
Abstract
This study investigated the major environmental factors structuring, for a year, phytoplankton assemblages in the Sfax saltern (Tunisia): salinity and nutrients. A STATICO analysis based on 11 environmental variables and the abundances of 64 phytoplanktonic species was conducted. STATICO is used to analyze the stable part of the relationships between the environment and species, and then to determine how these relationships change over time. The analysis confirmed that the salinity gradient had a considerable influence on the composition of the phytoplanktonic communities. Bacillariophyceae and Dinophyceae dominated in the least salty ponds, whereas Chlorophyceae and Cyanophyceae dominated in the saltiest ponds, in accordance with the halotolerance level estimated for each species by calculating the optimum salinity and salt tolerance. Nitrogen (N) and phosphorus (P) ions could have a secondary influence on the phytoplankton distribution and its dynamics. Dinophyceae seem to be favored by high ammonium (NH4 +) concentrations, whereas diatoms seem to be favored by high orthophosphates (PO4 3−) and nitrates (NO3 −) values. The Chlorophyceae Dunalliela salina thrived in the saltiest ponds when the NO3 − concentrations increased.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Phytoplankton plays a critical role in aquatic ecosystems (Koffi et al. 2009) since it contributes to carbon fixation from the atmosphere and produces the organic matter needed for the food chains to function (Dufour and Durand 1982; Richardson et al. 1998). Phytoplankton is composed of dynamic multi-species assemblages characterized by high diversity and rapid successive shifts in species composition in response to environmental changes (Mendes et al. 2009). Identifying the factors and main processes governing the seasonal succession of species in the phytoplankton and the structure of communities is central problems in ecology (Anneville et al. 2004). The development of algal populations is regulated not only by physical and chemical factors (Richardson and Pedersen 1998), but also by biological factors such as predation (Elloumi et al. 2009; Maar et al. 2002) and competition between species (Mallin and Paerl 1994; Pinckney et al. 1998). However, the availability of nutrients remains the main factor controlling the composition and biomass of the phytoplankton community in shallow water and marshes (Ortega-Mayagoitia et al. 2003; Lopez-Flores et al. 2006). These abiotic parameters vary over time and affect the composition and abundance of the algal flora both directly and indirectly (Koffi et al. 2009).
Salt marshes are paralic ecosystems with specific characteristics. Salt ecosystems are home to a multitude of permanent-resident or migrant birds (Ramírez et al. 2011) and provide ideal nesting places since they are favorable to the development of microorganisms and invertebrates (Britton and Johnson 1987; Trigui et al. 2011). Indeed, evaporation is an important factor in salty coastal ecosystems, where it concentrates the nutritional elements (Wen and Zhi-Hui 1999; Dolapsakis et al. 2005; Oren 2009) that promote the proliferation of robust phytoplankton (Dolapsakis et al. 2005), and hence that of zooplankton (Vos and De la Rosa 1980). However, excessive benthic production leads to reduced salt quality (Davis and Giordano 1996).
Salt marshes are generally accessible and consist of successive ponds with differing and fairly stable ecological characteristics (Ventosa and Arahal 2009; Khemakhem et al. 2010; Madkour and Gaballah 2012). They are therefore suitable for studying the diversity of communities and how they are influenced by their environment (Oren 2009). Despite differences due to climatic changes, water fluctuations (Lopez-Flores et al. 2006), geographical location, human management practices, and the availability of nutrients, saltern systems all share one common feature: They all have a salinity gradient along which communities inhabiting salt marshes are structured (Britton and Johnson 1987; Ayadi et al. 2004; Abid et al. 2008; Madkour and Gaballah 2012; Asencio 2013). However, some other parameters may also influence species distribution in salterns, such as the concentrations of phosphorus and nitrates (Joint et al. 2002; Dolapsakis et al. 2005) and the distance of each pond to the sea intake (Abid et al. 2008; Madkour and Gaballah 2012). Nutrient concentrations usually increase with salinity, and this can be due either to greater evaporation and/or to bacterial activity (Britton and Johnson 1987; Elloumi et al. 2007, 2009; Oren 2009).
Many phytoplankton species have specific optimum and tolerance levels for some environmental parameters, such as the pH, salinity, temperature, and the availability of nutrients and light (Lim et al. 2001; Resende et al. 2005). With regard to salinity, halophiles sensu largo are organisms that are able to live in hypersaline environments, such as natural and artificial salterns. Species living in salty water can be either halophilic stricto sensu, requiring a given NaCl concentration, or simply halotolerant (Larsen 1986; Chatchawan et al. 2011); they can be loosely classified as slightly, moderately, or extremely halophilic/halotolerant, depending on their requirement/tolerance for NaCl (Larsen 1986; DasSarma and Arora 2001).
Multivariate analyses are the preferred tools used to identify the links between environmental and biological parameters, and to understand the structure of ecological communities (Marques et al. 2011). In particular, correspondence analysis (Sylvestre et al. 2001), cluster analysis (Dolapsakis et al. 2005), canonical correspondence analysis (Lopez-Flores et al. 2006), and principal component analysis (PCA, Anneville et al. 2004; Grover and Chrzanowski 2006) are often used in ecological studies. In the Sfax saltern, the phytoplankton has been extensively studied using PCA (Ayadi et al. 2002; Elloumi et al. 2009; Kobbi-Rebai et al. 2013). However, since physical and biological processes are frequently intercorrelated and generally show complex patterns of variation, more structured analyses could be useful to understand the interactions. Few tools take into account the temporal variation of environmental parameters and biological assemblages, or the effect of the former on the latter (Carassou and Ponton 2007). The STATICO method (Thioulouse et al. 2004) is particularly suitable for studying the relationship between species abundance and environmental parameters when both vary over time. This method has previously been used by Mendes et al. (2009) to study the spatiotemporal structure of diatom assemblages in a temperate estuary (Ria de Aveiro, Western Portugal).
Most ecological studies carried in salterns have been conducted over periods of 1 year (e.g., Sylvestre et al. 2001; Lopez-Flores et al. 2006; Elloumi et al. 2009; Asencio 2013). An interannual study elaborated by Khemakhem et al. (2010) focused on phytoplankton succession in the salt marshes of Sfax. They found only minor differences between the different years studied (from 2000 to 2003), because of the interannual stability of the environmental conditions.
The present study set out firstly to characterize the phytoplankton community structure along the Sfax saltern in relation to environmental variables from June 2010 to May 2011, and to analyze its dynamics using the STATICO method. The second aim was to determine the ecological preferences of phytoplankton taxa by estimating their optimum and tolerance for the most influential variable in the solar ponds of Sfax.
Materials and methods
Study area and sampling site
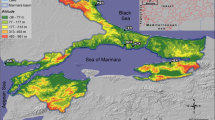
The Thyna saltworks (Fig. 1; Sfax, Tunisia), also known as the Sfax saltern, are located in the Gulf of Gabes (central eastern coast of Tunisia, 34°390′0.1″N–420′35E, Fig. 1) and cover 1,500 ha along a 13-km stretch of the Mediterranean coast (Kobbi-Rebai et al. 2013). They are classified as a RAMSAR (the Convention on Wetlands of International Importance, especially as Waterfowl Habitat) site and important bird area (IBA), since they are one of the most important crossroads for migratory birds that come from Europe and Sub-Saharan Africa and are breeding sites for several species of waterfowl (Ramírez et al. 2011). They consist of a series of interconnecting shallow ponds (20–70 cm deep) of increasing salinity, ranging from that of seawater to saturation (Khemakhem et al. 2010). The saltern is separated from the sea by an artificial red silt seawall (height: 4 m). The input of seawater and the circulation between the various ponds are controlled in order to ensure an annual yield of about 300,000 tons of halite and 25,000 tons of brine (Abid et al. 2008), but still depend on the meteorological conditions.

Map of the Thyna saltworks of Sfax (Tunisia) indicating the ponds sampled (designated A5, A16, C21, C41, M2, B1, and TS) and the direction of the water flow in the saltern
Seven ponds were chosen as sampling sites in the Thyna saltworks. They were located along the salinity gradient and designated A5, A16, C21, C41, M2, B1, and TS (Fig. 1). Sampling was performed fortnightly, over a period of 10 months extending over a period of 1 year (from June 2010 to May 2011, excluding January and February).
Environmental data
At each sampling, the water temperature was measured in situ, using a mercury glass thermometer with 0.1 °C graduations. Water samples for chemical analyses were collected and immediately stored under cool (4 °C), dark conditions, until further processing was possible. In the laboratory, these water samples were filtered through GF/C filters (0.47-µm pore). The filtrates were used to determine nutrient contents (NO2 −, NO3 −, NH4 + and PO4 3−, total nitrogen and total phosphorus). They were analyzed using a BRAN and LUEBBE type 3 autoanalyzer, and concentrations were determined colorimetrically using a UV–visible spectrophotometer. The pH was measured using a Metrohm® type pH meter. The dissolved oxygen content was calculated according to the equation of Sherwood et al. (1992). Brine salinity was assessed using a ZUZI-type refractometer.
Biological data
Water samples (200 mL) were fixed with Lugol’s iodine solution, and the phytoplankton organisms were counted under an inverted microscope (400×) using the Utermöhl (1958) method. Phytoplankton species identification was based on morphological criteria after consulting the standard floras of Sournia (1986), Ricard (1987), and Chrétiennot-Dinet (1990). Phytoplankton diversity (H′) was estimated using the Shannon–Weaver’s index (Shannon and Weaver 1949).
Statistical analysis
Kruskal–Wallis tests were used to identify the significant differences between the ponds sampled with regard to physico-chemical variables. These tests were chosen because of non-normal distribution of the data. The significance level was set at 0.05. Correlations between variables were calculated when necessary using Spearman’s rank correlation coefficient. Salinity optima (û k ) and tolerance (t k ) values were estimated for each species, after sampling at least five times, according to Resende et al. (2005):
where û k is the optimum value, t k is the tolerance of the taxon k for the parameter in cause, y ik is the abundance of the taxon k in the sample i, and x i is the value of the parameter in the sample i.
The STATICO analysis was used to explain the dynamics of the relationships between the species and their environment (Thioulouse et al. 2004). To do this, our data were processed using pairs of ecological tables: one table for the environmental variables, and the other for the species abundances for each sampling date. The 11 environmental variables and the abundances of 64 species were reported in columns in their respective tables. The sites sampled were indicated in rows and were the same in all the tables. A log(x + 1) transformation was applied to the species abundances prior to carrying out the calculations (Legendre and Legendre 1979) in order to minimize the effect of unusual catches.
STATICO performs a partial triadic analysis of a series of cross-tables resulting from the co-inertia analysis of each pair of tables (Thioulouse et al. 2004; Thioulouse 2011). It proceeds in three stages: (1) analyzing each table by a one-table method (normed PCA for the environmental variables, and centered PCA for the species data); (2) analyzing each pair of tables by a co-inertia analysis (Dolédec and Chessel 1994), which provides an image of the co-structure (species variables) at each sampling date; and (3) performing a partial triadic analysis (Thioulouse and Chessel 1987) on the resulting tables to get an average image of the co-structure. The third step reveals the part of the structure between the environment and species abundance tables that remains stable throughout the sampling period (known as the compromise). STATICO can also be used to plot the projection of the sites sampled at each sampling date onto the compromise axes, in terms of environmental variables and species abundance structures. This makes it possible to discuss the temporal variation of the species-environment structure (trajectories).
Analyses were performed using R (version 3.1.0, R Core Team 2014). The STATICO algorithm is implemented in the ade4 library of R.
Results
Environmental parameters
The range of the environmental variables in each pond and the Kruskal–Wallis tests are presented in Table 1. A clear longitudinal salinity gradient was observed along the saltern, following the direction of the water flow. Increasing salinity was found from the first studied pond (A5) to the last one (TS), rising from 42 ± 2.5 in A5 to 346.30 ± 41.4 in TS. Dissolved oxygen decreased with salinity (ρ = −0.94, p = 3.10−68), falling from 6.55 ± 0.07 mg L−1 in the least salty pond to 2.03 ± 0.07 mg L−1 in the saltiest pond. Mean water temperature showed no significant variation between the ponds, fluctuating between 25 ± 6.45 and 27 ± 6.63 °C. The lowest temperature (15 °C) was recorded in March 2011 in C21, and the highest temperature (39 °C) was recorded in August 2010 in M2. In general, the pH decreased with salinity (ρ = −0.40, p = 8.10−7), ranging from 8.23 ± 0.45 in A16 to 7.36 ± 0.42 in TS.
With regard to nutrients, ammonium (NH4 +) and total nitrogen (T-N) showed no significant variation among the ponds. Nitrites (NO2 −), nitrates (NO3 −), orthophosphates (PO4 3−), total phosphorus (T-P), and silicates (SiO3 2−) tended to increase with salinity (ρ = 0.33, 0.23, 0.21, 0.21, 0.20, p = 5.10−5, 5.10−3, 0.01, 0.01, 0.01, respectively).
Phytoplankton assemblage
The overall diversity of phytoplankton taxa was clearly linked to salinity (Table 1). H′ values were significantly different among the ponds and fell from A5 (H′ = 2.11) to TS (H′ = 0.38), indicating that the phytoplankton communities were far more diverse in the least salty ponds than in the saturation pond.
Phytoplankton was examined at the level of the genus or species. A total of 64 phytoplankton taxa were identified during this study. They belonged to five classes: Bacillariophyceae, Dinophyceae, Chlorophyceae, Cyanophyceae, and Euglenophyceae. Marked differences in phytoplankton composition were found among the ponds (Fig. 2); the relative abundance of each species compared to the total phytoplankton is presented in Table 2. Bacillariophyceae and Dinophyceae were the most diverse groups. Bacillariophyceae or diatoms represented 91.7, 77.5, and 61 % of the total phytoplankton in the A5, C21, and C41 ponds, respectively, in which salinity ranged from 42 to 96. The three most abundant species of Bacillariophyceae were Cylindrotheca closterium, Nitzschia spp., and Navicula spp (Table 2). Dinophyceae were numerous in the second and third ponds (A16 and C21), which had salinity values of 61 and 88, respectively, where they contributed for 37.12 and 30.84 %, respectively, to the total abundance of the phytoplankton. This class was virtually absent in salty ponds (0.39 and 0.01 % in the M2 and B1 ponds, respectively). The most abundant species in this class throughout the sampling period were Gymnodinium spp. and Peridinium spp. Chlorophyceae were found in all ponds. Their contribution did not exceed 13.1 % in the four first ponds (A5 to C41), whereas this class markedly dominated the phytoplanktonic community in the three last ponds (M2, B1, and TS), accounting for 73.83, 68.39, and 92.85 % of the total phytoplankton, respectively. The Cyanophyceae were represented by four species: Aphanothece sp., Oscillatoria spp., Phormidium versicolor, and Spirulina subsalsa. They were rare in the four first ponds (0.01, 0.04, 3.59, and 0.25 %, respectively) and relatively abundant in the M2 (13.66 %), B1 (31.31 %), and TS ponds (7.02 %). S. subsalsa is the least salt tolerant Cyanophyceae in our samples. It was detected in the A5, A16, C41, and M2 ponds, and flourished in the C21 pond, but was unable to proliferate in the ponds with high salinity (B1, TS). Euglenophyceae were poorly represented by Euglena sp. in the A5 pond and rarely found in saltier ponds. Globally, we can see a decrease in the number of taxa as the salinity increased (Table 2).

Contribution of the phytoplanktonic groups to the total phytoplanktonic density in the seven ponds of increasing salinity. (Euglenophyceae corresponded to less than 0.2 %)
In summary, species composition differed significantly from one pond to another. The presence of a particular species in one pond and its absence from another depended mainly on its salt tolerance. The calculation proposed by Resende et al. (2005) makes it possible to determine the optimum salinity for the growth of each species. Tolerance is a complementary parameter that indicates the range of salinity values in which the species is found (Table 2). A few species may be classified as marine (Grammatophora sp., Euglena sp.); they have a low salinity optimum (44–45) and a low tolerance (4.8–6.2). They are found only in the least salty ponds. On the contrary, some species may be classified as extremely halotolerant such as the Cyanophyceae Oscillatoria spp., Aphanotece sp., and P. versicolor, and the Chlorophyceae D. salina. These taxa have a high salinity optimum (216–245) and a medium to high salinity tolerance (44–106). They are found in all the ponds, mostly in the most salty ones. All those species form a continuum between the extreme salinity values. The species with a high salinity optimum usually have a quite high tolerance; all the species found in the most salty ponds are also found in the other ponds.
Statico
Interstructure
The first two axes of the interstructure graph represented, respectively, 24 and 7 % of the total variability (Fig. 3a). The first axis is largely predominant, and all the dates are on the same side of this axis (Fig. 3b). This indicates that the intertable correlations are all positive and that there are no strong discrepancies among the samplings. Therefore, a common structure can be highlighted (see “Compromise factor map”).

Interstructure of the STATICO analysis. a Eigenvalues barplot. b Interstructure factor map. The lengths of the arrows, in projection on the first axis, reflect the weight of the sampling dates in the analysis. c Typological value indices plot. The weight corresponds to the contribution of each table to the construction of the compromise and cos2 indicates the fit of each table to the compromise. June to December correspond to the year 2010, and March to May to the year 2011
Arrow lengths in the interstructure graph indicate that the samplings performed on November 9 and December 6 had the greatest structure, followed by those of July 13, March 10, October 5, and August 24 (Fig. 3b). This means that the relationships among environmental variables and species composition of the phytoplankton communities are the strongest at those dates.
Analysis of the weights and cos2 (Fig. 3c) informs about the contribution of each table to the compromise (weight) and shows how much the compromise reflects the information of the table (cos2). We can see a linear relationship between them, i.e., the compromise is particularly representative of the most contributive tables which are the most structured ones (November 9, December 6, July 13, March 10, and October 5). The samplings performed on June 29, May 10, August 10, April 12, and July 27 have the lowest weights and are the least well represented by the analysis. Globally, the samplings of the spring and the summer are less contributive and are less well represented by the compromise than those of the autumn and the winter.
Compromise factor map
The first axis is clearly dominant, explaining 88.1 % of the variance, whereas the second axis explains only 3.2 % of the variance (Fig. 4). Considering the environmental variables (Fig. 4a), the first axis describes a gradient of salinity opposed to a gradient of dissolved oxygen. These two variables were clearly the most influent on the stable part of the species distribution. The pH is on the low-salinity side (right side) of the factor map, and high temperature is on the high-salinity side. The nutrient elements and silicates are relatively abundant on the high-salinity side. The second axis mainly shows an N/P gradient, with nitrogen ions (particularly NH4 +) at the bottom and phosphate ions at the top of the factor map.

Compromise factor maps of the STATICO analysis. a Projection of the environmental variables onto the factor map. Variable codes are indicated in Table 1. Boxes mean projection of the environmental variables for each pond. b Projection of the species onto the factor map. Species codes are indicated in Table 2. Colors are used to distinguish the classes: green Chlorophyceae; pale blue Cyanophyceae; dark blue Dinophyceae; red Bacillariophyceae; and purple Euglenophyceae. The font size is indicative of the halotolerance level of the species. Boxes mean projection of the species for each pond. The superimposition of the two maps provides information about the stable part of the relationships between environment and phytoplankton. (Color figure online)
Most of the species found (Fig. 4b), mainly diatoms and Dinophyceae, which have a low or medium optimum salinity, are located on the low-salinity side. Only Cyanobacteria and green algae, which have a high optimum salinity and/or a high salinity tolerance value, are located on the high-salinity side (Table 2). Concerning the second axis, the mean coordinate of the diatom species is positive whereas that of the Dinophyceae is negative. Hence, diatoms and Dinophyceae seem to be, respectively, associated with rather high PO4 3− and NH4 + values.
The average position of each pond projected on the factor map is heavily influenced by salinity. The ponds (A5–A16), C21, C41, M2 (B1–TS) are ordered from right to left along the first axis, which corresponds to their succession in the water flow and hence to their salinity levels. This ordination of the ponds relative to the first axis was globally the same for species and for environmental variables, indicating that the composition of the phytoplanktonic communities depends above all on the salinity levels in the different ponds. Relative to the second axis, the ponds A5 and B1, which had very high PO4 3− values (Table 1), have positive coordinates for the environmental variables. A16, C21, C41, and TS, which had lower PO4 3− values, have negative coordinates. The central position of M2 indicates that this pond is badly represented by the compromise. Concerning the species, the first pond (A5), containing nearly exclusively diatoms, has a positive coordinate, whereas the next ponds (A16, C21, and C41), containing proportionately fewer diatoms and more numerous Dinophyceae, have negative coordinates. The last ponds (M2, B1, and TS), containing very few diatoms and Dinophyceae, have nearly null coordinates.
Trajectories
In addition to this overall pattern, the relationships between environment and species vary over time. The projections of the environmental variables (Fig. 5a) and of the species (Fig. 5b) on the comprise factor map at the most influent dates make it possible to visualize these variations. The most influent variables, salinity and dissolved oxygen, located on the first axis, were subjected to very little variation (Fig. 5a). Since the entire structure relies nearly exclusively on this first axis, the structure was globally very stable. Indeed, diatoms and Dinophyceae were usually located on the right side of the factor map, while Chlorophyceae and Cyanophyceae usually appeared on the left side (Fig. 5b), indicating the relative stability of the distribution of the classes in the different ponds, depending on their salinity levels, throughout the whole year. On the contrary, the influence of the variables related to N and P was more variable (Fig. 5a). Even if these variables have a limited influence, the consequences of those variations on the composition of the communities are visible on Fig. 5b. The Dinophyceae seem to be globally associated with high NH4 + values. Their global distribution was rather stable, except in March when their abundance decreased in A16, C21, and C41. The diatoms seem to be associated with high PO4 3− values and, from July to November, with high NO3 − values. Their global distribution was rather stable over the year. From December to March, the position of NO3 − on the factor map was decoupled from that of PO4 3− and moved to the left because of an increase in NO3 − values in the most salty ponds. At the same time, Dunaliella salina developed in these ponds. pH decreased in July in A5. At the same time, the Euglenophyceae developed in this pond.

Trajectories of the STATICO analysis for environmental variables and species. a Projection of the environmental variables onto the factor map at each date. Variable codes are indicated in Table 1. Colors indicate apparent association of some variables with a class (see Fig. 4). b Projection of the species onto the factor map at each date. Species codes are indicated in Table 2. Colors are used to distinguish the classes (see Fig. 4). (Color figure online)
The projection of the samplings on the compromise axes, in terms of environmental parameters and of species structure (Fig. 6), showed rather short arrows, indicating a strong relationship between environmental conditions and community structure in all the ponds and at all sampling dates. The projections of the ponds at each sampling dates were quite close to the mean projections (Fig. 4); the environmental and species structures were rather stable along the year. From November to March, however, the structure was slightly stronger along the first axis; the projections of the ponds A5 to C41, on the right, were more detached from those of the most salty ponds, M2 to TS, on the left, indicating higher differences in the environmental conditions and in the species structure between these two groups of ponds at these dates. This split may be explained mostly by the enrichment of the most salty ponds in NO3 − and in halotolerant species such as D. salina in the most salty ponds at these dates, causing a divergence in the community structure between the least salty ponds and the most salty ponds.

Trajectories of the STATICO analysis for the ponds. Projection of the sampling ponds onto the factor map at each date. For each pond, the filled dot indicates the projection of the environmental variables, and the circle indicates the projection of the species. The dot and the circle are linked by an arrow
Discussion
Despite the stressful effects of salt, there is a high diversity of phytoplankton taxa (64) in the saltern of Sfax compared with another one in Egypt (42) (Madkour and Gaballah 2012) and in southeast Spain (49) (Asencio 2013). However, phytoplankton diversity decreases as the salinity increases, as shown by a decrease in the diversity index H′ (Nagasathya and Thajuddin 2008; Asencio 2013). The STATICO analysis confirmed that salinity is the principal factor structuring the phytoplankton communities in the saltern. The great majority of the species are found mostly in the least salty ponds, explaining the high values of dissolved oxygen, along with the entry of seawater (Quintana et al. 1998; Asencio 2013). Indeed, dissolved oxygen can be related to the intensity of photosynthetic and respiration processes when the water depth is low (70 cm in our study) (Debelius et al. 2009). In particular, the least salty ponds shelter numerous species of diatoms and Dinophyceae and rarely Euglenophyceae. This phytoplankton composition differs from that of Las Salinas del Pinet in Spain where flagellates dominate in the least salty ponds (Asencio 2013). The saltiest ponds contain a few Chlorophyceae and Cyanophyceae, in particular the green alga D. salina and the cyanobacteria Aphanothece sp., P. versicolor, and Oscillatoria sp. These species adapted to salt are found in the saltiest ponds in numerous saline systems (DasSarma and Arora 2001; Dolapsakis et al. 2005; Oren 2009; Chatchawan et al. 2011). The distribution of the classes along the salinity gradient in the Sfax saltern is the same as that encountered in most salt marches (Dolapsakis et al. 2005; Oren 2009; Madkour and Gaballah 2012).
The distribution of the species in the saltern reflects their ability to develop in more or less salty water. Chatchawan et al. (2011) defined the halophilic species sensu stricto as species that require a salty environment, while halotolerant ones are species that are able to grow in the presence of NaCl, but do not require it. The species found in the saltern of Sfax do not require salt. Many species are reported simultaneously in the least salty ponds, in moderately salty ponds, and sometimes also in the most salty ponds. Most species with a high optimum salinity also had a high salinity tolerance value. In summary, the phytoplankton species of the Sfax saltern are marine or more or less halotolerant species depending on the classes and even on the genus to which they belong. Overall, the diatoms, the Dinophyceae, and the Euglenophyceae were usually slightly or moderately halotolerant, whereas the Chlorophyceae and Cyanophyceae were extremely halotolerant. Beyond 70 g L−1, the salinity and the ionic composition of the solar ponds differ from those of the sea, which causes a physiological constraint on marine organisms that become unable to colonize these environments (Britton and Johnson 1987). The salinity ranges where the different species were found were very similar to those identified in several salt marches (DasSarma and Arora 2001; Dolapsakis et al. 2005; Oren 2009). The tolerance level depends on the mechanisms of salt adaptation or acclimation of the species (Rai and Gaur 2001). Some microalgae, such as the extremely halotolerant D. salina, synthesize stress metabolites, such as proline, glycine betaine, and glycerol, to ensure that the osmotic balance is maintained and expels harmful sodium ions from the cells (Mishra et al. 2008). Sodium expulsion via ion transporters ensures cellular homeostasis (Chen and Jiang 2009). In cyanobacteria, the mechanism of adaptation determines the resistance potential of the cells. Strains able to resist salinity up to 40 accumulate sucrose and trehalose, strains able to resist salinity up to 100 accumulate glycosylglycerol, and strains able to resist salinity up to 156, such as Aphanothece sp., P. versicolor, and Oscillatoria sp., accumulate glycine betaine or glutamate betaine (Karandashova and Elanskaya 2005; Oren 2009). Diatoms, including Amphora, Nitzschia, and Navicula species, were commonly found, but rarely abundant in hypersaline environments. Although osmoregulation has not been studied extensively in diatoms, some species have been reported to accumulate proline and oligosaccharides (DasSarma and Arora 2001).
Nutrients are more abundant in the saltiest ponds than in the least salty ponds. This may be due to a higher nutrient uptake in the least salty pounds where phytoplankton is more abundant (Yin et al. 2000; Arrigo 2005; Sterner et al. 2008; Fehling et al. 2012), to progressive evaporation along the water flow, which concentrates the nutrients, and to the intense recycling of organic matter at high salinity (Britton and Johnson 1987; Quintana et al. 1998; Davis and Giordano 1996; Joint et al. 2002; Wieland and Kühl 2006). Nutrients are also structured along the second axis of the STATICO analysis, with phosphate at the top of the factor map and nitrogen ions at the bottom. The nitrate/phosphate ratio could therefore be a secondary structuring factor for phytoplankton communities. Dinophyceae seem to be favored at high NH4 + values. This is in accordance with previous results since Collos et al. (2004) found that Dinophyceae used uppermost NH4 + rather than NO3 −. Dinophyceae seem to be also associated, to a lesser extent, to total nitrogen, which suggests that some of them can change their diet and convert to mixotrophy (Ismael 2003; Lopez-Flores et al. 2006; Girault et al. 2013). On the contrary, diatoms seem to be favored by high NO3 − and PO4 3− values. Similar results were obtained in the northeastern coast of the Black Sea (Silkin et al. 2014). Collos et al. (2014) and Egge (1998) have also shown that phosphate may often be a limiting factor for diatoms. The enhanced P need of diatoms, in comparison with other phytoplankton classes, may be due to stressful conditions such as those encountered in the saltern. Vieler et al. (2007) found higher amounts of phospholipids in the cells of a diatom than in those of a green microalga, and Chen et al. (2008) showed that diatoms produced higher amounts of phospholipids at high salinity. Differences in nutrient needs between Dinophyceae and diatoms can be related to their own physiological characteristics (Girault et al. 2013).
The compromise discussed above describes the average situation, whereas trajectories focus on the variation over time of biotic and abiotic factors and of the relationships between them, in each pond. The composition of the phytoplankton communities varies depending on the environmental conditions. The global distribution of the diatoms and Dinophyceae, mostly found in the least salty ponds, showed no strong variation during the year. At the species level, some species bloomed successively, all year round, which is consistent with previous findings (Gilabert 2001; Abid et al. 2008; Khemakhem et al. 2010). This dynamic may be driven by various environmental factors including nutrients, and perhaps also by stochastic events (Ortega-Mayagoitia et al. 2003). The Euglenophyceae, also found in the least salty ponds, were usually rare. Only one remarkable bloom occurred in the first pond (A5) in July, apparently due to an acidification. Indeed, some Euglenophyceae are acidophilic (Olaveson and Stokes 1989). In the saltiest ponds, Chlorophyceae and Cyanophyceae thrived during the autumn and winter, which is consistent with what has been reported in some other systems (Andersson et al. 1994; Montoya 2009; Bazzuri et al. 2010; Chatchawan et al. 2011). The development of the green alga D. salina persisted in spring, as reported in some other hypersaline environments in USA and Greece (Stephens and Gillespie 1976; Dolapsakis et al. 2005; Salm et al. 2009). In our study, this seems to have been due to an increase in nitrogen ions, particularly nitrates, which could be explained by an increase in bacterial nitrifying activity (Pedros-Alio et al. 2000; Madkour and Gaballah 2012; Asencio 2013) in spring, linked to warmer temperatures (Elloumi et al. 2009).
Our study suggests that nutrients have an influence on the distribution and dynamics of phytoplankton, even if this influence is much lower than that of salinity. Physiological differences among phytoplankton taxa with regard to nutrient uptakes (Arrigo 2005; Lopez-Flores et al. 2006) are consistent with this hypothesis. Several studies have shown that nutrients play a critical role in saline environments (Moll 1977; Davis 2000; Ortega-Mayagoitia et al. 2003; Salm et al. 2009). This STATICO analysis has shown some global trends, but further work is needed to elucidate the influence of the principal nutrients in details and their interactions with climatic factors, zooplankton, and bacteria. Some other important parameters, such as trace elements, should also be taken into account in future studies. As reminded by Asencio (2013), salinization of waters is expected to rise worldwide as a result of climate changes (IPCC 2007). To ensure the production of salt of good quality in the future, it is necessary to well understand the functioning of saltern in order to maintain stable species concentration and composition by management techniques because they help enhance solar absorption and eliminate dissolved organic carbon and nutrients from the water column.
References
Abid O, Sellami-Kammoun A, Ayadi H, Drira Z, Bouain A, Aleya L (2008) Biochemical adaptation of phytoplankton to salinity and nutrient gradients in a coastal solar saltern, Tunisia. Estuar Coast Shelf Sci 80:391–400
Andersson A, Haecky R, Hagström A (1994) Effect of temperature and light on the growth of micro- nano- and pico-plankton: impact on algal succession. Mar Biol 120:511–520
Anneville O, Souissi S, Gammeter S, Straile D (2004) Seasonal and inter-annual scales of variability in phytoplankton assemblages: comparison of phytoplankton dynamics in three peri- alpine lakes over a period of 28 years. Freshw Biol 49(1):98–115
Arrigo KR (2005) Marine microorganisms and global nutrient cycles. Nature 437:349–355
Asencio AD (2013) Permanent salt evaporation ponds in a semi-arid Mediterranean region as model systems to study primary production processes under hypersaline conditions. Estuar Coast Shelf Sci 124:24–33
Ayadi H, Toumi N, Abid O, Medhioub K, Hammami M, Sime-Ngando T, Amblard C, Sargos D (2002) Étude qualitative et quantitative des peuplements phyto- et zooplanctoniques dans les bassins de la saline de Sfax. Tunis Rev Sci Eau 15(1):123–135
Ayadi H, Abid O, Elloumi J, Bouaïn A, Sime-Ngando T (2004) Structure of the phytoplankton communities in two lagoons of different salinity in the Sfax saltern (Tunisia). J Plankton Res 26:669–679
Bazzuri ME, Gabellone N, Solaril L (2010) Seasonal variation in the phytoplankton of a saline lowland river (Buenos Aires, Argentina) throughout an intensive sampling period. River Res Appl 26:766–778
Britton RH, Johnson AR (1987) An ecological account of a Mediterranean salina: the Salin de Giraud, Camargue (S. France). Biol Conserv 42:185–230
Carassou L, Ponton D (2007) Spatio-temporal structure of pelagic larval and juvenile fish assemblages in coastal areas of New Caledonia, southwest Pacific. Mar Biol 150:697–711
Chatchawan T, Peerapornpisal Y, Komárek J (2011) Diversity of cyanobacteria in man-made solar saltern, Petchaburi Province, Thailand—a pilot study. Fottea 11:203–214
Chen H, Jiang JG (2009) Osmotic responses of Dunaliella to the changes of salinity. J Cell Physiol 219:251–258
Chen GQ, Jiang Y, Chen F (2008) Salt-induced alterations in lipid composition of diatom Nitzshia laevis (Bacillariophyceae) under heterotrophic culture condition. J Phycol 44:1309–1314
Chrétiennot-Dinet MJ (1990) Atlas du phytoplancton marin volume 3: Chlororachinophycées, chlorophycées, chrysophycées, cryptophycées, euglénophycées, eustigmatophycées, prasinophycées, prymnésiophycées, rhodophycées, tribophycées. Éditions du centre national de la recherche scientifique, Paris
Collos Y, Gagne C, Laabir M, Vaquer A, Cecchi P, Souchu P (2004) Nitrogenous nutrition of Alexandrium catenella (Dinophyceae) in cultures and in Thau lagoon, Southern France. J Phycol 40:96–103
Collos Y, Jauzein C, Ratmaya W, Souchu P, Abadie E, Vaquer A (2014) Comparing diatom and Alexandrium catenella/tamarense blooms in Thau lagoon: importance of dissolved organic nitrogen in seasonally N-limited systems. Harmful Algae 37:84–91
DasSarma S, Arora P (2001) Halophiles, encyclopedia of life sciences. Nature Publishing Group. www.els.net. Accessed March 2012
Davis J (2000) Structure, function and management of the biological system for seasonal solar saltworks. Glob Nest 2:217–226
Davis J, Giordano M (1996) Biological and physical events involved in the origin, effects, and control of organic matter in solar saltworks. Int J Salt Lake Res 4:335–347
Debelius B, Gomez-Parra A, Forja JM (2009) Oxygen solubility in evaporated seawater as a function of temperature and salinity. Hydrobiologia 632:157–165
Dolapsakis NP, Tafas T, Abatzopoulos TJ, Ziller S, Economou-Amilli A (2005) Abundance and growth response of microalgae at Megalon Embolon solar saltworks in northern Greece: an aquaculture prospect. J Appl Phycol 17:39–49
Dolédec S, Chessel D (1994) Co-inertia analysis: an alternative method for studying species—environment relationships. Freshw Biol 31:277–294
Dufour P, Durand JR (1982) La production végétale des lagunes de Côte d’Ivoire. Rev Biol Trop 15:209–230
Egge JK (1998) Are diatoms poor competitors at low phosphate concentrations? J Mar Syst 16:191–198
Elloumi J, Guermazi W, Ayadi H, Bouaïn A, Abderrahmen, Aleya L (2007) Detection of water and sediments pollution of an arid saltern (Sfax, Tunisia) by coupling the distribution of microorganisms with hydrocarbons. Water Air Soil Pollut 187:157–171
Elloumi J, Carrias JF, Ayadi H, Sime-Ngando T, Bouaïn A (2009) Communities structure of the planktonic halophiles in the solar saltern of Sfax, Tunisia. Estuar Coast Shelf Sci 81:19–26
Fehling J, Davidson K, Bolch CJS, Brand TD, Narayanaswamy BE (2012) The relationship between phytoplankton distribution and water column characteristics in north west european shelf sea waters. PLoS One 7:e34098. doi:10.1371/journal.pone.0034098
Gilabert J (2001) Seasonal plankton dynamics in a Mediterranean hypersaline coastal lagoon: the Mar Menor. J Plankton Res 23:207–217
Girault M, Arakawa H, Hashihama F (2013) Phosphorus stress of microphytoplankton community in the western subtropical North Pacific. J Plankton Res 35:146–157
Grover JP, Chrzanowski TH (2006) Seasonal dynamics of phytoplankton in two warm temperate reservoirs: association of taxonomic composition with temperature. J Plankton Res 28:1–17
IPCC (Intergovernmental Panel on Climatic Change) (2007) Fourth assessment report. In: Climatic change impacts, adaptation and vulnerability. http://www.ipcc.ch/
Ismael AA (2003) Succession of heterotrophic and mixotrophic dinoflagellates as well as autotrophic microplankton in the harbour of Alexandria. Egypt J Plankton Res 25:193–202
Joint I, Henriksen P, Garde K, Riemann B (2002) Primary production, nutrient assimilation and microzooplankton grazing along a hypersaline gradient. FEMS Microbiol Ecol 39:245–257
Karandashova IV, Elanskaya IV (2005) Genetic control and mechanisms of salt and hyperosmotic stress resistance in cyanobacteria. Russ J Genet 41:1311–1321
Khemakhem H, Elloumi J, Moussa M, Aleya L, Ayadi H (2010) The concept of ecological succession applied to phytoplankton over four consecutive years in five ponds featuring a salinity gradient. Estuar Coast Shelf Sci 88:33–44
Kobbi-Rebai R, Annabi-Trabelsi N, Khemakhem H, Ayadi H, Aleya L (2013) Impacts of restoration of an uncontrolled phosphogypsum dumpsite on the seasonal distribution of abiotic variables, phytoplankton, copepods, and ciliates in a man-made solar saltern. Environ Monit Assess 185:2139–2155
Koffi K, Philippe DK, Marcel KA, Maryse AN, Kagoyire KA, Adingra AA (2009) Seasonal distribution of phytoplankton in Grand-Lahou Lagoon (Côte d’Ivoire). Eur J Sci Res 26:329–341
Larsen H (1986) Halophilic and halotolerant microorganisms—an overview and historical perspective. FEMS Microbiol Lett 39:3–7
Legendre L, Legendre P (1979) Ecologie numérique 1. Masson, Paris
Lim DSS, Douglas MSV, Smol JP (2001) Diatoms and their relationship to environmental variables from lakes and ponds on Bathurst Island, Nunavut, Canadian High Arctic. Hydrobiologia 450:215–230
Lopez-Flores R, Boix D, Badosa A, Brucet S, Quintana XD (2006) Pigment composition and size distribution of phytoplankton in a confined Mediterranean salt marsh ecosystem. Mar Biol 149:1313–1324
Maar M, Nielsen TG, Richardson K, Christaki U, Hansen OS, Zervoudaki S, Christou ED (2002) Spatial and temporal variability of food web structure during the spring bloom in the Skagerrak. Mar Ecol Prog Ser 239:11–29
Madkour FF, Gaballah MM (2012) Phytoplankton assemblage of a solar saltern in Port Fouad. Egypt Oceanol 54:687–700
Mallin MA, Paerl HW (1994) planktonic trophic transfer in an estuary: seasonal, diel, and community structure effects. Ecology 75:2168–2184
Marques SC, Pardal MÂ, Mendes S, Azeiteiro UM (2011) Using multitable techniques for assessing the temporal variability of species environment relationship in a copepod community from a temperate estuarine ecosystem. J Exp Mar Biol Ecol 405:59–67
Mendes S, Fernandez-Gomez MJ, Resende P, Pereira MJ, Galindo-Villardon MP, Azeiteiro UM (2009) Spatio- temporal structure of diatom assemblages in a temperate estuary. A STATICO analysis. Estuar Coast Shelf Sci 84:637–644
Mishra A, Mandoli A, Jha B (2008) Physiological characterization and stress-induced metabolic responses of Dunaliella salina isolated from salt pan. J Ind Microbiol Biotechnol 35:1093–1101
Moll RA (1977) Phytoplankton in a temperate-zone salt marsh: net production and exchanges with coastal waters. Mar Biol 42:109–118
Montoya H (2009) Algal and cyanobacterial saline biofilms of the Grande Coastal Lagoon, Lima, Peru. Nat Resour Environ Issue 15:127–134
Nagasathya A, Thajuddin N (2008) Cyanobacterial diversity in the hypersaline environment of the saltpans of southeastern coast of India. Asian J Plants Sci 7:473–478
Olaveson MM, Stokes PM (1989) Responses of the acidophilic alga Euglena mutabilis (Euglenophyceae) to carbon enrichment at pH 3. J Phycol 25:529–539
Oren A (2009) Saltern evaporation ponds as model systems for the study of primary production processes under hypersaline conditions. Aquat Microbiol Ecol 56:193–204
Ortega-Mayagoitia E, Rojo C, Rodrigo MA (2003) Controlling factors of phytoplankton assemblages in wetlands: an experimental approach. Hydrobiologia 502:177–186
Pedros-Alio C, Calderon-Paz JI, MacLean MH, Medina G, Marrase C, Gasol JM, Guixa-Boixereu N (2000) The microbial food web along salinity gradients. FEMS Microbiol Ecol 32:143–155
Pinckney JL, Paerl HW, Harrington MB, Howe KE (1998) Annual cycles of phytoplankton community- structure and bloom dynamics in the Neuse River Estuary, North Carolina. Mar Biol 131:371–381
Quintana XD, Amich RM, Comin FA (1998) Nutrient and plankton dynamics in a Mediterranean salt marsh dominated by incidents of flooding. Part 1: differential confinement of nutrients. J Plankton Res 20:2089–2107
Rai LC, Gaur JP (2001) Algal adaptation to environmental stresses: physiological, biochemical and molecular mechanisms. Springer, Berlin
Ramírez F, Abdennadher A, Sanpera C, Jover L, Wassenaar LI, Hobson KA (2011) Assessing waterbird habitat use in coastal evaporative systems using stable isotopes (δ13C, δ15N and δD) as environmental tracers. Estuar Coast Shelf Sci 92:217–222
Resende P, Azeiteiro U, Pereira MJ (2005) Diatom ecological preferences in a shallow temperate estuary (Ria de Aveiro, Western Portugal). Hydrobiologia 544:77–88
Ricard M (1987) Atlas du phytoplancton marin volume 2: Diatomophycées Éditions du centre national de la recherche scientifique, Paris
Richardson K, Pedersen FB (1998) Estimation of new production in the North Sea: consequences for temporal and spatial variability of phytoplankton. J Mar Sci 55:574–580
Richardson K, Nielsen TG, Bo Pedersen F, Heilmann JP, Lokkegaard B, Kaas H (1998) Spatial heterogeneity in the structure of the planktonic food web in the North Sea. Mar Ecol Prog Ser 168:197–211
Salm CR, Saros JE, Martin CS, Erickson JM (2009) Patterns of seasonal phytoplankton distribution in prairie saline lakes of the Northern Great Plains (USA). Saline Syst 5:1448–1746
Shannon CE, Weaver G (1949) The mathematical theory of communication. University of Illinois Press, Urbana
Sherwood JE, Stagnitti F, Kokkinn MJ, Williams WD (1992) A standard table for predicting equilibrium dissolved oxygen concentrations in salt lakes dominated by sodium chloride. J Int J Salt Lake Res 1:1–6
Silkin VA, Pautova LA, Pakhomova SV, Lifanchuk AV, Yakushev EV, Chasovnikov VK (2014) Environmental control of phytoplankton community structure in the NE Black Sea. J Exp Mar Biol Ecol 461:267–274
Sournia A (1986) Atlas du phytoplancton marin volume 1: Cyanophycées, dictyochophycées, dinophycées et raphidophycées. Éditions du centre national de la recherche scientifique, Paris
Stephens DW, Gillespie DM (1976) Phytoplankton production in the Great Salt Lake, Utah, and a laboratory study of algal response to enrichment. Limnol Oceanogr 21:74–87
Sterner RW, Andersen T, Elser JJ, Hessen DO, Hood JM, McCauley E, Urabe J (2008) Scale-dependent carbon: nitrogen: phosphorus seston stoichiometry in marine and freshwaters. Limnol Oceanogr 53:1169–1180
Sylvestre F, Beck-Eichler B, Duleba W, Debenay JP (2001) Modern benthic diatom distribution in a hypersaline coastal lagoon: the Lagoa de Araruama (RJ), Brazil. Hydrobiologia 443:213–231
R Core Team (2014) R: A language and environment for statistical computing. R foundation for statistical computing, Vienna, Austria. URL http://www.R-project.org/
Thioulouse J (2011) Simultaneous analysis of a sequence of paired ecological tables: a comparison of several methods. Ann Appl Stat 5:2300–2325
Thioulouse J, Chessel D (1987) Les analyses multitableaux en écologie factorielle. Acta Oecol-Oec Gener 8:463–480
Thioulouse J, Simier M, Chessel D (2004) Simultaneous analysis of a sequence of paired ecological tables. Ecology 85:272–283
Trigui H, Masmoudi S, Brochier-Armanet C, Maalej S, Dukan S (2011) Characterization of Halorubrum sfaxense sp. nov., a new halophilic archaeon isolated from the solar saltern of Sfax in Tunisia. Int J Microbiol. doi:10.1155/2011/240191
Utermöhl H (1958) Zur Vervollkommung der quantitativen Phytoplankton Methodik. Mitteilungen Internationale Vereinigung für Theoretische und Angewandte. Limnologie 9:1–38
Ventosa A, Arahal DR (2009) Physico-chemical characteristics of hypersaline environments and their biodiversity. Extremophiles 2:1–6
Vieler A, Wilhelm C, Goss R, Süß R, Schiller J (2007) The lipid composition of the unicellular green alga Chlamydomonas reinhardtii and the diatom Cyclotella meneghiniana investigated by MALDI- TOF MS and TLC. Chem Phys Lipids 150:143–155
Vos J, De la Rosa NL (1980) Manual on Artemia production in salt ponds in the Philippines. Quezon City. In: BFAR/UNDP/FAO, Brackishwater aquaculture and demonstration training project, PHI/75/005, FAO, Rome p 49
Wen Z, Zhi-Hui H (1999) Biological and ecological features of inland saline waters in North Hebei. China Int J Salt Lake Res 8:267–285
Wieland A, Kühl M (2006) Regulation of photosynthesis and oxygen consumption in a hypersaline cyanobacterial mat (Camargue, France) by irradiance, temperature, and salinity. FEMS Microbiol Ecol 55:195–210
Yin K, Qian PY, Chen JC, Hsieh DPH, Harrison PJ (2000) Dynamics of nutrients and phytoplankton biomass in the Pearl River estuary and adjacent waters of Hong Kong during summer: preliminary evidence for phosphorus and silicon limitation. Mar Ecol Prog Ser 194:295–305
Acknowledgments
The authors would like to express their special thanks to COTUSAL staff for giving us access to the saltern and permission to take samples. This work was conducted as part of a collaborative project between the University of Sfax (Tunisia) and the University of Maine (France). This study was supported by the Tunisian Ministry of Scientific Research and Technology. We thank Monika Ghosh for correcting the English language.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Bas W. Ibelings.
Rights and permissions
About this article

Cite this article
Masmoudi, S., Tastard, E., Guermazi, W. et al. Salinity gradient and nutrients as major structuring factors of the phytoplankton communities in salt marshes. Aquat Ecol 49, 1–19 (2015). https://doi.org/10.1007/s10452-014-9500-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10452-014-9500-5