
Abstract
Ecosystem-based fishery management programs require a reliable estimate of the trophic positions of aquatic resources. Both stable isotope analysis (SIA) and stomach content analysis (SCA) have been used to estimate the trophic positions (TP) of aquatic systems, but few studies have compared results from both methods. To determine whether the two methods produced similar results, we used SIA (δ15N and δ13C), SCA (estimated with the TROPH routine) and data from FishBase to estimate the TP of 66 fish species in a subtropical estuarine system of the Gulf of California. SIA values ranged from 2.6 to 5.6, with 56 % of the species having a SIA value above 4.5, and 14 % of the species having a SIA value below 4. The SCA values ranged from 2.6 to 4.8, with 76 % of the species having a SCA value of 3–4. Overall, SCA results underestimated TP (including FishBase data), while SIA yielded better results, particularly if both δ15N and δ13C are used. The TROPH routine has oversimplified assumptions such as the same TP for all organisms in the same taxon, but if SIA is unavailable, SCA could be used, accompanied by knowledge of the TP of the most important prey items.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In most parts of the world, fishery science is still developed within the realm of population ecology (Quinn II 2003; Mangel and Levin 2005). Single-species population dynamic models are core tools in the stock assessment process. For instance, although shrimp trawl fisheries of tropical regions are multispecific fisheries, single-species models apply, as a high quantity of bycatch is captured and discarded (Madrid-Vera et al. 2007).
There is a current global consensus that fishery management must shift from its traditional single-species focus to a more holistic approach toward utilization of aquatic resources while maintaining fully functional ecosystems (Clark et al. 2001; Marasco et al. 2007). Achieving sustainable use of fishery resources requires management on scale of all the organisms subject to exploitation rather than at the scale of those targeted directly by the fisheries (Koen-Alonso 2007). Using this new perspective, trophodynamic models now address the joint dynamics of fishery resources. Fish trophic position (TP) is currently recognized as a useful indicator of human disturbance, and trends in the mean trophic positions of fishery landings are often used as a sustainability and marine biodiversity indicator (Pauly and Watson 2005; Branch et al. 2010).
The TP of aquatic resources is a prerequisite for understanding how aquatic systems function (Baird et al. 1991; Baird and Ulanowicz 1993; Pasquaud et al. 2010) and for developing ecosystem-based fishery management programs (Bondavalli et al. 2006; Ulanowicz 1996). TP information also enables comparative community analysis, in which available community studies are re-expressed using trophic position as a common value (Pauly et al. 2000b).
However, a reliable estimate of fish TP is needed. Programs such as the web-based relational database FishBase estimate TP with stomach content analyses (SCA) and ecological modeling tools such as TROPH (Pauly et al. 2000a, b). FishBase provides TP values for close to 33,000 fish species (as of March 2015) that were estimated using diet information extracted from publications and provides an adequate source of information on the trophic ecology of fish. However, SCA is time intensive and requires high sample numbers, as well as considerable taxonomic expertise, especially when investigating the diets of benthic predators. Additionally, SCA provides detailed information on fish diet but does not account for long-term patterns of mass transfer; instead, it provides an instantaneous measure of an organism’s diet (Vander Zanden et al. 1997). Finally, spatial–temporal variability in prey (e.g., abundance, availability and capture efficiency) and ontogenetic shifts (e.g., increases with size and energetic requirements) may obscure fish TP estimates within the food web based on SCA.
In recent years, stable isotope analysis (SIA) has been recognized as a useful tool for TP estimation as an alternative to or combined with SCA (e.g., Codron et al. 2012; Mancinelli et al. 2013). TP is generally achieved using only δ15N signatures or in combination with δ13C measurements (Cabana and Rasmussen 1996; Vander Zanden et al. 1999; Post 2002). Naturally occurring stable isotopes of nitrogen (15N/14N) and carbon (13C/12C) are also used to investigate food webs, specifically in determination of food or energy sources (DeNiro and Epstein 1981; Minagawa and Wada 1984; Wada et al. 1991), dietary patterns, and trophic relationships within ecosystems (e.g., Deegan and Garritt 1997; Post 2002; Pasquaud et al. 2010). Depending on the tissue analyzed, SIA measurements can provide a longer integrative record of food assimilated by an organism than SCA. However, SIA also has limitations related to spatial–temporal fluctuations in the isotopic compositions of available foods. Further, SIA presents the possibilities of overlapping isotopic values in dietary items and inter- and intraspecific variability in the isotopic signatures of predators due to seasonal effects, size-dependent feeding, and the tissue lipid content (Post 2002; Mancinelli et al. 2013). Additionally, it does not provide direct evidence of an organism’s prey items (i.e., specific invertebrate taxa) and requires infrastructure and instrumentation that are not always available in developing countries.
The objective of this study was to compare SCA and SIA estimates of TP values of 66 estuarine fish species from a subtropical estuarine system in the Gulf of California. Because fish TP is critical to assessing ecosystem-level biological interactions, we sought to determine whether the methods yielded similar results. Two versions of each method were compared, and their similarity was evaluated with FishBase-estimated TP values. For the SIA method, TP estimated with (a) only δ15N and with (b) both isotopes (δ15N and δ13C). For the SCA method, TP was estimated with Pauly’s TROPH using (a) default TP values of prey items and (b) those corrected when known by our SCA results.
Few studies exist that compare TP values calculated experimentally using different versions of both methods. Similar studies have found comparable results in some cases (Kline and Pauly 1998; Rybczynski et al. 2008), while in other cases, seasonal and method-dependent relations were found (Carscallen et al. 2012; Mancinelli et al. 2013). However, no attempts have been made to compare TP values obtained with two SIA methods, in which δ13C information is incorporated into one method, or with SCA methods where the TP of the prey items is modified to the default TROPH value.
Materials and methods
Sample collection
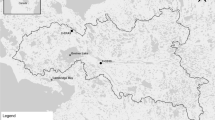
Fish specimens were collected in the Santa María la Reforma estuarine system, located on the continental shelf of the central Mexican Pacific. This is a type IIIA, inner-shelf coastal lagoon (Lankford 1977) populated by mangroves. Fish surveys were performed for five consecutive days at monthly intervals from December 2006 to March 2007 (winter season) at 29 stations (Fig. 1). Sampling was conducted onboard skiffs fitted with 115-HP outboard engines, equipped with the following fishing gear: (a) shrimp trawl net, fitted with a 24-m footrope and a 50-mm liner at the codend; (b) gillnet, 300 m long and fitted with a 75-mm liner; and (c) suripera net, which is a cast net modified for trawling fitted with a 3.5-cm liner and towed using the force of the wind or of the tide-generated current. A more detailed sampling protocol is available in Amezcua et al. (2006). Sampling hauls were limited to 10 min to minimize regurgitation or abnormal feeding. Specimens were immediately iced and then transported to the laboratory for analysis.

Studied area and sampling stations (crosses)
Additionally, we collected primary producers and consumers, detritus and seston from water column and sediments to establish the appropriate baseline for the estuarine food webs. Plankton samples were collected with 30 and 200 μm mesh conical nets at two knots for ~10 min for phytoplankton and zooplankton, respectively. Phytoplankton samples in the nets were cleansed with MilliQ water and filtered through GF/F filters with a 0.45-μm pore with the aid of a vacuum pump. Most of the zooplankton samples were processed and analyzed whole (as one category), and only a few samples were separated. Macroalgae, mangrove, saltwort, and cattail were also collected by hand with replicates (usually 15–20 of the second youngest leaves from 3 to 5 plants). Macroalgae and angiosperm samples were first rinsed with potable water to remove sediments and salt particles and then rinsed with MilliQ water. In each site and event, triplicated superficial water samples were collected in HCl-cleaned polyethylene 2 L bottles and transported to the laboratory for analysis. Suspended particulate matter (SPM), separated by filtering 500–2000 mL from water samples through a precombusted (500 °C, 4 h) glass fiber filter (GF/F), was determined by weight differences (before and after filtration) and was considered as ‘seston’ that comprises the living or dead phytoplankton and plant in disintegration. Surficial sediments were also collected in triplicate in each site by using a Van Veen bottom drag. Upper layers (few millimeters) were re-sampled from surface sediments. Polychaetes and other macrofaunal organisms were separated from sediment samples obtained with a 0.1-m2 grab and sieved through a 0.5-mm mesh. The organic matter in surface sediments considered as detritus, benthic microalgae and meiofauna (<0.5 mm) was not separated but analyzed as a whole category, detritus. All samples were packed in sealed doubled plastic bags and kept on ice immediately after collection until they were processed in the laboratory.
Sample preparation and analysis
Fish were identified to species level, with total length (TL) and total weight (TW) recorded for each specimen. Stomach contents were examined using a stereoscopic microscope. Prey items were counted, weighed, and identified to the lowest taxonomic level possible. Gravimetric measurements reflect dietary nutritional value (Macdonald and Green 1983), so stomach content results were expressed as gravimetric percentages, calculated for each item consumed by a fish species based on total stomach content mass (Hyslop 1980). A randomized cumulative curve was created plotting new prey types against the number of nonempty stomachs to determine whether the number of analyzed organisms was sufficient to describe the diet of each species (Ferry and Caillet 1996). If not, that species was removed from the analysis. In order to avoid ontogenetic diet changes, only species with enough adults for a meaningful SCA and SIA were selected. From the selected species, a subsample of 5–10 specimens of the same species and size range and collected in the same habitat (lagoonal, transitional and marine) and season (dry and rainy) were rinsed in Milli-Q water and stored frozen at −20 °C in a double-ziplock bag until dissection.
A total of 297 samples of fish were prepared for SIA. In order to minimize the effects associated with depleted δ13C values, we collected low-lipid dorsal muscle tissue from each selected specimen (Bodin et al. 2007). All samples were lyophilized (−44 °C, 33–72 mmHg, 72 h) and pulverized to a homogeneous powder using an agate mortar. Aliquots of homogenized fish muscle were weighed (1.0 ± 0.1 mg) with a microbalance, packed in tin cups (5 × 9 mm) and placed into sample trays until analysis (Costech, Valencia, CA) for carbon and nitrogen contents and their isotopes. Before carbon content and stable isotope analyses, samples were exposed to HCl vapor for 4 h (acid-fuming) to remove carbonates and then dried at 60 °C for 6 h (Harris et al. 2001).
All biota (primary producers and consumers), seston and detritus samples were stored frozen at −20 °C, lyophilized at −45 °C for 3 days and pulverized to a homogeneous powder in an agate mortar. The samples were then transferred to plastic containers and stored until analysis. Samples were treated with acid prior to isotopic analysis (HCl vapors for 4 h within a glass desiccator). Although only small aliquots (~0.2 mg of samples on a dry weight basis) are required for analyzing bulk carbon and nitrogen isotopes, planktonic samples are nonetheless difficult to compile. Therefore, in many cases, several individuals of the same or different species at a given site were combined to have sufficient biomass for SIA. Aliquots were weighed, pressed into tin capsules and sent to the Stable Isotope Facility at the University of California in Davis for determination of stable isotope ratios (13C/12C and 15N/14N). Analyses of stable isotope composition used a PDZ Europa ANCA-GSL elemental analyzer interfaced to a PDZ Europa 20-20 isotope ratio mass spectrometer (Sercon Ltd., Cheshire, UK). Isotope ratios of the samples were calculated using the equation δ (‰) = (R sample/R standard−1)] × 1000, where R = 15N/14N or 13C/12C. The R standard is relative to international standards, the Air and V-PDB (Vienna PeeDee Belemnite) for N and C, respectively. The analytical precision of these measurements was 0.2 % for δ13C and 0.3 % for δ15N (http://stableisotopefacility.ucdavis.edu/13cand15n.html).
Estimation of trophic position and associated standard errors
TP values based on SCA and their associated standard errors were estimated using a stand-alone quantitative application in TrophLab (Pauly et al. 2000a). TP was calculated by adding 1 to the mean trophic position and weighted by the relative abundance of all food items consumed by a species. Food items are assigned to discrete trophic levels, with basal resources and detritus located at a definitional trophic level of 1, following the convention established in the 1960s by the International Biological Program (Froese and Pauly 2015). A trophic position (TROPH) value is thus obtained and expresses an organism’s position in the food web. The TROPH of fish species i was estimated from the following equation:
where DC ij is the fraction of prey j and is the diet of consumer i, TROPH j is the trophic level of prey j, and G is the number of groups in the diet of i. The standard error (SE) of the TROPH was estimated using the weight contribution and the trophic level of each prey species in the stomach. TrophLab uses default TP values for various prey items (FishBase; Froese and Pauly 2015), but if known, the TP values can be replaced. In this sense, two separate SCA values for each predator species were determined—the first using the default TP value of the prey items (SCAD) and the second using the known trophic position value of fish prey items based on our SCA results (SCAC).
TP estimates based on SIA were also determined in two ways. First, TP was determined using only the N isotopic ratios of the consumer species (SIAN):
where λ is the trophic position of the food web base, δ15Nfish is the nitrogen signature of the species of fish being evaluated and F is the isotopic discrimination factor.
Selection of the most ecologically meaningful values of Eq. (2) parameters (TP in the base, δ15N in the baseline indicator and the enrichment factor) is a key step for TP calculations when applying SIA to the study of food webs (Post 2002; Layman et al. 2012). Here, we chose primary consumers as baseline indicators (λ = 2) to calculate the food web base because they showed a lower spatial–temporal variability compared to primary producers. In order to provide a more general and accurate baseline, we included zooplankton and benthic primary consumers collected in different habitats across the estuarine system and climatic seasons (e.g., rainy and dry). During the study, the grand mean δ15N of primary consumers was 8.84 ± 0.39 ‰, with interspecific variability in isotopic signatures considerably lower than that observed for basal resources such as primary producers, seston and detritus (data not included), ranging from a minimum of 8.46 ± 1.28 ‰ in zooplankton dominated by copepods to 9.39 ± 0.78 ‰ for fish larvae. In our study, the universal reference value of 3.4 ‰ was used as the trophic discrimination factor (Minagawa and Wada 1984). However, we recognized that F values might vary among fish species and among individuals of a given species according to nutritional, environment and individual conditions (Bojorquez-Mascareño and Soto-Jiménez in press) but knowledge on how the trophic discrimination vary is limited (Martínez del Rio et al. 2009).
Because primary consumers were collected from a two-source food web (e.g., benthic and pelagic food web), each with a separate set of primary producers and detritus sources, we used the following equation (Post 2002) to recalculate the trophic position SIANC for all fish species, taking into account the proportion N in the consumer ultimately derived from the base of food web 1 (α):
where δ15Nfish is the N signal in fish species, and δ15Nbase1 and δ15Nbase2 are the mean δ15N signatures of food web 1 (pelagic) and 2 (benthic) baselines, respectively. Finally, the constant α was estimated following a two-endmember-mixing model that distinguishes the both sources of carbon or energy:
The model in Eq. 4 assumes that (1) movement of N and C through both food webs is similar, (2) there is little or no trophic fractionation of C and (3) the mixing process is linear (Post, 2002). In our study, the constant α was estimated in 0.67 considering two complex mixtures of carbon sources, the pelagic that consisted of phytoplankton and seston (δ15N = 7.2–8.0 ‰ and δ13C = −24.7 to −19.78 ‰) and the benthic that consisted of live macroalgae and detritus of angiosperms and macroalgae (δ15N = 6.5–9.1 ‰ and δ13C = −21.22 to −28.23 ‰).
Statistical analyses
Trophic position values calculated with different methods were compared, and their relationships with FishBase estimates were tested. A two-way ANOVA was performed with the TP value as the dependent variable and the method (SCADd, SCACc, SIANC and SIAN) and species as fixed categorical factors or independent variables. This statistical arrangement is useful in determining differences in TP values according to methods, species and the interaction method/species. If significant differences were detected, multiple comparisons of means were performed using the Tukey’s HSD test. The homogeneity of variances was tested using Cochran’s C test, and the data were transformed when necessary.
Histograms of FishBase values and TP values estimated with each method were plotted as separate normal distributions with their corresponding means and standard deviations, in order to determine which method better characterized the trophic complexity and feeding guilds of the fish assemblage. Use of this procedure with TP values has been used previously successful for the identification of functional trophic groups (Stergiou and Karpouzi 2002).
Corresponding means and standard deviations (SD) of each TP of the normal distributions were calculated. Estimation of these parameters for each normal distribution was achieved using maximum likelihood criterion in order to obtain a probability value for each of the k possible outcomes (pk) in the n trials. To obtain the expected number of cases, the log-likelihoods from the following multinomial equation were used:
where LL{L | μ n , σ n } is the likelihood of any individual observation X, given μ (the population mean) and σ (the population standard deviation). μ n and σ n are the mean and standard deviations of the n cohorts considered. There are k length classes, and L i is the observed frequency of length class i, while p i -hat is the expected proportion of length class i from the combined normal distributions. A detailed explanation of this method can be found in Haddon (2001).
Once the normal distributions or modes for each method were identified, the multivariate multiple permutations test SIMPER (similarity percentages) was used to determine the main prey items that corresponded to the suite of fish species present in every mode for each method and therefore identify functional trophic groups for each of the methods used to estimate TP. To do this, a matrix containing fish species as samples (columns) and the prey items as variables (rows) was constructed, with the proportion of each prey item in the diet of each fish as the data. Four factors were then assigned to each fish species (SCADd, SCACc, SIANC and SIAN), which were used to estimate TP, and the factor values were the means obtained for each mode of every method with the maximum likelihood procedure. Then, the average similarity of each prey item within every group (normal distribution) was estimated with Euclidean distances. The analysis was performed with the analytical software PRIMER 6 (Clarke and Warwick 1994). In the case of the normal distributions obtained from the FishBase data, prey items for every mode were obtained directly from web site information.
Results
A total of 8992 adult specimens belonging to 66 fish species and 33 families were analyzed for the estimation of TP values with SCA and SIA and compared to those provided from FishBase (Table 1). The mean TL of adults ranged from 8.2 cm for a Pacific moonfish to 53.8 cm for a long-tail stingray. The cumulative prey item curve fit better with a logistic curve (R 2 > 0.94, P < 0.05) than that with a linear relationship (R 2 < 0.93, P > 0.05) for all 66 species; therefore, sample size was considered to be sufficient for describing the diet of all 66 species. The identified prey items included plankton, nekton, hyperbenthos and epibenthos.
Estimated TPSCAD values for stomach content data ranged from 2.65 for the big-scale anchovy to 4.5 for the Mexican barracuda and the Pacific cornetfish, while estimated TPSCAC values ranged from 2.65 for the big-scale anchovy to 5.21 for the toothed flounder. \({\text{TP}}_{{{\text{SIA}}_{\text{N}} }}\) values ranged from 2.74 (Pacific spadefish) to 4.96 (cominate sea catfish), while the \({\text{TP}}_{{{\text{SIA}}_{\text{NC}} }}\) values ranged from 2.99 to 5.21 for the same two species. Both SIA methods and the \({\text{TP}}_{{{\text{SCA}}_{\text{C}} }}\) identified a maximum trophic value of 5.21, but for different species.
Significant differences were found between the mean TP value of species (F (65,1057) = 40.235, p < 0.01), according to the method used (F (3,1057) = 794.15, p < 0.01), and the interaction species/method (F (195,1057) = 9.4215, p < 0.01). Based on the ANOVA results, a plot of fish species versus TP values determined by different methods was constructed, with the data plotted according to the TP values estimated with SIANC. TP values estimated by SIA methods did not differ significantly (Tukey’s HSD test), and in general, these values were higher than those obtained by SCA methods and reported by FishBase. TP values obtained by SCA methods were typically similar, but in 10 species the SCAC was significantly higher than the SCAD. Only for 5 of 10 species did the SCAC value not differ from the SIA values, while in the other 5 species, the SCAC values were significantly higher than both SIA values (Tukey’s HSD test; Fig. 2). Only 6 species had TP values that did not differ statistically among methods used—all 6 had medium-to-low TP values. For 4 species, both TP values estimated with SCA were significantly higher than those estimated with SIA, and all corresponded to species of lower TP according to SIA (left part of the graph). In general, in this part of the graph, the highest TP values estimated with the TROPH routine are found, including values from FishBase. The only 2 exceptions are the toothed and Panamic flounders, in which FishBase shows a low TP value. The TP value estimated with SCAD for these species is not different from those estimated with SIA, but the values estimated with SCAC are significantly higher. In fact, according to TROPH, these 2 species are the top predators in the studied ecosystem. For the other 50 species, the TP values estimated with SIA were significantly higher than those estimated with SCA.

Comparison of the mean TP of each fish species as estimated by the five methods. \({\text{TP}}_{{{\text{SIA}}_{\text{NC}} }}\): trophic position estimated with stable isotopes of nitrogen and carbon. \({\text{TP}}_{{{\text{SIA}}_{\text{N}} }}\): trophic position estimated with stable isotopes of nitrogen. \({\text{TP}}_{{{\text{SCA}}_{\text{C}} }}\): trophic position estimated with stomach content analysis using the known TP of the fish preys. \({\text{TP}}_{{{\text{SCA}}_{\text{D}} }}\): trophic position estimated with stomach content analysis using the default TP value of preys given by TROPH. TPFishBase: trophic position obtained from FishBase. Circles indicated species where significant differences were not found between TPSIA and TPSCA values. Arrows indicated species whose TPSCA value was significantly higher than the TPSIA value. Lined arrows indicated species whose \({\text{TP}}_{{{\text{SCA}}_{\text{C}} }}\) value was significantly higher than the \({\text{TP}}_{{{\text{SCA}}_{\text{D}} }}\) value
Both SIA methods showed five different modes when the histograms of the estimated TP values were decomposed into separate normal distributions. Both histograms showed similar patterns, with most data clustered toward the right of the graph, with TP values over 4.00, although the means estimated with \({\text{TP}}_{{{\text{SIA}}_{\text{NC}} }}\) were higher (3.1, 3.70, 4.36, 4.73 and 5.01) (Fig. 3a) than those obtained with \({\text{TP}}_{{{\text{SIA}}_{\text{N}} }}\) (2.91, 3.47, 4.12, 4.49 and 4.78) (Fig. 3b). The estimated \({\text{TP}}_{{{\text{SCA}}_{\text{C}} }}\) values displayed four normal distribution components with three main modes with TP means at 3.22, 3.68 and 4.15, and one minor mode with mean TP of 5.25. This pattern differed from that of the SIA method, where most of the values were under 3.8 (Fig. 3c). A similar pattern was observed between \({\text{TP}}_{{{\text{SCA}}_{\text{C}} }}\) and \({\text{TP}}_{{{\text{SCA}}_{\text{D}} }}\) methods, but with the fourth mode from the \({\text{TP}}_{{{\text{SCA}}_{\text{D}} }}\) method integrated to third and the last mode of \({\text{TP}}_{{{\text{SCA}}_{\text{C}} }}\) method (Fig. 3d). Finally, TROPH values obtained from FishBase displayed four normal distribution components, with two main modes at mean TP values of 3.49 and 4.00, and two minor modes with mean TP values of 2.00 and 3.14. The pattern was similar to that of the \({\text{TP}}_{{{\text{SCA}}_{\text{D}} }}\) method; however, FishBase had one species with a very low TP value (two for the striped mullet), and therefore, the mode appears left-shifted on the graph (Fig. 3e).

Histograms and decomposed modes of TP values estimated by each method. The organisms at the top of each mode denote the most frequently consumed prey item for that normal distribution according to SIMPER. The numbers underneath indicate the mean TP for that mode and standard deviations (in parentheses)
SIMPER analysis allowed us to assign functional trophic groups to every mode observed. For both SIA methods, the first modes included the bentho-pelagic, reef-associated spadefish and the demersal Peruvian mojarra, first-order carnivores that feed on benthic invertebrates and hydroids. The second group included the pelagic cornet fish and demersal small flatfish, omnivores with a preference for small benthic fauna and juvenile and larvae fish. The third functional group included pelagic, bentho-pelagic and demersal species, such as anchovies, mojarras, croakers, rays and puffers, and consisted of omnivorous fish with a preference for demersal fish, macrobenthos and infaunal crustaceans. The fourth group included pelagic, bentho-pelagic and demersal species, such as drums, grunts, corvinas, weakfish and morays, and consisted of omnivorous fish with a preference for macrobenthos and demersal fish species. The fifth group, which also included demersal, bentho-pelagic and pelagic species such as perch, grunts, croakers, snappers and the Pacific sierra, was comprised of carnivorous fish that consumed fish and benthos with similar high TP values (Fig. 3a, b).
For the \({\text{TP}}_{{{\text{SCA}}_{\text{C}} }}\) values, the first functional trophic group included pelagic and demersal species such as anchovies, small flatfish and mojarras, and consisted of second-order carnivores that fed on benthic organisms and planktivorous crustaceans. The second group, which included pelagic, bentho-pelagic and demersal species such as rays, mojarras, mullets, grunts, croakers, tonguefish, catfish, herring, eels and guitarfish, was comprised of omnivorous fish with a preference for decapod crustaceans. The third functional group included demersal and pelagic species such as weakfish, corvina, snappers and the Pacific sierra, and consisted of omnivorous fish with a preference for fish, squid and shrimp. The fourth group included demersal large flatfish (flounders) was comprised of omnivores with a preference for fish (Fig. 3c).
For the \({\text{TP}}_{{{\text{SCA}}_{\text{D}} }}\) values, the first functional trophic group, which included pelagic fish such as anchovies and herrings, consisted of second-order carnivores feeding on planktivorous crustaceans. The second group consisted of omnivorous fish with a preference for decapod crustaceans, demersal fish and mollusks, and included most of the fish species analyzed from pelagic, bentho-pelagic and demersal habitats. The third functional group, which included bentho-pelagic and demersal fish such as snappers, large flatfish (flounders), barracuda, snooks and cornet fish, consisted of omnivorous fish, carnivores that consume only fish and decapods (Fig. 3d).
Finally, for the FishBase TP values, the prey items in each mode were assigned according to the most common items from the suite of fish species in each normal distribution, based on information provided by this database. The first mode includes only the striped mullet, which feeds on algae and detritus according to FishBase (Froese and Pauly 2015). The second mode consists of mostly demersal species and the Pacific spadefish, which are second-order predators that feed on plankton and small benthic invertebrates. The third mode includes a majority of the species, with inhabitants of all habitats that prey on macrobenthos and small fish. Finally, the fourth mode includes demersal, pelagic and bentho-pelagic fish and includes the top predators of the system according to FishBase. These were omnivores or carnivores feeding mainly on fish, squid and decapoda.
Discussion and conclusions
Accurately estimating the TP of fish is essential for ecosystem-based fishery management (EBFM), as it has been recognized that the classical approach to single-species management is not adequate and that all exploited species should be considered, even when not the target species (Clark et al. 2001; Marasco et al. 2007). Shrimp trawl fishing is the most important industry in the Gulf of California’s estuarine, coastal and open sea regions. Previous studies have shown the industry to be the main generator of bycatch, which is primarily composed of fish primary harvested by small-scale fisheries (Amezcua et al. 2006, Madrid-Vera et al. 2007; Amezcua et al. 2009). Although the Gulf’s fishing is a small-scale industry, it supports thousands of local families. At present, adequate management for most demersal and bentho-pelagic fish species in the Mexican Pacific is nonexistent, as is the case for similar species in tropical and subtropical regions as well. The Mexican government recognizes its need for an EBFM (CONAPESCA 2010), for which knowledge of fish TPs is essential, as it enables estimation of the “Primary Production Required” to support fisheries in any locality, as well as evaluation of ‘fishing down the food web.’
Fish TP values are currently obtained with the following methods: (1) from the FishBase information; (2) from SCA and then applied a routine such as Pauly’s TROPH; and (3) from SIA. To the best of our knowledge, this is the first study that compares values obtained from these three methods and its variations, for finfish of subtropical coastal lagoons, and determines whether they could potentially be used interchangeably. Because the SCA and SIA each have variations that could yield different results, we calculated five TP values for every fish species: the FishBase value, a SCAD value obtained using the default TP values of prey items in TrophLab, a SCAC value also obtained with TrophLab but with modified TP values for fish preys, a SIAN value obtained with δ15N isotopes, and a SIANC value obtained with a combination of δ15N and δ13C isotopes.
The ANOVA results indicate that the fish included in this study come from the top part of the food chain, as most TP values were above 3 with all the methods employed. Although about half of fish had TP values around 4, distribution was scattered between the values of 3 and 5. The estuarine system studied has approximately 200 fish species (Amezcua et al. 2006); however, the 66 species used in this study represent ≅70 % of the abundance of those species. Although not our study goal, we suspect that the system could be dominated TP four fish, with few top predators, and with invertebrates as the first-order predators. However, we do not have sufficient evidence to support this supposition.
The ANOVA results indicated differences between methods, and although the FishBase TP values could not be included in our analyses, when graphed against the other results, some tendencies are evident. In general, the FishBase values were similar to those obtained in our study, as both were obtained from the same method (TrophLab). However, for about 12 species, the results were not alike. A reason for this is that FishBase uses values obtained from all over the world and spatial differences could cause these dissimilarities. Further, we only used adult fish, while FishBase does not specify fish maturity. Also, FishBase reports some trophic levels estimated on diet items known to be ingested by a given species using a randomized resampling routine, when the proportion of each prey item in the diet is not known. All these circumstances can cause our results to be different from those given by FishBase, even if these are estimated with the same method.
When TrophLab values are compared with both SIA methods, the SIA values are higher for 49 species, the same for 6 species and lower than SCA values for 4 species. In 10 cases, correcting the TP value of the prey caused the SCAC value to be higher than the SCAD value. The two SIA methods do not differ, but the values obtained with SIANC were always higher than those obtained with SIAN. Considering this, it is evident that the tree methods of determining TP fish values do not yield similar results, and it is necessary to carefully choose the best method.
The plot in Fig. 2 indicates that TrophLab results underestimate TP values. SCAC values further support this claim, as TP values increased when the TP values of fish preys were known and could be replace the default value given by TrophLab. In this work, we only could replace values of fish that appeared as prey items of other fish. However, the TP values of all invertebrates were not be greatly modified. When a diet was primarily contained fish, the TP SCAC value was significantly higher than the SCAD value. In this case, the TP SCAC values were generally similar to those obtained with both SIA methods. The maximum value yielded by TrophLab using default TROPH values for all prey items is 4.5, a TP that corresponds to top predators (Stergiou and Karpouzi 2002). TrophLab assumes that all the prey items of a taxon have the same TROPH value, which is not accurate. For example, the default TROPH value for any fish is 3.5, but it is known that fish with larger TROPH values can also be prey items of other fish species. This occurs with other taxa as well; for example, the default TROPH value of any gastropod is 2.37, a value that was likely estimated according to the fact that these organisms mainly feed on algae and detritus. However, during the course of our study, carnivorous gastropods from the family Muricidae were found in the stomachs of other fish. These gastropods likely have a TP greater than 2.37; if the trophic level of each prey was known and replaced in TrophLab, the results were likely be more similar.
Both SCA values were higher than the SIA values in cornet fish, spade fish, barracuda and a snook. For the two fish, the higher values might be attributed to the fact that both species eat fish larvae that were not possible to identify. TrophLab assigned a TP of 3.5, as if the fish were large, resulting in overestimated values. A similar scenario may have occurred with the barracuda, the snook and the two flounders, which had SCAC values higher than the SIA values. The diet of these species consists of large quantities of fish. Perhaps, SCAC values are overestimated.
When observing at the multimodal analyses and different functional trophic groups, SCA seems to oversimplify the trophic chain. The TP–SIA frequencies tended toward the left, which is logical, considering that higher TP values were obtained by these methods. Both methods showed five modes or groups that could be translated into trophic levels. Thus, according to the TP–SIA, this system has six trophic levels when considering the primary consumers and assuming that remaining fish and invertebrates are included in these modes. For both SIA methods, the pattern was similar, and the functional trophic groups identified with SIMPER shift from hydroid-consuming omnivores, and infaunal crustaceans in the lower levels to mere carnivores in the top levels. Conversely, the modes identified with all 3 TrophLab-base methods indicate that most of the fish species have TP values around 3.6. The SCAD and FishBase values show similar patterns. However, FishBase values show a small left-shifted mode, because it includes the mullet as an herbivore, whereas the SCAD values show a right-shifted mode, because the corrected values were higher than the initial values. Nonetheless, the trophic structure based on TrophLab methods is less complex than that observed with either SIA method and did not include carnivores in its top trophic positions. Further, we observed no a dietary pattern, unlike in the functional trophic groups found with both SIA methods, in which the fish with lower TP values shifted from hydroids and infernal crustaceans to diets including fish larvae, fish and other macrobenthos, to mere carnivores. We concluded that stable isotope analysis yielded a more accurate community structure than the SCA. This finding is of importance because identified trophic groups that are identified may serve as the basis for the maintenance of trophic position balance in estuarine systems when evaluating corresponding indicators (e.g., aggregate annual removal from each trophic group) and reference points (e.g., maximum percentage removal from each trophic group). If the groups are not accurately estimated, proper management is not possible, as is the case when the TP–SCA groups are used.
Each of these two methods has its advantages and disadvantages (Polunin and Pinnegar 2000; Stergiou and Polunin 2000). The SCA is easy to apply and provides an instantaneous record of an animal’s food type and quantity (Costa et al. 1992; Elliott and Hemingway 2002). SCA also provides an initial view of the ichthyological trophic structure of the system by describing the relationships between a fish species and its prey. However, it relies on various assumptions such as the proper identification and quantification of all food items, must to take the differential digestion rates of prey species into account (Polunin and Pinnegar 2000; Stergiou and Polunin 2000). This alone is a critical issue, as organism identification proves difficult, such as gelatinous zooplankton and detritus (Polunin and Pinnegar 2000), as well as highly digestible invertebrates and fish. In addition, carnivores frequently have empty stomachs (e.g., Beyer 1998), and the SCA offers mere ‘snapshots’ of diet, rendering impossible the identification of the origin of the organic matter source (Pasquaud et al. 2007). Most importantly, the TrophLab method makes oversimplified assumptions, such as the fact that all potential prey in one taxon has similar TROPH values, when in fact, prey TP values are often unknown (Pinnegar and Polunin 1999, 2000; Polunin and Pinnegar 2000).
The SIA method has been proven useful in many studies (Post et al. 2000; Vander Zanden et al. 1999). But also have its advantages and disadvantages (Polunin and Pinnegar 2000). With omnivorous fish, SIA is preferable to SCA, as it provides an integrated signal of what the fish has assimilated and positions multiples species along each other. In addition, SIA uses energy transfer information to position species in food webs. However, in environments including estuaries, SIA does not allow precise identification and quantification of complex mixtures of organic matter at the base of fish food webs. Furthermore, SIA does not proportionalize the prey items consumed, which makes it difficult to establish trophic interactions. In the Gulf of California, a system characterized by productive and complex food webs (Vega-Cendejas and Arreguín-Sánchez 2001; Zetina-Rejón et al. 2003), as well as changing temporal and spatial environmental conditions (Lankford 1977), there are a large variety of autochthons and allochthonous organic carbon sources. The trophic contributions to multiple carbon sources in this ecosystem play a crucial role in maintaining high biotic diversity and productivity, making it extremely important to quantify the role of each carbon source in the nutrition supporting food webs of these coastal ecosystems. In this regard, the δ13C isotopic analyses provide information useful for elucidating food origins and energy flow through estuarine food webs—task that are not possible with only SCA. For example, in this study, most fish had δ13C values related to their potential food sources. The pelagic species primarily related to phytoplankton, and the bentho-pelagic and demersal species primarily related to macrophyte detritus and benthic microalgae.
Previous studies have shown that the combined use of stomach content analysis and nitrogen stable isotope analysis can provide a detailed picture of the structure of an estuarine fish food web. They have recommended the use of both these methods to gain a better understanding of estuarine systems (e.g., TrophLab: Christensen and Pauly 1992; Pauly et al. 2000a; δ15N values: Kline and Pauly 1998; Polunin and Pinnegar 2000). Thus, a cross-validation using different methods is essential. However, our study has proven that use of only SCA and TrophLab for estimating the fish TP values is not adequate. We contend that \({\text{TP}}_{{{\text{SIA}}_{\text{NC}} }}\) that combines both isotopes yields the most accurate results, providing a common currency for large-scale, cross-system comparisons of trophic structures and energy flow in this ecosystem characterized by two food web bases. It also consumes less time and money, as diet can be adequately described by 500–1000 stomachs, depending on whether organisms are food specialists or food generalists (Link and Almeida 2000). However, if stomach content studies are used to estimate fish TP values, seasonal samples (rather than circumstantial samples) must be collected to estimate an overall feeding pattern. Further, it is necessary to correct FishBase TP values of prey.
References
Amezcua F, Madrid-Vera J, Aguirre-Villaseñor H (2006) Effect of the artisanal shrimp fishery on the ichthyofauna in the coastal lagoon of Santa Maria la Reforma, Gulf of California. Cienc Mar 32:97–109
Amezcua F, Madrid-Vera J, Aguirre H (2009) Incidental capture of juvenile fish from an artisanal fishery in a coastal lagoon in the Gulf of California. North Am J Fish Manag 29:245–255
Baird D, Ulanowicz RE (1993) Comparative study on the trophic structure, cycling and ecosystem properties of four tidal estuaries. Mar Ecol Prog Ser 99:221–237
Baird D, McGlade JM, Ulanowicz RE (1991) The comparative ecology of 6 marine ecosystems. Philos Trans R Soc Lond B Biol Sci 333:15–29
Beyer JE (1998) Stochastic stomach theory of fish: an introduction. Ecol Model 114:71–93
Bodin N, Le Loc’h F, Hily C (2007) Effect of lipid removal on carbon and nitrogen stable isotope ratios in crustacean tissues. J Exp Mar Biol Ecol 341:168–175
Bojorquez-Mascareño EI, Soto-Jiménez MF (in press) Isotopic turnover rate and trophic fractionation of nitrogen in shrimp Litopenaeus vannamei (Boone) by experimental mesocosms: implications for the estimation of the relative contribution of diets. Aquac Res doi:10.1111/are.12757
Bondavalli C, Bodini A, Rossetti G, Allesina S (2006) Detecting stress at a whole ecosystem level: the case of a mountain lake (Lake Santo, Italy). Ecosystems 9:1–56
Branch TA, Watson R, Fulton EA, Jennings S, McGilliard CR, Pablico GT, Ricard D, Tracey SR (2010) The trophic fingerprint of marine fisheries. Nature 468:431–435
Cabana G, Rasmussen JB (1996) Comparison of aquatic food chains using nitrogen isotopes. Proc Natl Acad Sci 93:10844–10847
Carscallen WMA, Vandenberg K, Lawson JM, Martinez ND, Romanuk TN (2012) Estimating trophic position in marine and estuarine food webs. Ecosphere 3(3):art25
Christensen V, Pauly D (1992) ECOPATH II—a software for balancing steady-state ecosystem models and calculating network characteristics. Ecol Model 61:169–185
Clark JS, Carpenter SR, Barber M, Collins S, Dobson A, Foley JA, Lodge DM, Pascual M, Pielke R, Pizer W, Pringle C, Reid W, Rose KA, Sala O, Schlesinger WH, Wall DH, Wear D (2001) Ecological forecasts: an emerging imperative. Science 293:657–660
Clarke KR, Warwick RM (1994) Change in marine communities: an approach to statistical analysis and interpretation. PRIMER, Plymouth
Codron D, Sponheimer M, Codron J, Newton I, Lanham J, Clauss M (2012) The confounding effects of source isotopic heterogeneity on consumer-diet and tissue-tissue stable isotope relationships. Oecologia 169:939–953
CONAPESCA (2010) Políticas de Ordenamiento para la Pesca y Acuacultura Sustentables, en el marco de Programa Rector de Pesca y Acuacultura. Comisión Nacional de Acuacultura y Pesca, Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación, México (In Spanish)
Costa JL, Assis CA, Almeida PR, Moreira FM, Costa MJ (1992) On the food of the European eel, Anguilla anguilla (L.), in the upper zone of the Tagus estuary, Portugal.J Fish Biol 41:841–850
Deegan LA, Garritt RH (1997) Evidence for spatial variability in estuarine food webs. Mar Ecol Prog Ser 147:31–47
DeNiro MJ, Epstein S (1981) Influence of the diet on the distribution of nitrogen isotopes in animals. Geochim Cosmochim Acta 45:341351
Elliott M, Hemingway K (2002) Fishes in estuaries. Blackwells, London
Ferry LA, Caillet GM (1996) Sample size and data analysis: are we characterizing and comparing diet properly? In: MacKinlay D, Shearer K (eds) Proceedings of the symposium on the feeding ecology and nutrition in fish. American Fisheries Society, San Francisco, pp 71–80
Froese R, Pauly D (2015) FishBase. http://fishbase.org. Accessed 02 Feb 2015
Haddon M (2001) Modelling and quantitative methods in fisheries. CRC Press, Boca Raton
Harris D, Horwath R, Van Kessel C (2001) Acid fumigation of soils to remove carbonates prior to total organic carbon or carbon-13 isotopic analysis. Soil Sci Soc Am J 65:1853–1856
Hyslop EJ (1980) Stomach contents analysis e a review of methods and their application. J Fish Biol 17:411–429
Kline TC, Pauly D (1998) Cross-validation of trophic level estimates from a mass balance model of Prince William sound using 15N/14N data. In: Quinn TJ II, Funk F, Heifetz J, Ianelli JN, Powers JE, Schweigert JF, Sullivan PJ, Zhang CI (eds) Fishery stock assessment models. Alaska Sea Grant, Fairbanks, pp 693–702
Koen-Alonso M (2007) A process-oriented approach to the multispecies functional response. In: Rooney N, McCann KS, Noakes DLG (eds) From energetics to ecosystems: the dynamics and structure of ecological systems. Springer, Netherlands, pp 1–36
Lankford RR (1977) Coastal lagoons of Mexico, their origin and classification. Estuar Processes 2:182–215
Layman CA, Araujo MS, Boucek R, Hammerschlag-Peyer CM, Harrison E, Jud ZR, Matich P, Rosenblatt AE, Vaudo JJ, Yeager LA, Post DM, Bearhop S (2012) Applying stable isotopes to examine food-web structure: an overview of analytical tools. Biol Rev 7:545–562
Link JS, Almeida FP (2000) An overview and history of the food web dynamics program of the Northeast Fisheries Science Center, Woods Hole, Massachusetts. NOAA Technical Memorandum NMFS-NE-159
Macdonald JS, Green RH (1983) Redundancy of variables used to describe importance of prey species in fish diets. Can J Fish Aquat Sci 40:635–637
Madrid-Vera J, Amezcua F, Morales-Bojórquez E (2007) An assessment approach to estimate biomass of fish communities from bycatch data in a tropical shrimp-trawl fishery. Fish Res 83:81–89
Mancinelli G, Vizzini S, Mazzola A, Macia S, Basset A (2013) Cross-validation of δ15N and FishBase estimates of fish trophic position in a Mediterranean lagoon: the importance of the isotopic baseline. Estuar Coast Shelf Sci 135:77–85
Mangel M, Levin PS (2005) Regime, phase and paradigm shifts: making community ecology the basic science for fisheries. Philos Trans R Soc B Biol Sci 360:95–105
Marasco RJ, Goodman D, Grimes CB, Lawson PW, Punt AE, Quinn TJ II (2007) Ecosystem-based fisheries management: some practical suggestions. Can J Fish Aquat Sci 64:928–939
Martínez del Rio C, Sabat P, Anderson-Sprecher R, Gonzalez SP (2009) Dietary and isotopic specialization: the isotopic niche of three Cinclodes ovenbirds. Oecologia 161:149–159
Minagawa M, Wada E (1984) Stepwise enrichment of 15N along food chains: further evidence and the relation between δ15N and animal age. Geochim Cosmochim Acta 48:1135–1140
Pasquaud S, Lobry J, Elie P (2007) Facing the necessity of describing estuarine ecosystems: a review of food web ecology study techniques. Hydrobiologia 588:159–172
Pasquaud S, Pillet M, David V, Sautour B, Elie P (2010) Determination of fish trophic levels in an estuarine system. Estuar Coast Shelf Sci 86:237–246
Pauly D, Watson R (2005) Background and interpretation of the ‘Marine Trophic Index’ as a measure of biodiversity. Phil Trans R Soc B Biol Sci 360:415–423
Pauly D, Christensen V, Walters C (2000a) Ecopath, Ecosim, and Ecospace as tools for evaluating ecosystem impacts on marine ecosystems. ICES J Mar Sci 57:697–706
Pauly D, Froese R, Rius MJFD (2000b) Trophic pyramids. In: Froese R, Pauly D (eds) FishBase 2000: concepts, design and data sources. ICLARM, Manila, pp 203–204
Pinnegar JK, Polunin NVC (1999) Differential fractionation of δ13C and δ15N among fish tissues: implications for the study of trophic interactions. Funct Ecol 13:225–231
Pinnegar JK, Polunin NVC (2000) Contributions of stable isotope data to elucidating food webs of Mediterranean rocky littoral fishes. Oecologia 122:399–409
Polunin NVC, Pinnegar JK (2000) Trophic-level dynamics inferred from stable isotopes of carbon and nitrogen. In: Briand F (ed) Fishing down the Mediterranean food webs. CIESM workshop series, Kerkyra, pp 69–72
Post DM (2002) Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology 83:703–718
Post DM, Pace ML, Hairston NG (2000) Ecosystem size determines food-chain length in lakes. Nature 405:1047–1049
Quinn TJ II (2003) Ruminations on the development and future of population dynamics models in fisheries. Nat Resour Model 16(4):341–392
Rybczynski SM, Walters DM, Fritz KM, Johnson BR (2008) Comparing trophic position of stream fishes using stable isotope and gut contents analyses. Ecol Freshw Fish 17:199–206
Stergiou KI, Karpouzi VS (2002) Feeding habits and trophic levels of Mediterranean fish. Rev Fish Biol Fish. doi:10.1023/A:1020556722822
Stergiou KI, Polunin N (2000) Executive summary. In: Briand F (ed) Fishing down the Mediterranean food webs. CIESM workshop series, Kerkyra, pp 7–15
Ulanowicz R (1996) Trophic flow networks as indicators of ecosystem stress. In: Polis GA, Winemiller KO (eds) Food webs: integration of patterns and dynamics. Springer, Dordrecht, pp 358–370
Vander Zanden MJ, Cabana G, Rasmussen JB (1997) Comparing trophic position of freshwater fish calculated using stable nitrogen isotope ratios (δ15N) and literature dietary data. Can J Fish Aquat Sci 54:1142–1158
Vander Zanden MJ, Casselman JM, Rasmussen JB (1999) Stable isotope evidence for the food web consequences of species invasions in lakes. Nature 401:464–467
Vega-Cendejas ME, Arreguín-Sánchez F (2001) Energy fluxes in a mangrove ecosystem from a coastal lagoon in Yucatán Peninsula, Mexico. Ecol Model 137:119–133
Wada E, Mizutani H, Minagawa M (1991) The use of stable isotopes for food web analysis. Crit Rev Food Sci 30:361–371
Zetina-Rejón MJ, Arreguín-Sánchez F, Chávez EA (2003) Trophic structure and flows of energy in the Huizache-Caimanero lagoon complex on the Pacific coast of Mexico. Estuar Coast Shelf Sci 57:803–815
Acknowledgments
The authors thank all of the students who helped us with the SIA and SCA, and E. Jaffer for the English editing. We thank the editor and anonymous reviewers for their constructive comments, which helped us to improve the manuscript. This work was supported by the research project PAPIIT-IN217408-3 and IN208613.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Thomas Mehner.
Rights and permissions
About this article

Cite this article
Amezcua, F., Muro-Torres, V. & Soto-Jiménez, M.F. Stable isotope analysis versus TROPH: a comparison of methods for estimating fish trophic positions in a subtropical estuarine system. Aquat Ecol 49, 235–250 (2015). https://doi.org/10.1007/s10452-015-9517-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10452-015-9517-4