work and their spatial and temporal variability. Indeed, all the unsuccessful applications were associated with insufficient initial knowledge leading to inappropriate method selection or undersized system.
We therefore recommend the following criteria before choosing among the physical restoration methods:
-
1.
Define the critical phosphorus level needed to achieve the predicted outcome
-
2.
Assess the dynamics and relative role of internal loading compared to external loading
-
3.
Assess the sediment characteristics to determine whether internal loading can be controlled
-
4.
Quantify the link between internal load and cyanobacterial biomass
-
5.
Scale the restoration method as a function of the internal load and the size of the lake
-
6.
Evaluate the potential to cause adverse effects to aquatic biota
-
7.
Set a long-term monitoring program before, during and after the restoration
The above criteria are agreement with those proposed by Schauser et al. (2003) and Hickey and Gibbs (2009). Schauser et al. (2003) presented a decision support tool for the selection of an appropriate in-lake method to reduce internal P load. They highlighted that selection criteria should include the evaluation of the magnitude of the external versus internal load, the dynamics of the internal load and the lake morphology. Another decision support framework to choose appropriate management options for reducing internal loads, and thus cyanobacterial blooms, has been provided for New Zealand lakes that are more windy and mix more deeply than continental lakes (Hickey and Gibbs 2009). These authors also show that the choice is a function of the lake and sediments characteristics and of the remediation method characteristics. They also highlighted the need to evaluate the potential to cause adverse effects to aquatic biota. When a lake is being restored, we also recommend a long-term physical, chemical and biological monitoring program including phytoplankton, zooplankton, fish and benthic fauna. Indeed, a lake response to reduction in internal load will take time and will only be effective once external loading is also controlled. Moreover, a cyanobacterial response to reduced loading will only be effective below a threshold associated with cyanobacterial growth switching from light to nutrient limited. Long-term monitoring is required as climate variability together with climate changes will influence the success of the method selected.
Reviewing the different physical methods commonly used to control internal phosphorus release from sediments, we have summarized the applicability, the effectiveness in decreasing internal P load, the cost and the ecological impact on biota in Table 2. Hypolimnetic aeration and oxygenation are a short-term effective method in deep stratified systems where complete destratification is not recommended. It has a relatively high operating cost associated with the generally continuous mode of operation for successful applications. The ecological impacts are mostly beneficial for both zooplankton and fish habitat. Hypolimnetic withdrawal is a long-term low capital and low operating costs method appropriate for small deep stratified systems. It requires discharged water to be partly compensated by high-quality inflow to avoid problems associated with water-level variations and discharged water to be treated to avoid downstream water quality problems of low DO, high reduced elements and nuisance odors. Sediment dredging is a short term, extremely high cost (of removal, disposal and treatment) restoration method and therefore only practically applicable in small and shallow systems. While it directly removes the cyanobacterial inoculum and reduce the labile P content of the sediment, it can have negative impacts on benthic organisms. We conclude that each method has its strength and weakness depending on the system considered and that a combination of restoration methods is often more successful than a single method. It is important to keep in mind that internal phosphorus release is likely to increase from expected climatic changes. Control methods should therefore include expected increase in internal loading in their applicability and potential effectiveness.
References
Ashley KI (1985) Hypolimnetic aeration: practical design and application. Water Res 19:735–740
Beutel MW, Horne AJ (1999) A review of the effects of hypolimnetic oxygenation on lake and reservoir water quality. Lake Reserv Manag 15:285–297
Bjork S, Pokorny J, Hauser V (2010) Restoration of lakes through sediment removal, with case studies from lakes Trummen, Sweden and Vajgar, Czech Republic. In: Eiseltova M (ed) Restoration of lakes, streams, floodplains, and bogs in Europe: principles and case studies. Springer Publishing, pp 101–122
Bolam SG, Barry J, Bolam T, Mason C, Rumney HS, Thain JE, Law RJ (2011) Impacts of maintenance dredged material disposal on macrobenthic structure and secondary productivity. Mar Pollut Bull 62(10):2230–2245
Bormans M, Condie S (1998) Modelling the distribution of Anabaena and Melosira in a stratified river weir pool. Hydrobiologia 364:3–13
Bormans M, Ford PW, Fabbro L, Hancock G (2004) Onset and persistence of cyanobacterial blooms in a large impounded tropical river, Australia. Mar Fresh Res 55:1–15
Boström B, Andersen JM, Fleischer S, Jansson M (1988) Exchange of phosphorus across the sediment-water interface. Hydrobiologia 170:229–244
Burgi H, Stadelman P (2002) Change of phytoplankton composition and biodiversity in Lake Sempach before and during restoration. Hydrobiologia 469:33–48
Burton GA, Johnston EL (2010) Assessing contaminated sediments in the context of multiple stressors. Environ Toxicol Chem 29(12):2625–2643
Callieri C, Bertoni R, Contesini M, Bertoni F (2014) Water level fluctuations boost toxic cyanobacterial “oligotrophic blooms”. PLos One 9(10):e109529
Chorus I, Mur LE (1999) Preventive measures. In: Chorus I, Bartram J (eds) Toxic cyanobacteria in water. E & FN SPON, London and New York, pp 235–273
Chorus I, Schauser I (2011) Oligotrophication of lake Tegel and Schlachtensee, Berlin—analysis of system components, causalities and response thresholds compared to responses of other waterbodies. Technical report (45/2011) of the Federal Environment Agency (Umweltbundesamt)
Codd GA, Morrison LF, Metcalf JS (2005) Cyanobacterial toxins: risk management for health protection. Toxicol Appl Pharmacol 203:264–272
Cooke GD, Welch EB, Peterson SA, Nichols SA (2005) In: Cooke GD (ed) Restoration and management of lakes and reservoirs, 3rd edn. Taylor and Francis, Boca Raton, Florida
Cooper KM, Curtis M, Hussin WMRW, Frojan CRSB, Defew EC, Nye V, Paterson DM (2011) Implications of dredging induced changes in sediment particle size composition for the structure and function of marine benthic macrofaunal communities. Mar Pollut Bull 62(10):2087–2094
Cox P, Fisher I, Kastl G, Jegatheesan V, Warnecke M, Angles M, Bustamante H, Chiffins T, Hawkins P (2003) Sydney 1998—lessons from a drinking water crisis. J Am Water Works Assoc 95(5):147–161
Dunalska J, Wisniewski G, Mientki C (2007) Assessment of multiyear (1956–2003) hypolimnetic withdrawal from Lake Kortowskie, Poland. Lake Res Manag 23:377–387
Effler SW, Perkins MG, Brooks CM (1986) The oxygen resources of the hypolimnion of ionically enriched Onondaga Lake, NY, USA. Water Air Soil Pollut 29:93–108
Eiseltova M (ed) (1994) Restoration of lake ecosystems—a holistic approach, vol 32. International Waterfowl and Wetlands Research Bureau, Slimbridge, Gloucester, UK
Gächter VR (1976) Die Tiefenwasserableitung, ein Weg zur Sanierung von Seen. Swiss J Hydrol 38:1–29
Gächter R, Müller B (2003) Why the phosphorus retention of lakes does not necessarily depend on the oxygen supply to their sediment surface? Limnol Oceanogr 48:929–933
Garcia-Orellana J, Canas L, Masque P, Obrador B, Olid C, Pretus J (2011) Chronological reconstruction of metal contamination in the Port of Mao (Minorca, Spain). Mar Pollut Bull 62(8):1632–1640
Hickey CW, Gibbs MM (2009) Lake sediment phosphorus release management—decision support and risk assessment framework. NZ J Mar Freshw Res 43:819–856
Huisman J, Van Oostveen P, Weissing FJ (1999) Species dynamics in phytoplankton blooms: incomplete mixing and competition for light. Am Nat 154:46–68
Ibelings BW, Portielje R, Lammens EH, Noordhuis R, van den Berg MS, Joosse W, Meijer ML (2007) Resilience of alternative stable states during the recovery of shallow lakes from eutrophication: Lake Veluwe as a case study. Ecosystems 10:4–16
Istvánovics V, Somlyódy L, Clement A (2002) Cyanobacteria-mediated internal eutrophication in shallow Lake Balaton after load reduction. Water Res 36:3314–3322
Jeppesen E, Søndergaard M, Meerhoff M, Lauridsen TL, Jensen JP (2007) Shallow lake restoration by nutrient loading reduction—some recent findings and challenges ahead. Hydrobiologia 584:239–252
Jiang X, Jin X, Yao Y, Li L, Wu F (2008) Effects of biological activity, light, temperature and oxygen on phosphorus release processes at the sediment and water interface of Taihu Lake, China. Water Res 42:2251–2259
Kennedy RH (1999) Reservoir design and operation: limnological implications and management opportunities. In: Tundishi JG, Straskraba M (eds) Theoretical reservoir ecology and its applications. Backhuys Publishers, The Netherlands, pp 1–28
Keto A, Lehtinen A, Makela A, Sammalkorpi I (2004) Lake restoration. In: Eloranta P (ed) Inland and coastal waters of Finland. Palmenia Publishing, University of Helsinki
Kleeberg A, Kohl JG (1999) Assessment of the long-term effectiveness of sediment dredging to reduce benthic phosphorus release in shallow Lake Muggelsee (Germany). Hydrobiologia 394:153–161
Kortmann RW, Knoecklein GW, Bonnell CH (1994) Aeration of stratified lakes: theory and practice. Lake Reserv Manag 8:99–120
Lehman EM, McDonald KE, Lehman JT (2009) Whole lake selective withdrawal experiment to control harmful cyanobacteria in an urban impoundment. Water Res 43:1187–1198
Lewis MA, Weber DE, Stanley RS, Moore JC (2001) Dredging impact on an urbanized Florida bayou: effects on benthos and algal-periphyton. Environ Pollut 115:161–171
Lindenschmidt KE, Chorus I (1997) The effect of aeration on stratification and phytoplankton populations in Lake Tegel, Berlin. Arch Hydrobiol 139:317–346
Lindenschmidt KE, Hamblin PF (1997) Hypolimnetic aeration in Lake Tegel, Berlin. Water Res 31:1619–1628
Little JC (1995) Hypolimnetic aerators: predicting oxygen transfer and hydrodynamics. Water Res 29:2475–2482
Lorenzen MW, Fast AW (1977) A guide to aeration/circulation techniques for lake management. Ecol Res Ser EPA-600/3-77-004, US Environmental Protection Agency
Lurling M, Faassen EJ (2012) Controlling toxic cyanobacteria: effects of dredging and phosphorus-binding clay on cyanobacteria and microcystins. Water Res 46(5):1447–1459
MacIntyre S, Flynn KM, Jellison R, Romero JR (1999) Boundary mixing and nutrient flux in Mono Lake, California. Limnol Oceanogr 44:512–529
Manning PG, Murphy TP, Prepas EE (1994) Forms of iron and the bioavailability of phosphorus in eutrophic Amisk Lake. Can Mineral 32:459–468
Mantzouki E, Visser PM, Bormans M, Ibelings BW (2016) Understanding the key ecological traits of cyanobacteria as a basis for their management and control under expected environmental changes. Aquat Ecol. doi:10.1007/s10452-015-9526-3
Martinez A, Hornbuckle KC (2011) Record of PCB congeners, sorbents and potential toxicity in core samples in India Harbor and Ship Canal. Chemosphere 85(3):542–547
McComas S (2002) Lake and pond management guidebook. Lewis Publishers, Boca Raton, Florida
Moore BC, Christensen D (2009) Newman Lake restoration—a case study. Part I. Chemical and biological responses to phosphorus control. Lake Reserv Manag 25(4):337–350
Moore BC, Cross BK, Beutel M, Dent S, Preece E, Swanson M (2012) Newman Lake restoration: a case study Part III: hypolimnetic oxygenation. Lake Reserv Manag 28(4):311–327
Moss B, Stansfield J, Irvine K, Perrow M, Phillips G (1996) Progressive restoration of a shallow lake: a 12-year experiment in isolation, sediment removal and biomanipulation. J Appl Ecol 33(1):71–86
Nürnberg GK (2007) Lake responses to long term hypolimnetic treatments. Lake Reserv Manag 24:388–409
Nürnberg GK (2009) Assessing internal phosphorus load—problems to be solved. Lake Reserv Manag 25:419–432
Nürnberg GK, Hartley R, Davis E (1987) Hypolimnetic withdrawal in two North American lakes with anoxic phosphorus release from the sediment. Water Res 21(8):923–928
Nürnberg GK, LaZerte BD, Olding DD (2003) An artificially induced Planktothrix rubescens surface bloom in a small kettle lake in southern Ontario compared to blooms world-wide. Lake Reserv Manag 19:307–322
Nürnberg GK, Tarvainen M, Ventela A-M, Sarvala J (2012) Internal phosphorus load estimation during biomanipulation in a large polymictic and mesotrophic lake. Inland Waters 2(3):147–162
O’Neil JM, Davis TW, Burdford MA, Gobler CJ (2012) The rise of harmful cyanobacteria blooms: the potential roles of eutrophication and climate change. Harmful Algae 14:313–334
Paerl HW, Xu H, Hall NS, Rossignol KL, Joyner AR, Guangwei Z, Boqiang Q (2015) Nutrient limitation dynamics examined on a multi-annual scale in Lake Taihu, China: implications for controlling eutrophication and harmful algal blooms. J Freshw Ecol 30(1):5–24
Pannard A, Bormans M, Lagadeuc Y (2007) Short term variability in physical forcing in temperate reservoirs: effects on phytoplankton dynamics and sedimentary fluxes. Freshw Biol 52:12–27
Pannard A, Beisner BE, Bird DF, Braun J, Planas D, Bormans M (2011) Recurrent internal waves in a small lake: potential ecological consequences for metalimnetic phytoplankton populations. Limnol Oceanogr Fluids Environ 1:91–109
Pearl H, Huisman J (2008) Climate: blooms like it hot. Science 320:57–58
Pearl H, Hall NS, Calandrino ES (2011) Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic induced change. Sci Tot Environ 409:1739–1745
Peterson SA (1981) Sediment removal as lake restoration technique. EPA report-600/3-81-016, US Environmental Protection Agency
Pettersson K, George G, Noges P, Noges T, Blenckner T (2010) The impact of the changing climate on the supply and recycling of phosphorus. In: George DG (ed) The impact of climate change on European lakes, Aquatic Ecology Series, vol 4. Springer Publishing
Phlips EJ, Cichra M, Havens K, Hanton C, Badylak S, Rueter B, Randall M, Hansen P (1997) Relationships between phytoplankton dynamics and the availability of light and nutrients in a shallow sub-tropical lake. J Plankton Res 19(3):319–342
Pokorny J, Hauser V (2002) The restoration of fish ponds in agricultural landscapes. Ecol Eng 18(5):555–574
Prepas EE, Burke JM (1997) Effects of hypolimnetic oxygenation on water quality in Amisk Lake, Alberta, a deep eutrophic lake with high internal phosphorus loading rates. Can J Fish Aquat Sci 54:2111–2120
Pu PM, Hu WP, Yan JS, Wang GX, Hu CH (1998) A physico-ecological engineering experiment for water treatment in a hypertrophic lake in China. Ecol Eng 10:179–190
Reynolds CS (1984) The ecology of freshwater phytoplankton. Cambridge University Press, Cambridge
Rieberger K (1992) Metal concentrations in fish tissue from uncontaminated B.C. lakes. Water Quality Section, Water Quality Branch, Province of British Columbia, Victoria, B.C
Rigosi A, Carey CC, Ibelings BW, Brookes JD (2014) The interaction between climate warming and eutrophication to promote cyanobacteria is dependent on trophic state and varies among taxa. Limnol Oceanogr 59(1):99–114
Roberts DA (2012) Causes and ecological effects of resuspended contaminated sediments (RCS) in marine environments. Environ Int 40:230–243
Sagrario G, María A, Jeppesen E, Goma J, Søndergaard M, Lauridsen T, Landkildehus F (2005) Does nitrogen loading prevents clearer water conditions in shallow lakes at intermediate high phosphorus conditions? Fresh Biol 50:27–41
Schauser I, Chorus I (2007) Assessment of internal and external lake restoration measures for two Berlin lakes. Lake Reserv Manag 23:366–376
Schauser I, Chorus I (2009) Water and phosphorus mass balance of Lake Tegel and Schachtensee—a modeling approach. Water Res 43:1788–1800
Schauser I, Lewandowski J, Hupfer M (2003) Decision support for the selection of an appropriate in-lake measure to influence the phosphorus retention in sediments. Water Res 37:801–812
Scheffer M, Rinaldi S, Gragnani A, Mur LR, vanNes EH (1997) On the dominance of filamentous cyanobacteria in shallow, turbid lakes. Ecology 78:272–282
Sherman B, Whittington J, Oliver R (2000) The impact of artificial destratification on water quality in Chaffey Reservoir. Arch Hydrobiol Spec Issue Adv Limnol 55:15–29
Sherman B, Ford P, Hatton P, Whittington J, Green D, Baldwin D, Oliver R, Shiel R, Van Berkel J, Beckett R, Grey L, Maher B (2001) In: Sherman B (ed) The Chaffey Dam story, CSIRO Land & Water. Final report for the CRCFE
Singleton VL, Little JC (2006) Designing hypolimnetic aeration and oxygenation systems—a review. Environ Sci Technol 40:7512–7520
Soltero RA, Sexton LM, Ashley KI, McKee KO (1994) Partial and full lift hypolimnetic aeration of medical lake, WA to improve water quality. Water Res 28:2297–2308
Søndergaard M, Jensen JP, Jeppesen E (2001) Retention and internal loading of phosphorus in shallow eutrophic lakes. Sci World 1:427–442
Søndergaard M, Jensen JP, Jeppesen E (2003) Role of sediment and internal loading of phosphorus in shallow lakes. Hydrobiologia 506–509:135–145
Søndergaard M, Jensen JP, Jeppesen E (2005) Seasonal response of nutrients to reduced phosphorus loading in 12 Danish lakes. Freshw Biol 50:1605–1615
Soranno PA, Carpenter SR, Lathrop RC (1997) Internal phosphorus loading in Lake Mendota: response to external loads and weather. Can J Fish Aquat Sci 54:1883–1893
Steinman A, Chu X, Ogdahl M (2009) Spatial and temporal variability of internal and external phosphorus loads in Mona Lake, Michigan. Aquat Ecol 43:1–18
Toffolon M, Ragazzi M, Righetti M, Teodoru CR, Tubino M, Defrancesco C, Pozzi S (2013) Effects of artificial hypolimnetic oxygenation in a shallow lake. Part 1: phenomenological description and management. J Envir Manage 114:520–529
Van der Does J, Verstraelen P, Boers P, Vanroestel J, Roijackers R, Moser G (1992) Lake restoration with and without dredging of phosphorus-enriched upper sediment layers. Hydrobiologia 233(1–3):197–210
Visser PM, Ibelings BW, Bormans M, Huisman J (2016) Artificial mixing to control cyanobacterial blooms: a review. Aquat Ecol. doi:10.1007/s10452-015-9537-0
Vollenweider RA (1975) Input–output models with special reference to the phosphorus loading concept in limnology. Schweiz Z Hydrol 37:53–84
Vrhovsek D, Kosi G, Kralj M, Bricelj M, Zupan M (1985) The effect of lake restoration measures on the physical, chemical and phytoplankton variables of Lake Bled. Hydrobiologia 127:219–228
Webb DJ, Roberts RD, Prepas EE (1997) Influence of extended water column mixing during the first two years of hypolimnetic oxygenation on the phytoplankton community of Amisk Lake, Alberta. Can J Fish Aquat Sci 54:2133–2145
Weston DP, Jarman WM, Cabana G, Bacon CE, Jacobson LA (2002) An evaluation of the success of dredging as remediation at a DDT-contaminated site in San Francisco Bay, California, USA. Environ Toxicol Chem 21(10):2216–2224
Wetzel RG (2001) Limnology: lake and river ecosystems, 3rd edn. Academic Press, San Diego
Zhong J, You B, Fan C, Li B, Zhang L, Ding S (2008) Influence of sediment dredging on chemical forms and release of phosphorus. Pedosphere 18(1):34–44
Acknowledgments
The authors would like to thank Caroline Klavins for contributing to the literature search as part of her Master’s project in Hydro3 at the University of Rennes 1. This review contributes to a special issue on “Cyanobacterial blooms: ecology, prevention, mitigation and control” which has been initiated by the EU-CYANOCOST action ES1105. Part of this study was supported as a long-term research development project No. RVO 67985939 (Institute of Botany of the ASCR). We are grateful to the reviewers and the editor whose comments have contributed to significantly improve the original manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Guest editors: Petra M. Visser, Bas W. Ibelings, Jutta Fastner & Myriam Bormans/Cyanobacterial blooms. Ecology, prevention, mitigation and control.
Rights and permissions
About this article

Cite this article
Bormans, M., Maršálek, B. & Jančula, D. Controlling internal phosphorus loading in lakes by physical methods to reduce cyanobacterial blooms: a review. Aquat Ecol 50, 407–422 (2016). https://doi.org/10.1007/s10452-015-9564-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10452-015-9564-x