
Abstract
In Patagonian rocky salt marshes, the presence of the austral cordgrass Spartina densiflora provide habitat for diverse faunal assemblages. Two different mechanisms may influence the distribution and abundance patterns of these associated organisms: those generated by the biological properties of cordgrasses and those caused or mediated by the physical structure supplied by the plants. The aims of this study were: (1) to determine the effect of cordgrasses on macroinvertebrate assemblages dominating a rocky marsh and (2) to determine how much of this effect is caused by the physical structure supply by the plants. In order to achieve these objectives, we conducted two field manipulative experiments. In the first one, we manipulated the presence of defaunated transplants of cordgrass, and in the second one, we did the same with plastic cordgrass mimics simulating the architecture of Spartina densiflora. In both experiments, sessile and mobile assemblages were separately examined to evaluate whether they respond in the same way or not. Abundance and richness of mobile and sessile organisms were higher in transplant and mimic cordgrass plots compared to the controls, indicating that the presence of cordgrasses, either transplanted or artificial, has a positive effect on sessile and mobile assemblages. Furthermore, we found that the composition of mobile and sessile macroinvertebrates assemblages did not differ between transplants and mimics. Therefore, the physical structure of cordgrass was found to be the major factor influencing macroinvertebrates assemblages in the studied Patagonia rocky salt marshes. Within the period of 1 year, mobile and sessile fauna colonized several times more abundantly transplant and artificial cordgrass than the control unvegetated plots, highlighting the key role of cordgrasses in optimizing the colonization rate of macroinvertebrate communities in this rocky bottom environment. Since Spartina densiflora is invading different regions worldwide, our study may help to predict its potential effect on the invaded communities.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Habitat-forming organisms such as mussels, tube-worms and plants play an important role in structuring intertidal faunal assemblages by modifying the abiotic and biotic environment (Jones et al. 1997; Bouma et al. 2009). Since they are able to provide resources (i.e., food or space to live), ameliorate harsh physical parameters and increase habitat complexity, they are usually associated with highly diverse and abundant macrofaunal assemblages (Crain and Bertness 2006; Bouma et al. 2009). Indeed, the simple addition of physical structure by a habitat-forming species is likely to cause critical modifications in the environment. For instance, patches of standing structures, such as tubes, leaves or shells, can enhance soil accretion by favoring sediment deposition and stabilization (Terrados and Duarte 1999; Schwindt et al. 2004; Bos et al. 2007; Bouma et al. 2007). At the same time, these structures also supply new surfaces for larval attachment of sessile organisms (Ambrose and Irlandi 1992; Barnes 2000; Callaway 2003), shelter from desiccation and wave action (Gosselin and Chia 1995; Hammond and Griffiths 2004), and refuge from predators for mobile and sessile fauna (Minello and Zimmerman 1983; Gribben and Wright 2006; Canion and Heck 2009; Henninger et al. 2009). Thus, these physical structures influence the composition of associated benthic communities and may even be a necessary precondition for the presence of such communities (Schwindt et al. 2001). However, in addition to the physical structure supplied by these organisms, there are also biological properties (such as biochemical and chemical constitution, metabolic products and byproducts; e.g., Wahl 1989; Pawlik 1992; Duffy and Hay 1994; Caraco et al. 2006) and ecological properties and processes including positive, neutral and negative interactions (e.g., Bertness 1984; Lee et al. 2001; Henninger et al. 2009) intrinsically linked to them and which may also have strong effects on the associated faunal assemblages. Thus, the effects exerted by a habitat-forming organism on the associated fauna may have either abiotic or biotic origins, or both.
The rocky salt marshes (see Bortolus et al. 2009; Bortolus 2010) are located on top of hard bottoms substrata, exposed to wave action and are devoid of the meandrous tidal/estuarine channels typically seen in muddy salt marshes. In this kind of coastal marshes, the crevices and tide pools across the intertidal are literally filled up with Spartina rhizomes and roots, shell fragments and sand, what makes it look like as environmental hybrid between a rocky intertidal and a muddy salt marsh. The addition of new and abundant biogenic structure on top of a rocky wave-cut platform may have important consequences for the local faunal assemblages (Bortolus et al. 2009; Sueiro et al. 2010). The presence of the austral cordgrass Spartina densiflora in this environment is associated with a diverse faunal assemblage that includes sessile organisms, such as mussels, sea anemones and barnacles, as well as mobile organisms, such as amphipods, isopods, gastropods and terrestrial invertebrates. Some of these organisms are only found in the vegetated areas of the marsh (Bortolus 2006; Sueiro et al. 2010). Spartina densiflora’s physical structure can enhance the sedimentation rate and reduce the surface temperature and desiccation stress by shading the substrate, while providing refuge in its dense shoots, leaves, roots and rhizomes (Bortolus et al. 2002, 2004). Additionally, S. densiflora is an important food resource for benthic invertebrates (Bortolus and Iribarne 1999; Silliman and Bortolus 2003; Bortolus et al. 2004) and a source of detritus and litter for the environment (e.g., Vera et al. 2009). Thus, two different mechanisms may influence the distribution and abundance patterns of the macrofaunal species associated with S. densiflora patches, those generated by its biological properties and those caused or mediated by the physical structure supplied by the plants.
Considering the above statements, this study tests the hypothesis that the physical structure supplied by the cordgrass Spartina densiflora to the rocky bottom shoreline is the major property of the plant affecting and shaping the patterns of the associated macroinvertebrate composition, richness and abundance. If these organisms strictly utilize S. densiflora as refuge and/or as attachment surface, regardless of their biological properties, then we expect that the effects of transplanted and artificial plants on the composition, richness and abundance of associate macroinvertebrates will be similar. Therefore, the aims of this study were: (1) to determine experimentally the effect of the cordgrass Spartina densiflora on sessile and mobile macroinvertebrate assemblages dominating a Patagonian rocky marsh and (2) to determine how much of this effect is caused by the physical structure supply by the plants. Sessile and mobile organisms were considered separately because it has been proposed that the physical and biological attributes of the habitat may affect these groups differently (see Palomo et al. 2007; Sellheim et al. 2010). In order to achieve these objectives, we conducted two field experiments manipulating the presence of Spartina densiflora transplants and mimics. Our results will help to predict the potential effects of this cordgrass on the invaded communities worldwide.
Materials and methods
Study site
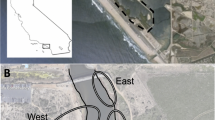
Two experiments were conducted in Ambrosetti Rocky Salt Marsh (42º50′S, 64º50′W, Patagonia, Argentina, Fig. 1). This kind of marsh develops on top of wave-cut platforms of sedimentary rock and is dominated by the austral cordgrass Spartina densiflora (Bortolus et al. 2009; Bortolus 2010). In Ambrosetti, the cordgrasses occupy the middle–high intertidal (~3.50 m above the Argentinean hydrographic zero) along a fringe of approximately 30 m wide and 370 m long, in this fringe cordgrasses typically grow as monospecific stands, with clearly defined boundaries forming patches. Within these patches, the average number of stems of S. densiflora is 14 stems·100 cm−2 (SD ± 2) with an average height of 27 cm (SD ± 6) and it has a compact mat of roots and rhizomes with an average thickness of 6.2 cm (SD ± 0.7). The roots and rhizomes are covered by the native little mussels Brachidontes purpuratus and B. rodriguezii. These mussels are also found in the unvegetated areas, while natural bare areas are not observed throughout the length and width of the vegetated fringe. The climate at this latitude is mostly arid with low precipitation (<235 mm year−1), annual temperatures ranging from 39 to −7.5°C, and strong cold winds predominantly from the southwest, with a mean annual speed of up to 22 km h−1 and gusts that may exceed 100 km h−1.

Image showing a map with the location of the experimental site, a photograph of a landscape of a typical Patagonian rocky salt marsh and a close-up of a Spartina densiflora patch. (Photograph credits: A. Bortolus)
Effect of Spartina densiflora on sessile and mobile macroinvertebrate assemblages (Experiment 1)
To evaluate the effect of Spartina densiflora on these two different groups of macroinvertebrate assemblages, we performed a transplant experiment for which we defaunated and pulled of the substrate ten clumps of S. densiflora (25 × 25 cm), which had been removed from the marsh. The macrofauna was first carefully removed by hand and later by flooding the samples in a freshwater bath for ~24 h to ensure that any remaining marine invertebrates were killed. Each clump was then carefully checked again and dead animals remaining in the clumps were removed. The use of freshwater baths to defaunate plants has been found virtually 100% effective (Virnstein and Curran 1986, and this study) without secondary effects on the plants. After this procedure, all clumps were transplanted back to the marsh (hereafter Spartina-transplants, Fig. 2A) and pinned down with stakes to randomly chosen parcels of substratum previously cleaned with scrapers. As control treatments, ten areas free of Spartina (25 × 25 cm each, Fig. 2B) on vegetated fringe were randomly chosen and were defaunated by carefully scraping the marsh surface with standard metal scrapers and by hand until the surface was free of macrofauna. Spartina-transplants were checked once a month for regular maintenance to make sure they remained pinned down to the ground throughout the experiment from December 2007 to December 2008. After this period, all treatment plots (i.e., transplants and controls) were removed and transported to the laboratory in coolers for further processing (see Sampling methods). At the same time, ten additional Spartina samples (25 × 25 cm each; hereafter “natural-Spartina,” Fig. 2C) were collected at random from the marsh in order to quantify and compare the similarity between the experimental (without historical contingencies) and the natural (with historical contingencies) scenarios.

Treatments at the beginning of the experiments. A Spartina-transplants; B control; C natural-Spartina and D Spartina-mimic
Effect of Spartina’s physical structure on sessile and mobile macroinvertebrate assemblages (Experiment 2)
The results of the previous experiment indicated differences in abundance, richness and composition of sessile and mobile macroinvertebrate assemblages between transplant and control treatments (see “Results”). To evaluate how much of this effect is caused by the physical structure supplied by the plants, a second field experiment was conducted in the same marsh. The experiment included a treatment with inert plastic models (n = 10) that resemble Spartina densiflora (hereafter Spartina-mimic, Fig. 2D). Each Spartina-mimic was constructed on a 25 × 25 cm rigid plastic grid (mesh size = 2 × 2 cm). Over this plastic grid, a plastic cable of 0.5 cm diameter was interwoven simulating the thick and compact mat of S. densiflora roots and rhizomes. Then, 40 plastic leaf-like structures (40 cm height, 0.5 cm width) were attached around the mimics to recreate S. densiflora’s physiognomy in terms of cover and density of aerial and subterranean structures (Bortolus 2006). The Spartina-mimics were then pinned down with stakes to randomly chosen parcels of substratum previously cleaned with scrapers. Again, as control treatments, ten areas free of Spartina (25 × 25 cm each, Fig. 2B), on vegetated fringe, were randomly chosen and defaunated by cleaning the surface with scrapers. After 12 months (from December 2008 to December 2009), Spartina-mimics and all the organisms found in control plots were removed from the field and transported in coolers to the laboratory for macrofaunal analyses as explained above. As in the previous experiment, ten natural-Spartina plots (25 × 25 cm each, Fig. 2C) were obtained with scrapers and transported to the laboratory in order to compare natural and experimental scenarios.
Sampling methods
The macrofauna of the experimental plots were first sorted by hand from plant material. Any remaining fauna were obtained by carefully sieving the rest of the samples through a 0.5-mm mesh. The retained material was fixed in 10% formalin for 48 h and then preserved in 70% ethanol. The organisms found were counted and identified to the lowest possible taxonomic level under stereomicroscope (80×) with the help of expert taxonomists. Then, organisms were assigned to sessile or mobile based on their mobility. Complementary within both mobility groups, the feeding type of each organism was identified in order to determine whether the abundance or taxonomic richness of any of them was particularly affected by the treatments. Considering the potential undesirable consequences of taxonomic misidentifications (Bortolus 2008), we requested the assistance of taxonomic specialists for each taxon. A voucher of each specimen collected was deposited in the CENPAT invertebrate collection (http://www.cenpat.edu.ar).
Data analysis
One-way fixed ANOVAs were used to test for differences among treatments in total abundance (total number of individuals per plot) and taxonomic richness (number of taxa per sample). Normality and homogeneity of variance assumptions were evaluated with Kolmogorov–Smirnov and Levene tests, respectively. Significant differences among mean values were evaluated with a Tukey’s post hoc test. When variances were heterogeneous and could not be stabilized after different transformations, the nonparametric Kruskal–Wallis test was used and significant results were analyzed with multiple comparisons of mean ranks (Zar 1999).
The macroinvertebrate assemblage composition was analyzed by using Primer Statistical software (Clarke and Warwick 1994). The data matrix of all invertebrate species was fourth-root transformed in order to down-weight the abundant species. Non-metric multidimensional scaling (MDS) was used to explore similarities and differences among assemblages. The significance of spatial differences between assemblages was assessed with analysis of similarity (ANOSIM). These analyses were based on Bray–Curtis similarity indexes.
Results
Twenty taxa were identified, 80% of which were mobile organisms and 20% sessile; however, sessile organisms were dominant in terms of abundance (Table 1). The mobile taxa included three feeding types: deposit-detritus feeders (mostly crustaceans), predators (including polychaetes and nemertean worms) and grazers (represented only by one species: the limpet Siphonaria lessoni). The sessile taxa included three species of suspension feeders (mostly bivalves) and a passive predator sea anemone (Table 1).
Effect of Spartina densiflora on sessile and mobile macroinvertebrate assemblages
At the end of the experiment, the transplant plots of all treatments were massively colonized by fauna and visually indistinguishable from the surrounding vegetated areas. The presence of transplanted Spartina densiflora had a positive effect on the abundance and richness of sessile and mobile macroinvertebrates (Table 2, Fig. 3A). Spartina-transplants showed mean values of mobile and sessile abundance significantly higher than controls (Table 2, Fig. 3A). Similarly, feeding types showed the same trend in their mean abundances between Spartina-transplants and controls (Table 3, Fig. 3A). After 1 year, the abundance of mobile and sessile fauna in Spartina-transplants was fourteen and four times greater, respectively, than in the controls. The taxonomic richness of mobile fauna was approximately two times larger in Spartina-transplants than controls; in this case, the observed difference was driven by deposit-detritus feeders and predators (Table 2, 3, Fig. 3A). On the other hand, sessile taxonomic richness, and consequently suspension feeder, showed no differences between Spartina-transplants and controls (Table 2, Fig. 3A).

A Means and SD for abundances and richness of sessile and mobile taxa and of each feeding type and B MDS ordinations comparing macroinvertebrate composition in Spartina-transplants (transp, black diamonds), control (cont, white circles) and natural-Spartina (nat, cross squares) treatments. Treatments with different letters resulted in significant differences (Tukey unequal N HSD tests or multiple comparisons of mean ranks, p < 0.05). Feeding types are indicated as suspension feeder (SF), deposit-detritus feeder (DDF), predator (P) and grazer (G)
Between Spartina-transplants and natural-Spartina plots, no statistical differences were found in the mean abundance and taxonomic richness of sessile fauna (Table 2, Fig. 3A). However, we found a detectable difference in the mean total abundance and taxonomic richness of mobile fauna due to the absence of the amphipods Melita palmata and Monocorophium insidiosum and also a few rare polychaetes in the Spartina-transplants (Fig. 3A). The amphipods mentioned above were also responsible for the significant differences observed between Spartina-transplants and natural-Spartina plots in mean abundance of detritus feeders (Table 3, Fig. 3A). The grazer Siphonaria lessoni showed a higher mean abundance in Spartina-transplants than natural-Spartina plots, whereas predators and suspension feeders did not show significant differences either in richness or in abundances (Table 3, Fig. 3A). The multivariate analysis indicated that the assemblage composition of sessile organisms was different between Spartina-transplants and controls (ANOSIM, R = 0.8, P = 0.001; Fig. 3B) and between controls and natural-Spartina (ANOSIM, R = 0.8, P = 0.001; Fig. 3B) but did not differ between Spartina-transplants and natural-Spartina plots (ANOSIM, R = 0.2, P = 0.02; Fig. 3B). Finally, the composition of mobile fauna differed significantly in all pairwise comparison between treatments (ANOSIM, Spartina-transplants vs. control, R = 0.9, P = 0.001; Spartina-transplants vs. natural-Spartina, R = 0.9, P = 0.001; control vs. natural-Spartina, R = 0.9, P = 0.001; Fig. 3B).
Effect of Spartina’s physical structure on sessile and mobile macroinvertebrate assemblages
Spartina-mimics exerted an effect on the abundance, richness and composition of benthic macroinvertebrate assemblages of the rocky salt marshes (Fig. 4). The mean total abundance of functional groups, as well as the feeding types that they include, was significantly higher in mimics than in control plots (Table 2 and 3, Fig. 4A). The same pattern was observed for taxonomic richness, except for the grazer and suspension feeders, which showed no significant difference between mimics and controls (Table 2 and 3, Fig. 4A). The invertebrate abundance and richness in Spartina-mimics were not significantly different from the natural-Spartina plots (Table 2, Fig. 4A) except for the richness of mobile species and the abundance of the grazer Siphonaria lessoni, which showed higher values in the mimics (Fig. 4A). Differences in taxonomic richness of mobile fauna between mimics and natural-Spartina plots were caused primarily by the complete absence of the amphipods Melita palmata and Monocorophium insidiosum in mimics, as well as by the absence of some rare polychaetes in natural-Spartina plots. The MDS plots and ANOSIM analysis showed that the composition of mobile macroinvertebrates was significantly dissimilar between treatments plots (ANOSIM, Spartina-mimics vs. control, R = 0.8, P = 0.001; Spartina-mimics vs. natural-Spartina, R = 0.9, P = 0.001; control vs. natural-Spartina, R = 0.9, P = 0.001, Fig. 4B). On the other hand, the composition of sessile assemblages did not show differences between Spartina-mimics and natural-Spartina (R = 0.1, P ≥ 0.05; Fig. 4B), but differed between the control and Spartina-mimics (R = 0.8, P = 0.001; Fig. 4B) and the control and natural-Spartina (R = 0.9, P = 0.001; Fig. 4B).

A Means and SD for abundances and richness of sessile and mobile taxa and of each feeding type and B MDS ordinations comparing macroinvertebrate composition in Spartina-mimic (art, black squares), control (cont, white circles) and natural-Spartina (nat, cross squares) treatments. Treatments with different letters resulted in significant differences (Tukey unequal N HSD tests or multiple comparisons of mean ranks, p < 0.05). Feeding types are indicated as suspension feeder (SF), deposit-detritus feeder (DDF), predator (P) and grazer (G)
When data of both experiments was pooled in a single data matrix and MDS and ANOSIM analysis were performed, we found that the controls and natural-Spartina of both experiments did not significantly differ (ANOSIM, mobile assemblages: control experiment 1 vs. control experiment 2, R = 0.3, P = 0.01; natural-Spartina experiment 1 vs. natural-Spartina experiment 2, R = 0.2, P = 0.006; sessile assemblages: control experiment 1 vs. control experiment 2, R = 0.5, P = 0.003; natural-Spartina experiment 1 vs. natural-Spartina experiment 2, R = 0.2, P = 0.02; Fig. 5). Therefore, this lack of differences allows us to go a step further in analyzing the results obtained, which indicate that the composition of mobile and sessile macroinvertebrates assemblages did not significantly differ between transplants and mimics Spartina (ANOSIM, mobile assemblages, R = 0.4, P = 0.001; sessile assemblages, R = 0.1, P = 0.1; Fig. 5).

MDS ordinations comparing macroinvertebrate composition in Spartina-transplants (black diamonds), Spartina-mimic (black squares), control experiment 1 (white circles), control experiment 2 (white diamonds), natural-Spartina experiment 1 (cross squares) and natural-Spartina experiment 2 (crosses) treatments
Discussion
Our results show that the cordgrass Spartina densiflora shape the macroinvertebrate assemblages of the rocky marshes where it dominates. The experimental plant mimics combined with the transplant of living plants allowed us to show that this ecological effect is caused by the plants’ physical structure. Indeed, the finding that in the brief period of 1 year, macroinvertebrates responded similarly to both biotic (transplants) and abiotic (mimic) structures strongly suggests that the chemical and biological properties of the plants are not essential to the initial process structuring these faunal assemblages. These results are consistent with those found for a variety of other organisms (e.g., coralline turf: Kelaher 2002; mussel beds: Crooks and Khim 1999; Palomo et al. 2007; polychaete tubes: Callaway 2003; seagrasses: Lee et al. 2001), which have a major influence on the associated assemblages mediated by the physical structure they provide.
The mechanisms by which the physical structure of plants can have a significant effect on communities are many. For instance, in intertidal environments, heat and dehydration stress are two important forces affecting the distribution and abundance of fauna (Tomanek and Helmuth 2002; Harley and Helmuth 2003); however, the shelter supplied by plants can attenuate these stressful conditions (Bortolus et al. 2002; Canepuccia et al. 2007) and reduce predation pressure (Ambrose and Irlandi 1992; Bortolus et al. 2002). In addition, standing structures like cordgrasses and seagrasses modify their local hydrodynamic environment, reducing current velocities and altering turbulent structure in and around the canopy (Fonseca and Koeh 2006), minimizing faunal removal (Bouma et al. 2007), favoring the entrapment and settlement of water-borne fauna and suspended sediment, and enhancing the accretion of organic materials that are in turn utilized by filter and deposit feeders (Bouma et al. 2009). Moreover, plant structure serves as surface of settlement of epiphytes which are an important food source for many grazers. Therefore, the kind of structure supplied by the plants is likely to mediate the abundance of primary producers, consumers and their predators (Bologna and Heck 1999; Boström and Mattila 1999; Edgar 1999).
The dominant animal species found among the Spartina stems (either transplanted or mimics) were adults of the little mussels Brachidontes purpuratus and B. rodriguezii, and a small amount (<1%) were newly recruited. This result was likely due to the local dispersal of small groups of mussels by wave action, a common disturbance on intertidal environments (Witman and Suchanek 1984; Hunt and Scheibling 2001). These small groups of adult mussels were able to colonize the experimental plots after lodging there by rapidly attaching their byssal threads. Thus, although they were not able to control the trajectory after the dislodgment, the final product was a massive colonization that made the experimental plots (either mimics or transplants) undistinguishable from their surroundings just a few weeks after the experiments started. This process not only minimizes the death/loss of mature fertile individuals from the local population, but it also optimizes the colonization of the available substrate, even at times when larvae are not dispersing or when conditions do not favor larval post-settlement survival (Hunt and Scheibling 2000). The presence of the control treatment in both experiments contributes to highlight an interesting outcome about colonization rates. Within the period of 1 year, mobile and sessile fauna colonized several times more abundantly transplant and artificial cordgrass than control plots. These results stand out the dynamic character of the community in rocky salt marshes, and the key role of cordgrasses in structuring the macroinvertebrate communities.
When transplanted and artificial cordgrasses were compared to natural-Spartina plots, no differences in assemblages of sessile organisms were found, but there were some differences due to larger abundance and richness of some mobile organisms between transplant and artificial plots. This result may have been influenced by the fact that transplant and artificial plots formed small discrete patches, whereas the natural habitats were sampled from larger patches of more continuous habitat. Small or isolated patches of habitat may enhance local densities and richness (Virnstein and Curran 1986; Wahl 2001; Healey and Hovel 2004). It is possible (although difficult to achieve due to logistic constrains) to evaluate the relative effect of spatial scale by designing similar experiments in which the scale is manipulated by deploying mimic and natural plots increasingly large (Cardoso et al. 2007).
The cordgrass Spartina densiflora has been reported to be aggressively invading a variety of different environments around the world having important consequences on native communities (Bortolus 2006). Within invaded regions, the austral cordgrass is known to exploit primary substrate for native organisms but it is also likely to provide settlement space for sessile macroinvertebrates and habitat for mobile taxa. Given that non-native habitat-forming species are ubiquitous in both marine and terrestrial environments, it is important to improve our knowledge of their effects on their native environments in order to predict effects in the invaded communities.
References
Ambrose WG, Irlandi EA (1992) Height of attachment on seagrass leads to trade-off between growth and survival in the bay scallop Argopecten irradians. Mar Ecol Prog Ser 90:45–51
Barnes M (2000) The use of intertidal barnacle shells. Oceanogr Mar Biol Ann Rev 38:157–187
Bertness MD (1984) Ribbed mussels and Spartina alterniflora production in a New England salt marsh. Ecology 65:1794–1807
Bologna PAX, Heck KL (1999) Macrofaunal associations with seagrass epiphytes. Relative importance of trophic and structural characteristics. J Exp Mar Biol Ecol 242:21–39
Bortolus A (2006) The austral cordgrass Spartina densiflora Brong.: its taxonomy, biogeography and natural history. J Biog 33:158–168
Bortolus A (2008) Error cascades in the biological sciences: the unwanted consequences of using bad taxonomy in ecology. Ambio 37:114–118
Bortolus A (2010) Marismas Patagónicas: las últimas de Sudamérica. Ciencia Hoy 19:10–15
Bortolus A, Iribarne OO (1999) The effect of the Southwestern Atlantic burrowing crab Chasmagnathus granulata on a Spartina salt-marsh. Mar Ecol Prog Ser 178:79–88
Bortolus A, Schwindt E, Iribarne OO (2002) Positive plant-animal interaction in the high marsh of an Argentinean coastal lagoon. Ecology 83:733–742
Bortolus A, Laterra P, Iribarne OO (2004) Crab-mediated phenotypic changes in Spartina densiflora Brong. Est Coast Shelf Sci 59:97–107
Bortolus A, Schwindt E, Bouza PJ, Idaszkin YL (2009) A characterization of Patagonian salt marsh. Wetlands 29:772–780
Bos AR, Bouma TJ, de Kort GLJ, van Katwijk MM (2007) Ecosystem engineering by annual intertidal seagrass beds: sediment accretion and modification. Est Coast Shelf Sci 74:344–348
Boström C, Mattila J (1999) The relative importance of food and shelter for seagrass-associated invertebrates: a latitudinal comparison of habitat choice by isopod grazers. Oecologia 120:162–170
Bouma TJ, van Duren LA, Temmerman S, Claverie T, Blanco-Garcia A, Ysebaert T, Herman PMJ (2007) Spatial flow and sedimentation patterns within patches of epibenthic structures: combining field, flume and modelling experiments. Cont Shelf Res 27:1020–1045
Bouma TJ, Olenin S, Reise K, Ysebaert T (2009) Ecosystem engineering and biodiversity in coastal sediments: posing hypotheses. Helgol Mar Res 63:95–106
Callaway R (2003) Long-term effects of imitation polychaete tubes on benthic fauna: they anchor Mytilus edulis (L.) banks. J Exp Mar Biol Ecol 283:115–132
Canepuccia AD, Escapa M, Daleo P, Alberti J, Botto F, Iribarne OO (2007) Positive interactions of the smooth cordgrass Spartina alterniflora on the mud snail Heleobia australis, in South Western Atlantic salt marshes. J Exp Mar Biol Ecol 353:180–190
Canion CR, Heck KL Jr (2009) Effect of habitat complexity on predation success: re-evaluating the current paradigm in seagrass beds. Mar Ecol Prog Ser 393:37–46
Caraco N, Cole J, Findlay S, Wigan C (2006) Vascular plants as engineers of oxygen in aquatic systems. Bioscience 56:219–225
Cardoso PG, Raffaelli D, Pardal MA (2007) Seagrass beds and intertidal invertebrates: an experimental test of the role of habitat structure. Hydrobiologia 575:221–230
Clarke KR, Warwick RM (1994) Change in marine communities: an approach to statistical analysis and interpretation. Natural Environment Research Council, Plymouth
Crain CM, Bertness MD (2006) Ecosystem engineering across environmental gradients: implications for conservation and management. Bioscience 56:211–218
Crooks JA, Khim HS (1999) Architectural vs. biological effects of habitat-altering exotic mussel, Musculita senhousia. J Exp Mar Biol Ecol 240:53–75
Duffy JE, Hay ME (1994) Herbivore resistance to seaweed chemical defense: the roles of mobility and predation risk. Ecology 75:1304–1319
Edgar GJ (1999) Experimental analysis of structural versus trophic importance of seagrass beds. I Effects on macrofaunal and meiofaunal invertebrates. Vie Milieu 49:239–248
Fonseca MS, Koeh MAR (2006) Flow in seagrass canopies: the influence of patch width. Est Coast Shelf Sci 67:1–9
Gosselin LA, Chia FS (1995) Distribution and dispersal of early juvenile snails: effectiveness of intertidal microhabitats as refuges and food sources. Mar Ecol Prog Ser 128:213–223
Gribben PE, Wright JT (2006) Invasive seaweed enhances recruitment of a native bivalve: roles of refuge from predation and the habitat choice of recruits. Mar Ecol Prog Ser 318:177–185
Hammond H, Griffiths CL (2004) Influence of wave exposure on South African mussel beds and their associated infaunal communities. Mar Biol 144:547–552
Harley CDG, Helmuth BST (2003) Local- and regional-scale effects of wave exposure, thermal stress, and absolute versus effective shore level on patterns of intertidal zonation. Limnol Oceanogr 48:1498–1508
Healey D, Hovel KA (2004) Seagrass bed patchiness: effects on epifaunal communities in San Diego Bay, USA. J Exp Mar Biol Eco 313:155–174
Henninger TO, Froneman PW, Richoux NB, Hodgson AN (2009) The role of macrophytes as a refuge and food source for the estuarine isopod Exosphaeroma hylocoetes (Barnard, 1940). Est Coast Shelf Sci 82:285–293
Hunt HL, Scheibling RE (2000) Movement and wave dislodgement of mussels on a wave-exposed rocky shore. Veliger 45:273–277
Hunt HL, Scheibling RE (2001) Predicting wave dislodgment of mussels: variation in attachment strength with body size, habitat, and season. Mar Ecol Prog Ser 213:157–164
Jones CG, Lawton JH, Shachak M (1997) Positive and negative effects of organisms as physical ecosystem engineers. Ecology 78:1946–1957
Kelaher BP (2002) Influence of physical characteristics of coralline turf on associated macrofaunal assemblages. Mar Ecol Prog Ser 232:141–148
Lee SY, Fong CW, Wu RSS (2001) The effects of seagrass (Zostera japonica) canopy structure on associated fauna: a study using artificial seagrass units and sampling of natural beds. J Exp Mar Biol Ecol 259:23–50
Minello TJ, Zimmerman RJ (1983) Fish predation on juvenile brown shrimp Penaeus aztecus Ives: the effect of simulated Spartina structure on predation rate. J Exp Mar Biol Ecol 72:211–231
Palomo MG, People J, Chapman MG, Underwood AJ (2007) Separating the effects of physical and biological aspects of mussel beds on their associated assemblages. Mar Ecol Prog Ser 344:131–142
Pawlik JR (1992) Chemical ecology of the settlement of benthic marine invertebrates. Oceanogr Mar Biol Ann Rev 30:273–335
Schwindt E, Bortolus A, Iribarne OO (2001) Invasion of a reef-builder polychaete: direct and indirect impacts on the native benthic community structure. Biol Inv 3:137–149
Schwindt E, Iribarne OO, Isla FI (2004) Physical effects of an invading reef-building polychaetes on an Argentinean estuarine environment. Est Coast Shelf Sci 59:109–120
Sellheim K, Stachowicz JJ, Coates RC (2010) Effects of a nonnative habitat-forming species on mobile and sessile epifaunal communities. Mar Ecol Prog Ser 398:69–80
Silliman BR, Bortolus A (2003) Underestimation of Spartina alterniflora production in Western Atlantic salt marshes. Oikos 143:549–555
Sueiro MC, Bortolus A, Schwindt E (2010) Habitat complexity and community composition: relationships between different ecosystem engineers and the associated macroinvertebrate assemblages. Helgol Mar Res. doi:10.1007/s10152-010-0236-x
Terrados J, Duarte CM (1999) Experimental evidence of reduced particle resuspension within a seagrass (Posidonia oceanica L.) meadow. J Exp Mar Biol Ecol 243:45–53
Tomanek L, Helmuth B (2002) Physiological ecology of rocky intertidal organisms: a synergy of concepts. Integ Comp Biol 42:771–775
Vera F, Gutiérrez JL, Ribeiro PD (2009) Aerial and detritus production of the cordgrass Spartina densiflora in a southwestern Atlantic salt marsh. Botany 87:482–491
Virnstein RW, Curran MC (1986) Colonization of artificial seagrass versus time and distance from source. Mar Ecol Prog Ser 29:279–288
Wahl M (1989) Marine epibiosis. I. Fouling and antifouling: some basic aspects. Mar Ecol Prog Ser 58:175–189
Wahl M (2001) Small scale variability of benthic assemblages: biogenic neighborhood effects. J Exp Mar Biol Ecol 258:101–114
Witman JD, Suchanek TH (1984) Mussels in flow: drag and dislodgement by epizoans. Mar Ecol Prog Ser 16:259–268
Zar JH (1999) Biostatistical analysis. Prentice Hall, New Jersey
Acknowledgments
We are particularly grateful to J. Gerwein (State Coastal Conservancy, USA) for making exhaustive revision of the language on an early version of this paper, and for supplying useful comments as well. We are also enormously grateful to G. Alonso (Museo Argentino de Ciencias Naturales), B. Doti and D. Roccatagliata (UBA) and to E. Diez (CENPAT) for assisting us with expert taxonomic identifications of amphipods, isopods, tanaids and polychaetes, respectively (sample specimens were deposited in the CENPAT invertebrate collection), and to M. Cuestas, Y. Idaszkin, M.M. Mendez, V. Savoya and P. Raffo for helping us with the field work. CONICET, ANPCyT-FONCyT (PICT No. 2206 to AB and PICT No. 20621 to ES) supplied financial support. This work is part of the doctoral thesis of the first author at Universidad de Buenos Aires (UBA), Argentina. We also thank the comments and constructive criticisms of three anonymous reviewers who greatly helped to improve the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Piet Spaak
Rights and permissions
About this article
Cite this article
Sueiro, M.C., Bortolus, A. & Schwindt, E. The role of the physical structure of Spartina densiflora Brong. in structuring macroinvertebrate assemblages. Aquat Ecol 46, 25–36 (2012). https://doi.org/10.1007/s10452-011-9379-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10452-011-9379-3