
Abstract
Primary productivity, community respiration, chlorophyll a concentration, phytoplankton species composition, and environmental factors were compared in the Yolo Bypass floodplain and adjacent Sacramento River in order to determine if passage of Sacramento River through floodplain habitat enhanced the quantity and quality of phytoplankton carbon available to the aquatic food web and how primary productivity and phytoplankton species composition in these habitats were affected by environmental conditions during the flood season. Greater net primary productivity of Sacramento River water in the floodplain than the main river channel was associated with more frequent autotrophy and a higher P:R ratio, chlorophyll a concentration, and phytoplankton growth efficiency (αB). Total irradiance and water temperature in the euphotic zone were positively correlated with net primary productivity in winter and early spring but negatively correlated with net primary productivity in the late spring and early summer in the floodplain. In contrast, net primary productivity was correlated with chlorophyll a concentration and streamflow in the Sacramento River. The flood pulse cycle was important for floodplain production because it facilitated the accumulation of chlorophyll a and wide diameter diatom and green algal cells during the drain phase. High chlorophyll a concentration and diatom and green algal biomass enabled the floodplain to export 14–37% of the combined floodplain plus river load of total, diatom and green algal biomass and wide diameter cells to the estuary downstream, even though it had only 3% of the river streamflow. The study suggested the quantity and quality of riverine phytoplankton biomass available to the aquatic food web could be enhanced by passing river water through a floodplain during the flood season.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Floodplains are considered to be important for aquatic production worldwide because they are a source of phytoplankton carbon to riverine food webs (Junk et al. 1989). Net primary productivity is high in floodplains where a high ratio of the euphotic zone depth to mixing zone depth reduces the loss of gross primary productivity to respiration (Heip et al. 1995). The shallow water depth and long residence time in floodplains also facilitates sedimentation of suspended solids that increase the total irradiance available for phytoplankton growth in the water column (Tockner et al. 1999). Long residence time in floodplains increases the availability of phytoplankton biomass to the food web by accumulating phytoplankton cells, particularly during the drain phase of the flood pulse cycle (Kiss 1987; Lewis 1988; Van den Brink et al. 1993; Hein et al. 1999).
Floodplains may also be a good source of high-quality phytoplankton for the lower food web because they contain abundant diatom and green algal species compared with adjacent rivers (Kiss 1987). The wide spherical diameter and high carbon content of these algal cells provide high-quality food for zooplankton at the base of the aquatic food web locally and their transport enhances riverine production regionally (Hansen et al. 1994; Lewis et al. 2001; Keckeis et al. 2003). Despite the potential importance of floodplains to estuarine food web production, little is known about the relative magnitude and controlling mechanisms associated with primary productivity and respiration in floodplain versus riverine habitat and the potential contribution of phytoplankton biomass in floodplains to the aquatic food web in adjoining estuaries.
Primary productivity in the Yolo Bypass floodplain (Yolo Bypass) is hypothesized to be a net source of phytoplankton carbon to the aquatic food web in San Francisco Estuary (SFE) (Jassby et al. 2002; Sommer et al. 2001a). Net primary productivity in the freshwater tidal channels of SFE is low because high suspended sediments reduces total irradiance in the water column and low euphotic zone depth to mixing depth ratios increase carbon loss to respiration (Jassby et al. 2002). In contrast, field measurements suggest shallow-water habitats along the margin of the marine bays and within the interior of the freshwater tidal reaches of SFE are highly productive and a potential source of phytoplankton carbon to the estuary (Cole and Cloern 1984; Caffrey et al. 1998; Lucas et al. 2002). Modeling studies also indicated Yolo Bypass is a net source of organic carbon to the estuary because its shallow depth enhances carbon production (Jassby and Cloern 2000). Field measurements confirmed the greater chl a concentration per unit volume in Yolo Bypass than the adjacent Sacramento River (Sommer et al. 2004b). High chl a concentration in the floodplain just after flooding also suggested riverine–floodplain lateral exchange contributed directly to chl a in the floodplain (Schemel et al. 2004). A predictive model indicated high chl a concentration in the Yolo Bypass is produced by long hydraulic residence time, high surface to volume ratio and high water temperature (Sommer et al. 2004b). It was unknown if the greater chl a in Yolo Bypass compared with the adjacent Sacramento River channel was the result of higher phytoplankton growth rate or biomass accumulation. Comparable field measurements of primary productivity and respiration in the floodplain and river are lacking.
The purpose of this research was to determine if passage of Sacramento River water through the Yolo Bypass increases the quantity and quality of phytoplankton carbon available for bottom-up food web production, how environmental conditions affect the quantity and quality of phytoplankton carbon in the floodplain and river and if the floodplain contributes to the downstream load of phytoplankton carbon. Such information is needed to assess the potential use of floodplains to enhance estuarine fish production in SFE. Since juvenile native fish density is high and accompanied by high fish growth rate in Yolo Bypass, floodplain habitat is thought to be important for native fish production in SFE (Sommer et al. 2001b). It is hypothesized that the floodplain enhances juvenile fish production by stimulating bottom-up production through the growth of phytoplankton carbon. The importance of phytoplankton carbon to fishery production is supported by correlation between total and diatom phytoplankton biomass and zooplankton and Neomysis shrimp biomass in the freshwater tidal and brackish water reaches of SFE (Orsi and Mecum 1996; Jassby et al. 1995; Lehman 1992, 2004).
Materials and methods
Study area
Yolo Bypass is a managed 240 km2 floodplain that is flooded periodically between January and June when high discharge caused by high precipitation and snowmelt runoff is diverted from the Sacramento River at Fremont Weir (Fig. 1). Sacramento River is a large deep river that drains a 70,000 km2 watershed in California and has a mean annual discharge of 800 m3 s−1. The periodic flooding (flood phase) and subsequent drainage (drain phase) of Yolo Bypass creates a flood pulse cycle with flood level defined as the water depth at 3 m or above. The floodplain remains dry during the late summer, fall, and early winter when it is used for agriculture.

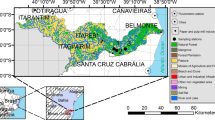
Map showing sampling stations at Lisbon Weir (YB) in the Yolo Bypass floodplain (shaded area) and Sherwood Harbor (SR) on the Sacramento River
Physical, chemical, and biological measurements were made at Lisbon Weir (YB) in a secondary channel that drains the eastern boundary of Yolo Bypass and diverts water from the floodplain to the estuary downstream. Sampling at YB provided a unique opportunity to determine the influence of floodplain habitat on Sacramento River water because only Sacramento River water remains near the eastern boundary of Yolo Bypass and drains eastward to YB (Sommer et al. 2001a). Yolo Bypass demonstrates an extreme case of hydrologic banding in which shallow water depth, low gradient, and low roughness in the floodplain prevent the four major tributaries that enter Yolo Bypass from mixing over the 61 km length of the floodplain (Sommer et al. 2001a; Schemel et al. 2004); this phenomenon has characterized Yolo Bypass for over 30 years (T. Sommer, personal communication). Spatial variability along the eastern boundary of the floodplain was not measured during this study due to poor access and high discharge but it was presumed to be low. Land preparation for summer dry land farming reduces topographic variation and water quality is homogeneous along the eastern boundary (Schemel et al. 2004; Schemel and Cox 2007). Between January and June 2003, median discharge was 23 m−3s−1 and median depth was 2.25 m at YB. Models developed for Yolo Bypass indicated that under these conditions residence time on the floodplain at YB was 9 days (Sommer et al. 2004b).
The primary productivity of Sacramento River water that passed through Yolo Bypass at YB was compared with Sacramento River water in the main channel of the river at Sherwood Harbor (SR) located directly east of YB (Fig. 1). The Sacramento River is one of two major rivers that feed SFE and drain 40% of California. Median discharge during the study period was 800 m−3 s−1, median depth was 6 m, and average residence time was 2 days at SR.
Field methods and materials
Net primary productivity and community respiration (respiration; phytoplankton plus bacterial) were computed from the change in dissolved oxygen concentration in water samples from in situ light and dark bottle incubations (Vollenweider 1974). Water samples for primary productivity measurements were obtained biweekly between January and June with a van Dorn water sampler; environmental conditions precluded primary productivity measurements only on 2 days at YB. Water samples were collected at 0.3 m depth which provided a representative sample of the water quality and phytoplankton in the water column based on chl a concentration, water temperature, pH, and specific conductance in vertical profiles made with a YSI 6600 water quality sonde (Yellow Springs Instruments, Yellow Springs, OH, USA). Primary productivity and respiration were measured using replicate 300 ml borosilicate glass bottles which were overflowed three times to remove gas bubbles and stoppered in the dark. The variation in net productivity with irradiance was obtained by wrapping some of the bottles in polypropylene screens and the resulting bottle array produced a light gradient of 0%, 20%, 30%, 50%, and 100% of the ambient light in the water column. Prepared bottles were incubated horizontally at 0.3–0.45 m in the water column at each station. This shallow depth was selected in order to expose the incubation bottles to light in the euphotic zone (depth of 1% surface irradiance) which ranged between 0.6 and 1.4 m in YB and 1.2 and 3.3 m in SR. Bottles were incubated for 24 h in order to include the diel variation in solar irradiance. Net primary productivity and respiration were determined from the change in dissolved oxygen concentration measured immediately after incubation using a YSI 5000 dissolved oxygen meter combined with a 5010 BOD probe and its attached stirrer (Yellow Springs Instruments, Yellow Springs, OH, USA). Accuracy of the dissolved oxygen meter was verified using Winkler titration (APHA et al. 1998). Oxygen units for production and respiration were converted to carbon units for discussion using a photosynthetic quotient of 1.25 (mol O2 per mol C produced or respired; Vollenweider 1974).
Incubations were increased three times in February and early March to 72 h in order to obtain significant differences of at least 0.20 mg l−1 in dissolved oxygen concentration over the incubation period in both light and dark bottles (Vollenweider 1974). Longer incubations were needed because of an extended period of dense ground fog (Tule Fog) that occurs during the winter in Sacramento Valley. These longer incubations probably had minimal impact on the growth of phytoplankton communities at this time of year when water temperature, surface irradiance, phytoplankton biomass, growth rate, and respiration were low and nutrient concentration was high.
Gross primary productivity (mg C m−3 day−1) was estimated for 0.15-m depth intervals from photosynthetic parameters, irradiance (I, mol quanta m−2 s−1) at depth and chl a concentration using a photosynthesis–irradiance (P–I) model that allowed for photoinhibition: GPP = chl a × P m B × (1 − exp((−αB × I)/P m B)) × exp((−β × I)/P m B) (Platt and Sathyendranath 1990). Photosynthetic parameters included the photosynthetic capacity from the chl a-specific light saturated rate of photosynthesis (P m B , mg C (mg chl a)−1 d−1), the photosynthetic efficiency from the chl a-specific initial slope (αB; mg C (mg chl a)−1 (mol quanta m−2)−1), the photoinhibition parameter from the chl a-specific negative slope of the P–I curve above light saturation (βB, mg C (mg chl a)−1 (mol quanta m−2)−1). Areal gross primary productivity in the euphotic zone and water column was computed by integrating gross primary productivity values over depth using the trapezoidal rule. Gross primary productivity values for the P–I curve were obtained by adding net primary productivity and respiration.
Photosynthetically active surface irradiance (PAR) was measured in Langleys before April from daily irradiance at Davis, CA (http://www.ipm.ucdavis.edu; Fig. 1) and after April by quanta at 15-min intervals using a LICOR 190SA quantum sensor (LI-COR, Inc., Lincoln, NE, USA) at YB. Langleys were converted to quanta using linear correlation (r 2 = 0.91; p < 0.01). Irradiance in the water column was measured at 0.15-m intervals by vertical profiles of a LICOR 193SA spherical quantum sensor (LI-COR, Inc., Lincoln, NE, USA) and total irradiance in the euphotic zone was computed by integration over depth. Total depth varied with precipitation and snowmelt runoff and was determined hourly from stage recorders at both YB an SR. Discharge was computed from hourly depth and discharge regressions at SR and measured by an acoustic Doppler current profiler at YB. Water temperature and specific conductance were measured, respectively, with an Onset continuous temperature logger (ONSET Computer Corporation, Bourne, MA, USA) and a YSI Model 85 (Yellow Springs Instruments, Yellow Springs, OH, USA) temperature probe at both stations.
Water samples for water quality analysis were collected at 0.3 m depth using a Van Dorn water sampler, stored at 4°C and prepared for laboratory analyses within 4 h of collection. Water samples for chl a and phaeophytin concentration were filtered through APFF glass fiber filters (Millipore Corporation, Billerica, MA, USA). Filters were preserved with 1% magnesium carbonate and frozen until analysis. Pigments on the filters were extracted in 90% acetone and analyzed for chl a (corrected for phaeophytin) and phaeophytin using spectrophotometry (method 10200H, APHA et al. 1998). Water samples for soluble reactive phosphorus, nitrate and ammonium were filtered through 0.45 μm pore size Millipore HATF04700 filters (Millipore Corporation, Billerica, MA, USA) and along with raw water samples for total phosphorus were analyzed for nutrient concentration by colorimetric techniques (US EPA 1983). Silica concentration was determined by the molybdate blue method (USGS 1985). Water samples for identification and enumeration phytoplankton species and measurement of phytoplankton cell dimension were preserved and stained with Lugol’s iodine solution (Van Waters and Rogers Scientific Products, Brisbane, CA, USA). Phytoplankton was counted and identified to at least the genus level at 700× magnification using the inverted microscope technique (Utermöhl 1958). This magnification allowed clear identification of phytoplankton cells >6 μm in diameter. Phytoplankton carbon (biomass) was calculated from volume based on cell dimensions using simple geometrical shapes and corrected for the small plasma volume in diatoms by equations in Menden-Deuer and Lessard (2000). These computed cell volumes were used to calculate estimated spherical diameter of each cell (Hansen et al. 1994).
Statistical methods
In order to obtain the most robust results and to adjust for the lack of a normal distribution associated with small sample size, non-parameteric statistics were used for the statistical analysis of the data. Median and 25th and 75th percentiles were used to describe the central distribution of each variable. Single comparison tests were made using the Mann–Whitney U test. Linear correlation was computed using Spearman rank-order correlation (r s). All statistical analyses were computed using Statistical Analysis System (SAS 2004) software (SAS Institute Inc., Cary, NC, USA).
Results
Primary productivity
Areal gross and net primary productivity of the water column and euphotic zone were greater from winter to summer at YB than SR and were associated with a higher chl a-specific net primary productivity and αB; there was no difference in P m B or Bβ (Table 1). Greater areal net primary productivity of the water column and euphotic zone at YB was primarily caused by higher gross primary productivity because both total and chl a-specific respiration did not differ between YB and SR. The total water column respiration at YB and SR was unexpectedly similar because of the combined influence of chl a concentration and depth. Respiration associated with the factor of 4 higher (p < 0.01) chl a concentration in the shallow 2 m water column at YB did not differ significantly from the respiration produced by low chl a concentration in the deep 6 m water column at SR (Table 1). The greater areal net primary productivity of the water column at YB was also associated with nearly a factor of 2 greater euphotic zone depth to total depth ratio (median 0.60 and 0.37 at YB and SR, respectively).
Areal net primary productivity of the water column at YB alternated between autotrophy and heterotrophy and was highest in early spring (Fig. 2). Areal net primary productivity was positive and significantly greater (p < 0.01) in March and June compared with negative values in January, February, April, and May. Autotrophy of the water column in March and June was produced by greater (p < 0.05) chl a-specific net primary productivity (median 6.8 mg C (mg chl a)−1 d−1 for March and June and −8.6 mg C (mg chl a)−1 d−1 for January, February, April, and May). Heterotrophy of the water column in January, February, April, and May was produced by a factor of 2 lower (p < 0.01) P:R ratio, but there was no significant increase in the chl a-specific respiration. A somewhat higher percentage of phaeophytin to total pigment (chl a plus phaeophytin) concentration in April through June (31%) compared with March (19%) suggested the high respiration in the late spring varied with a seasonal decline in phytoplankton health, but the difference was not significant. In contrast with YB, areal net productivity of the water column at SR was consistently near zero or negative and did not increase significantly in early and or late spring (Fig. 2).

Daily mean (bar) and standard deviation (vertical line) of areal gross and net primary productivity and community respiration in the water column between January and June 2003 in the Yolo Bypass floodplain and the Sacramento River
Phytoplankton biomass and community composition
Chl a concentration was greater (p < 0.05) on a volumetric basis but not on an areal basis at YB than SR (median volumetric 9.9 and 2.4 μg chl a l−1; median areal 20.4 and 15.7 mg chl a m−2, respectively; Fig. 3). The factor of 4 greater chl a concentration at YB was associated with a 50% greater percentage (p < 0.01) of diatom and green algae compared with SR where cryptophytes (p < 0.01) were dominant (Fig. 4). Diatoms also had the widest estimated spherical diameter (p < 0.05) among phytoplankton groups and their abundance contributed to the wider (p < 0.01) estimated spherical diameter of phytoplankton at YB than SR (median 7 μm; range 2–46 μm and median 6 μm; range 3–22 μm, respectively). The greater spherical diameter of cells at YB was produced by the presence of a greater percentage of cells with spherical diameter wider than 10 μm (median 20% in YB and 0% in SR). A large portion of the 40% greater diatom and green algal biomass at YB was produced by a difference in the distribution of biomass among species and not a difference in species composition between stations. About 45% of the diatom and green algal carbon at YB was produced by species in common with SR, including the diatoms, Cyclotella sp. (11%), Synedra sp. (6%) and Thalassiosira eccentrica (1%) and the green algae Ankistrodesmus falcatus (6%), Chlamydomonas sp. (6%), and Chlorella sp. (5%).

Daily mean (bar) and standard deviation (vertical line) of chlorophyll a concentration and areal chlorophyll a in the water column for Yolo Bypass floodplain and the Sacramento River between January and June 2003

Median (bar) and 25th and 75th percentiles (vertical line) of percent carbon among phytoplankton groups in Yolo Bypass floodplain and the Sacramento River between January and June 2003
Environmental factors
Net primary productivity of the water column was both positively and negatively influenced by light in the euphotic zone at YB. Net primary productivity in the euphotic zone was limited by light in the late winter and early spring between January and March. During this period the depth of P m B (Z k) and the compensation depth (Z c) where net primary productivity is zero were at or near the surface (0 and 0.7 m, respectively) due to low surface irradiance and high light attenuation in the water column from suspended sediment (Fig. 5). Net primary productivity in the euphotic zone did not increase to high values as expected in response to the seasonal increase in surface irradiance after March (Fig. 6). Instead, net primary productivity was relatively low and associated with a decrease (p < 0.05) in the median P:R ratio from 1.4 to 0.8 after March. The lower P:R ratio in the euphotic zone after March was partially caused by higher respiration produced by an increase in the depth of the euphotic zone from 0.9 to 1.2 m. It was not produced by an increase in the chl a-specific respiration. Further, the increased respiration in the euphotic zone in the late spring may be an underestimate because incubations were not at the surface.

Daily surface irradiance, total irradiance in the euphotic zone, and water temperature in the Yolo Bypass floodplain and Sacramento River and water depth in Yolo Bypass between January and June 2003

Daily areal net primary productivity in the euphotic zone, water temperature, and daily surface irradiance in the Yolo Bypass floodplain and the Sacramento River
The low P:R ratio after March was associated with a decrease in P m B. P m B decreased with irradiance and was negatively correlated with surface irradiance (r s = −0.86; p < 0.01) and total irradiance in the euphotic zone (r s = −0.68; p < 0.05) after January at YB (Fig. 5). The decrease in the P m B between March and April alone from 93.2- to 67 mg C (mg chl a)−1 d−1 in combination with a downward shift in Z k from the surface to 0.29 m produced a 50% decrease in chl a-specific net productivity in the euphotic zone (median 23 mg C (mg chl a)−1 d−1 in March to 12 mg C (mg chl a)−1 d−1). The decrease in P m B after March was not associated with a shift in species composition.
Relatively low net primary productivity and high respiration in the euphotic zone after March coincided with an increase in water temperature above 17°C at YB (Fig. 6). Both total and chl a-specific respiration were positively correlated with water temperature after January (r s = 0.95, p < 0.01; r s = 0.78, p < 0.05, respectively) and contrasted with P m B which was negatively correlated with water temperature after January (r s = −0.89; p < 0.01). A high positive correlation between water temperature and respiration for all months (r s = 0.93, p < 0.01) and chl a-specific respiration after January (r s = 0.67, p < 0.05) suggested high water temperature contributed to respiration in the light for the euphotic zone. Water temperature and chl a-specific respiration were positively correlated for both stations despite the greater (p < 0.01) water temperature at YB than SR (median 15.9°C for YB and 13.1°C for SR; Fig. 5).
Nutrient concentration probably had little impact on primary productivity at YB because the nutrients were usually not limiting. Concentrations of dissolved inorganic nitrogen, soluble reactive phosphorus, and silica were 0.12–0.99 mg N l−1, 0.02–0.42 mg P l−1, and 8.10–15.90 mg Si l−1, respectively. These concentrations were above the limiting values for dissolved inorganic nitrogen, soluble reactive phosphorus, and silica of 0.07 mg N l−1, 0.03 mg P l−1, and 0.15 mg l−1 SiO2, respectively (Jassby 2005). Soluble reactive phosphorus concentration was limiting only once on May 6 (0.02 mg P l−1).
Unlike YB, water temperature and total irradiance in the euphotic zone were not significantly correlated with primary productivity at SR (Fig. 6). Instead, gross primary productivity in the euphotic zone was positively correlated with chl a concentration (r s = 0.76; p < 0.05) and streamflow (r s = −0.62; p < 0.05). Low (p < 0.01) chl a concentration at SR led to low net primary productivity despite the greater total irradiance and depth of the euphotic zone at SR than YB resulting from low suspended sediment concentration (median extinction coefficient of 2.2 and 3.5 m−1 and median euphotic zone depth of 2.0 and 1.1 m for SR and YB, respectively; Fig. 5). Like YB, concentrations of the major dissolved nutrients inorganic nitrogen, soluble reactive phosphorus and silica (0.09–0.26 mg N l−1, 0.02–0.04 mg P l−1, and 16.00–19.10 mg Si l−1, respectively) were not limiting at SR (Jassby 2005).
Flood pulse cycle
The phytoplankton biomass at YB varied with the flood pulse cycle. Both chl a and phaeophytin concentration were at least twice as high during the drain phase of the flood pulse cycle (13.0 and 6.2 μg l−1 for chl a and 6.7 and 2.0 μg l−1 for phaeophytin during the drain and flood, respectively; p < 0.05) and negatively correlated with total depth (r s = −0.81; p < 0.01; r s = −0.64, p < 0.05, respectively; Fig. 7). The high chl a concentration during the drain phase was not associated with a greater chl a-specific net or gross primary productivity, P m B or αB.

Daily mean (bar) and standard deviation (line) of chlorophyll a concentration and percent carbon of phytoplankton groups present during drain and flood phases in the Yolo Bypass floodplain at Lisbon Weir
Chl a concentration was poorly correlated with physical and chemical conditions during the flood pulse cycle. Neither water temperature, total irradiance in the euphotic zone, soluble reactive phosphorus nor silica concentration were significantly correlated with the phases of the flood pulse cycle (Figs. 5, 7). Nitrate concentration was twice as high during the drain than the flood phase (0.7 and 0.3 mg l−1, respectively; p < 0.05) but concentrations were not limiting (Fig. 7; Jassby 2005).
High chl a concentration in the drain phase was accompanied by a greater percentage of diatom plus green algae (r s = 0.84; p < 0.01) than other phytoplankton groups (Fig. 8). Most of the phytoplankton biomass in the drain phase was associated with the diatoms Achnanthes gibberula, Aulacoseira spp., and Coscinodiscus sp. and the green algae Closterium setaceum, Oocystis sp., and Hyalotheca sp. Green algae also had a significantly wider estimated spherical diameter (p < 0.05) and greater range of values during the drain phase compared with the flood phase (median 5.9 μm, range 36.7–1.8 μm versus 4.6 μm, range 8.9–2.6 μm, respectively). The flood phase was characterized by phytoplankton other than diatom and green algae (p < 0.01). The individual species varied for each flood and combined had a median spherical cell diameter of 6.3 μm. Species which comprised most of the biomass during floods included the cryptophyte Rhodomonas sp. in January, the chrysophyte, Dinobryon sertularia in March, and the bluegreen alga Aphanizomenon flos-aquae in May.

Mean daily (bars) and standard deviation (vertical line) of silica, soluble reactive phosphorus, and nitrate concentration present during drain and flood phases in the Yolo Bypass floodplain at Lisbon Weir
Carbon load
Passing Sacramento River water through Yolo Bypass increased the phytoplankton carbon load to the estuary downstream because of the high concentrations of chl a and diatom and green algal biomass produced in the floodplain. The flux of phytoplankton carbon at YB contributed 14% of the chl a, 14% of the diatom, 31% of the green algae, and 8% of the cryptophyte biomass of the combined YB plus SR load of each constituent to the estuary downstream. The wide spherical diameter of the phytoplankton cells at YB also accounted for 37% of the total estimated spherical diameter in the combined downstream load. The contribution of YB to the downstream load was relatively large considering the streamflow past YB of 23 m−3 s−1 was only 3% of the streamflow at SR of 766 m−3 s−1.
The net carbon load from primary productivity integrated over the water column was heterotrophic over the sampling period for both sampling stations (n = 10); −2,632 kg C d−1 at YB and −58,970 kg C d−1 at SR. The greater carbon load from YB than SR was due to the positive net carbon load from primary productivity in the euphotic zone (2,379 kg C d−1 at YB and −23,089 kg C d−1 at SR). The daily carbon load of the water column at YB was highly variable and ranged many fold from −1,468 kg C d−1 in January to 1,462 kg C d−1 in May. The daily carbon load was even more variable at SR; range −17,653 kg C d−1 in February to 2,838 kg C d−1 in March.
Discussion
Primary productivity
Passage of Sacramento River water across the Yolo Bypass increased net primary productivity of the river water. Turbid, shallow-freshwater habitats are thought to enhance primary productivity because the increased vertical mixing in these habitats exposes phytoplankton cells to surface light more frequently than deep-water habitats (Mallin and Pearl 1992). Net primary productivity in shallow- and deep-water habitats is commonly controlled by the euphotic zone depth to mixed zone depth ratio in estuaries (Cole et al. 1992; Kemp et al. 1997) and is an important factor affecting primary productivity in SFE where nutrients are in excess and net primary productivity is light limited (Jassby et al. 2002). A shallow mixing depth was probably a critical factor affecting the irradiance in the water column available for phytoplankton growth in Yolo Bypass because the concentration of total suspended solids was higher in YB than the river. High irradiance in the water column due to shallow water depth and sedimentation of suspended solids is considered to be a major factor contributing to the greater net primary productivity in floodplains compared with rivers world wide (Unrein 2002; Tockner et al. 1999). Phytoplankton in Yolo Bypass may have further compensated for the high-suspended solids concentration by having a high αB. This may partially explain the dominance of diatom and green algal species in Yolo Bypass because some diatom and green algal species grow more efficiently at low light than other phytoplankton due to lower maintenance respiration at low light intensity (Langdon 1988; Reynolds 1997). A high αB was also associated with increased net primary productivity in shallow-water habitats, but had little effect in deep-water habitats in the Hudson River (Cole et al. 1992).
The factor of 2 higher P:R ratio was largely responsible for the greater areal net primary productivity of the water column and euphotic zone at YB than SR (median 0.74 and 0.32, respectively). The respiration to gross primary productivity ratio similarly increased from shallow- to deep-water habitats when Yolo Bypass, Suisun Bay, and Sacramento River were compared (1-, 2-, and 6-fold, respectively; Rudek and Cloern 1996). Water column respiration measured at YB of 61–786 mg C m−2 d−1 was also less than respiration measured throughout the year in SFE of 200–2,746 mg C m−2 d−1 (Rudek and Cloern 1996). Respiration at YB was most likely due to phytoplankton and not bacteria. Bacteria accounted for only 25% of the decomposition of organic carbon in Yolo Bypass and contributed less to respiration processes in Yolo Bypass than adjacent river channels in long-term respiration studies for SFE (Sobczak et al. 2002).
High and positive net primary productivity of the euphotic zone and water column characterized the floodplain station in early spring. Autotrophy also characterized south San Francisco Bay (see Fig. 1 for location) in spring, but the peak was somewhat later, March and April, versus March in Yolo Bypass (Caffrey et al. 1998). An earlier peak in net primary productivity with distance landward was also measured in 1980 for SFE; April in San Pablo Bay seaward and March in Suisun Bay landward (Cole and Cloern 1984) and supported by modeling studies that suggested peak primary productivity occurs earlier in upstream tributaries to the estuary (Jassby and Cloern 2000). Autotrophy in early spring was similarly measured for the floodplains of the Orinoco River in Venezuela and Danube River in Austria (Lewis 1988; Hein et al. 1999). Autotrophy is generally thought to occur in the early spring for freshwater lakes and rivers because respiration is low at low water temperature and light (Ward and Wetzel 1980).
High variability characterized daily primary productivity at YB and may be characteristic of SFE where gross primary productivity varied by a factor of 20 in the shoals of south San Francisco Bay (Caffrey et al. 1998). Due to this variability, autotrophic conditions may have occurred more frequently at YB than measured by the biweekly sampling program in this study and a higher frequency sampling program might have measured the expected positive net carbon load to the estuary over the sampling period. However, high respiration in the late spring suggested heterotrophy was still a likely outcome. Information on the daily variation in net primary productivity needed to produce a complete picture of the periodicity of autotrophy and net carbon flux for floodplains is generally lacking. Only a few low-frequency measurements are available to access the variability of primary productivity in floodplains of the Orinoco River, Venezuela, and Danube River, Austria (Lewis 1988; Hein et al. 1999). The variation of primary productivity in floodplains is usually inferred from chl a concentration (Tockner et al. 1999; Hein et al. 2004), but these estimates may be poor. Chl a was a poor indicator of net primary productivity at YB.
The influence of spatial variability in the Yolo Bypass on the net primary productivity, respiration, and phytoplankton community composition measured at YB is unknown. Spatial differences in primary productivity and chl a concentration across floodplains are considered to be large and produced by varying residence time, hydrological connectivity, and distance from the main river to floodplain ponds and channels (Tockner et al. 1999; Hein et al. 2004). High variability might be expected in a large floodplain such as Yolo Bypass because chl a concentration varied by as much as 4- to 17-fold over a small 0.36 km2 floodplain of the Cosumnes River just upstream of SFE (Ahearn et al. 2006) and the carbon load from adjacent flooded islands in the freshwater tidal region of SFE varied many fold from 900 to −2,300 kg C d−1 (Lucas et al. 2002). Yet, the influence of spatial variation on net primary productivity at YB may be smaller than expected because Sacramento River water is restricted to a discrete hydrologic band of water near the eastern boundary of the floodplain that drains eastward to the sampling station at YB (Sommer et al. 2001a). Spatial variability of this area is low because it is leveled for dry agriculture during the summer and water quality conditions are primarily influenced by the Sacramento River (Schemel et al. 2004; Schemel and Cox 2007).
Controlling mechanisms
The lower than expected net primary productivity of the water column and euphotic zone at YB in late spring and early summer despite the seasonal increase of surface irradiance suggested high light near the surface reduced primary productivity in the floodplain. Light-inhibited photosynthesis near the surface can be important in shallow-water habitats where vertical mixing frequently exposes algal cells to high surface irradiance (Banaszak and Neale 2001; Neale et al. 1991). This contrasts with deep-water habitats like the Sacramento River where phytoplankton are vertically mixed into the surface layer only occasionally (Heip et al. 1995). Failure to account for photoinhibition increased predicted values of gross primary productivity in the euphotic zone during the late spring by 19–42% at YB when models were based on the P–I curve (Jassby and Platt 1976; Platt and Sathyendranath 1990). Widely different estimates of primary productivity are produced by models with and without photoinhibition and are linked to differences in αB (Frenette et al. 1993). This may be important for YB where αB varied by a factor of 3. Modeling studies for SFE also suggested the growth rate was not as high as expected in response to irradiance in the water column because shallowing of the surface layer alone did not increase phytoplankton bloom potential as expected from the Sverdrup Critical Depth model (Lucas et al. 1998).
Net primary productivity in YB was also influenced by water temperature. The increase of phytoplankton growth rate with water temperature at sub-optimal water temperature (Langdon 1988) contributed to the gradual increase in gross primary productivity over the winter early spring at YB. Yet the seasonal increase in water temperature contributed to the relatively high respiration and resulting low net primary productivity in late spring and early summer. Since most of the respiration in Yolo Bypass was probably due to phytoplankton (Sobczak et al. 2002), phytoplankton species composition could have contributed to the impact of water temperature on net primary productivity at YB. The P:R ratio at a given water temperature varies with species composition (Smith and Kemp 2001) and was high at YB in the early spring when diatoms and green algae were abundant. Some diatom and green algae that occur in early spring have high P:R ratios because their maintenance respiration is low and their αB is high at low water temperature and light compared with other phytoplankton (Langdon 1988; Reynolds 1997). Water column respiration could also have increased in the late spring and early summer when non-diatom biomass was high because non-diatoms are more sensitive to high irradiance and ultraviolet radiation than diatoms (Banaszak and Neale 2001). Net primary productivity in early spring was probably enhanced by the abundance of wide diameter diatom and green algal cells. The P:R ratio was greater when wide diameter phytoplankton cells were abundant in the Chesapeake Bay (Smith and Kemp 2001).
Flood pulse cycle
The flood pulse cycle enhanced accumulation of phytoplankton biomass during the drain phase at YB. The absence of an increase in the chl a-specific gross or net primary productivity, αB or P m B during the drain phase indicated the high chl a and diatom and green algal carbon present during the drain phase was produced by accumulation and not an increase in phytoplankton growth rate. Previous research measured high chl a concentration in Yolo Bypass during the drain phase but it was unknown if this was produced by an accumulation of biomass or an increase in the chl a-specific growth rate (Schemel et al. 2004; Sommer et al. 2004b). Yolo Bypass differed from floodplains of the Orinoco and Danube rivers where P m B or primary productivity were greater during the drain phase (Lewis 1988; Hein et al. 1999). Chl a concentration and primary productivity do not always vary together (Smith and Kemp 2001) and seasonal changes in primary productivity in the Orinoco River floodplain were more attributed to the variation in phytoplankton biomass than growth rate (Lewis 1988). The high phytoplankton biomass during the drain phase at YB was probably influenced by the presence of wide diameter diatom and green algal cells because these cells have high cellular carbon content (Lehman 1996). Green algae were also abundant during the drain phase in the Lower Rhine and Meuse River floodplains (Van den Brink et al. 1994) and both green algae and wide diameter phytoplankton comprised the largest percentage of the total phytoplankton biomass in El Tigre floodplain-lake during the isolation period (drain phase) from the Parana River (Garcia de Emiliani 1997).
Food web impact
It is possible that food web production supported by autotrophy in Yolo Bypass during early spring is important to fishery production in SFE. Feeding and rearing are important habitat uses for many of the 42 fish species found in the YB between winter and early summer (Sommer et al. 2001a). Food web production in early spring may be essential for the survival of native juvenile fish that occur in Yolo Bypass earlier in the spring than exotic species (Sommer et al. 2004a). Accumulation of phytoplankton biomass at YB also suggested Yolo Bypass may contribute to the greater fish growth rate measured in the floodplain by facilitating efficient bottom-up energy transfer through the food web (Sommer et al. 2001b).
The high concentration of diatom and green algal biomass and wide spherical diameter phytoplankton cells at YB provided good quality food for the food web locally and the estuary downstream. Diatoms and green algae have the highest cellular carbon content in the SFE phytoplankton community and the spherical diameter of diatom and green algal cells at YB spanned the range of phytoplankton cells needed to optimize copepod feeding in SFE (Lehman 1996, 2000; Hansen et al. 1994). Laboratory research also suggested phytoplankton was the most biologically available carbon source and produced the highest growth rate for the zooplankter Daphnia magnum in SFE (Mueller Solger et al. 2002; Sobczak et al. 2002). High-quality food may further contribute to the faster growth rate of fish in Yolo Bypass by facilitating more efficient energy transfer within the food web (Sommer et al. 2001b). The importance of phytoplankton community composition to food web production in SFE was supported by the correlation between diatom biomass, mesozooplankton biomass and mysid shrimp biomass throughout upper SFE over a 19-year period (Lehman 2004). Live phytoplankton was also the primary food source for herbivores in the fish food chain for the Amazon and Parana floodplains in South America (Forsberg et al. 1993; Lewis et al. 2001). Riverine food webs may depend on the high-quality organic matter produced in shallow-water habitats like floodplains where autotrophy is common, even though the load may be small compared to within channel and near-channel regions of the river (Lewis 1988; Junk et al. 1989).
Management implications
This study provides direction for future management and research aimed at using floodplains to enhance primary productivity and phytoplankton biomass in SFE. First, divert water into the floodplains early in the spring. Early flooding would enhance net primary productivity by taking advantage of the low water temperature and surface irradiance in early spring that reduces carbon loss to community respiration. Early flooding may also enhance the growth of diatom and green algae with wide spherical diameter and high carbon content that respire less at low light and water temperature than other phytoplankton. Early flooding may be critical for production of native juvenile fish species in SFE which occur earlier in the floodplain than exotic species and may have evolved to take advantage of high net primary productivity in early spring.
Second, extend the duration of the drain phase in the floodplain. Increasing the duration of the drain phase allows accumulation of total, diatom, and green algal biomass. Accumulation of phytoplankton biomass facilitates efficient bottom-up transfer of energy through the food web by aggregating food resources of optimum size and high carbon content for use by aquatic organisms. Most fish species only remain in the floodplain for a short period and aggregation of food resources may reduce the energy needed for fish to obtain food as they move through the floodplain. Food aggregation may be particularly important late in the season when high respiration reduces net primary productivity.
Third, frequently release small discharges of river water through the floodplain to enhance phytoplankton carbon load to the estuary downstream. The floodplain station had high concentrations of chl a and diatom and green algal biomass, particularly during the drain phase. Regular and small discharge would move this accumulated phytoplankton biomass to the estuary downstream where it can support bottom-up food web production.
Fourth, manage the timing of primary productivity in the floodplain to meet the resource needs of aquatic organisms. The heterotrophy of the flood season at YB called into question the management strategy of enhancing estuarine fishery production in SFE by increasing bottom-up food web production in shallow wetland or floodplain habitats along rivers (CALFED 2000). However, as long as the carbon produced in the floodplain matches the energy needs of food web organisms at high frequency spatial and temporal scales, fishery production should be enhanced regardless of whether the floodplain is autotrophic or heterotrophic over the flood season. Seasonal sums mean little to fish that do not stay in the floodplain throughout the flood season (Sommer et al. 2004a). The successful use of floodplains as a management tool to enhance fishery production will depend on our ability to provide the quantity and quality of food needed by aquatic organisms at different life stages and requires a thorough understanding of the high frequency spatial and temporal variability of food web dynamics in floodplains.
Conclusions
Passing Sacramento River water over the Yolo Bypass floodplain increased net primary productivity and the production of total, diatom, and green algal biomass and phytoplankton cells with wide spherical diameter and high cellular carbon content. The high phytoplankton biomass produced in the floodplain contributed between 14% and 37% of the combined floodplain and riverine load of chl a, diatom, and green algal biomass and phytoplankton cells with wide spherical diameter to the estuary downstream. The greater net primary productivity and phytoplankton biomass in the floodplain than the river over the flood season was associated with high net primary productivity and low respiration in early spring, accumulation of total, diatom, and green algal biomass during the drain phase of the flood pulse cycle and high growth efficiency. This research suggested the quantity and quality of phytoplankton biomass available to the aquatic food web could be enhanced by passing river water through a floodplain during the flood season.
References
Ahearn DS, Viers JH, Mount JF, Dahlgren RA (2006) Priming the productivity pump: flood pulse driven trends in suspended algal biomass distribution across a restored floodplain. Freshwat Biol 51:1417–1433
American Public Health Association, American Water Works Association and Water Environment Association (APHA) (1998) Standard methods for the examination of water and wastewater, 20th edn. American Public Health Association, Washington, DC, USA
Banaszak AT, Neale PJ (2001) Ultraviolet radiation sensitivity of photosynthesis in phytoplankton from an estuarine environment. Limnol Oceanogr 46:592–603
Caffrey JM, Cloern JE, Grenz C (1998) Changes in production and respiration during a spring phytoplankton bloom in San Francisco Bay, California, USA: implications for net ecosystem metabolism. Mar Ecol Prog Ser 172:1–12
California Bay-Delta Authority Programmatic Record of Decision (CALFED) (2000) The CALFED Bay Delta Ecossytem Restoration Program, Sacramento, CA. http://www.calfed.water.ca.gov/Archives/
Cole BE, Cloern JE (1984) Significance of biomass and light availability to phytoplankton productivity in San Francisco Bay. Mar Ecol Prog Ser 17:15–24
Cole JJ, Caraco NF, Peierls BL (1992) Can phytoplankton maintain a positive carbon balance in a turbid, freshwater, tidal estuary? Limnol Oceanogr 37:1608–1617
Forsberg BR, Araujo-Lima CARM, Martinelli LA, Victoria RL, Bonassi JA (1993) Autotrophic carbon sources for fish of the central Amazon. Ecology 74:643–652
Frenette J-J, Demers S, Legendre L, Dodson J (1993) Lack of agreement among models for estimating the photosynthetic parameters. Limnol Oceanogr 38:679–687
Garcia de Emiliani MO (1997) Effects of water level fluctuations on phytoplankton in a river-floodplain lake system (Parana River, Argentina). Hydrobiologia 357:1–15
Hansen BH, Bjornsen PK, Hansen PJ (1994) The size ratio between planktonic predators and their prey. Limnol Oceanogr 39:395–403
Hein T, Heiler G, Pennetzdorfer D, Riedler P, Schagerl M, Schiemer F (1999) The Danube Restoration Project: functional aspects and planktonic productivity in the floodplain system. Regul Rivers Res Manage 15:259–270
Hein T, Baranyi C, Reckendorfer W, Schiemer F (2004) The impact of surface water exchange on the nutrient and particle dynamics in side-arms along the River Danube, Austria. Sci Total Environ 328:207–218
Heip CHR, Goosen NK, Herman PMJ, Kromkamp J, Middelburg JJ, Soetaert K (1995) Production and consumption of biological particles in temperate tidal estuaries. Oceanogr Mar Biol Annu Rev 33:1–149
Jassby AD (2005) Phytoplankton regulation in a eutrophic tidal river (San Joaquin River, California). San Francisco Estuaries Watershed Sci 3:1–2
Jassby AD, Cloern JE (2000) Organic matter sources and rehabilitation of the Sacramento-San Joaquin Delta (California, USA). Aquat Conserv Mar Fresh Ecosys 10:323–352
Jassby AD, Cloern JE, Cole BE (2002) Annual primary production: patterns and mechanisms of change in a nutrient-rich tidal ecosystem. Limnol Oceanogr 47:698–712
Jassby AD, Kimmerer WJ, Monismith SG, Armor C, Cloern JE, Powell TM, Schubel JR, Vendlinski TJ (1995) Isohaline position as a habitat indicator for estuarine populations. Ecol Appl 5:272–289
Jassby AD, Platt T (1976) Mathematical formulation of the relationship between photosynthesis and light for phytoplankton. Limnol Oceanogr 21:540–547
Junk WJ, Bayley PB, Sparks RE (1989) The flood-pulse concept in river-floodplain systems. In Dodge DP (ed) Proceedings of the international large river symposium. Can Spec Pub Fish Aquat Sci 106:110–127
Keckeis S, Baranyi C, Hein T, Holarek C, Riedler P, Schiemer F (2003) The significance of zooplankton grazing in a floodplain system of the River Danube. J Plank Res 25:243–253
Kemp WM, Smith EM, Marvin-DiPasquale M, Boynton WR (1997) Organic carbon balance and net ecosystem metabolism in Chesapeake Bay. Mar Ecol Prog Ser 150:229–248
Kiss KT (1987) Phytoplankton studies in the Szigetkoz section of the Danube during 1981–1982. Arch Hydrobiol 78. Algol Stud 47:247–273
Langdon C (1988) On the causes of interspecific differences in the growth-irradiance relationship for phytoplankton. II. A general review. J Plank Res 10:1291–1312
Lehman PW (1992) Environmental factors associated with long-term changes in chlorophyll concentration in the Sacramento-San Joaquin delta and Suisun Bay, California. Estuaries 15:335–348
Lehman PW (1996) Changes in chlorophyll a concentration and phytoplankton community composition with water-year type in the upper San Francisco Bay Estuary. In: Hollibaugh JT (ed) San Francisco Bay: the ecosystem. Pac Div Am Assoc Adv Sci, San Francisco, CA, pp 351–374
Lehman PW (2000) Phytoplankton biomass, cell diameter and species composition in the low salinity zone of northern San Francisco Bay Estuary. Estuaries 23:216–230
Lehman PW (2004) The influence of climate on mechanistic pathways that affect lower food web production in northern San Francisco Bay Estuary. Estuaries 27:312–325
Lewis WM Jr (1988) Primary production in the Orinoco River. Ecology 69:679–692
Lewis WM Jr, Hamilton SK, Rodriguez MA, Saunders JF III, Last MA (2001) Food web analysis of the Orinoco floodplain based on production estimates and stable isotope data. J N Am Benthol Soc 20:241–254
Lucas LV, Cloern JE, Koseff JR, Monismith SG, Thompson J (1998) Does the Sverdrup critical depth model explain bloom dynamics in estuaries? Mar Res 56:375–415
Lucas LV, Cloern JE, Thompson JK, Monsen NE (2002) Functional variability of habitats within the Sacramento-San Joaquin Delta: restoration implications. Ecol Appl 12:1528–1547
Mallin MA, Pearl HW (1992) Effects of variable irradiance on phytoplankton productivity in shallow estuaries. Limnol Oceanogr 37:54–62
Menden-Deuer S, Lessard EJ (2000) Carbon to volume relationships for dinoflagellates, diatoms and other protest plankton. Limnol Oceanogr 45:569–579
Muller-Solger AB, Jassby AD, Muller-Navarra DC (2002) Nutritional quality for zooplankton (Daphnia) in a tidal freshwater system (Sacramento-San Joaquin River Delta, USA). Limnol Oceanogr 47:1468–1476
Neale PJ, Talling JF, Heaney SI, Reynolds CS, Lund JWG (1991) Long time series from the English Lake District: irradiance-dependent phytoplankton dynamics during the spring maximum. Limnol Oceanogr 36:751–760
Orsi JJ, Mecum WL (1996) Food limitation as the probable cause of a long-term decline in the abundance of Neomysis mercedis the opossum shrimp in the Sacramento-San Joaquin Estuary. In: Hollibaugh JT (ed) San Francisco Bay: the ecosystem. Pac Div Am Assoc Adv Sci, San Francisco, CA, pp 375–401
Platt T, Sathyendranath S (1990) Primary production by phytoplankton: analytic solutions for daily rates per unit area of water surface. Proc Royal Soc London B 241:101–111
Reynolds CS (1997) Vegetation processes in the pelagic: a model for ecosystem theory. In: Kinne O (ed) Excellence in ecology. Ecology Institute Nordbünte, Germany
Rudek J, Cloern JE (1996) Planktonic respiration rates in San Francisco Bay. In: Hollibaugh JT (ed) San Francisco Bay: the ecosystem. Pac Div Am Assoc Adv Sci, San Francisco, CA, pp 289–304
SAS Institute, Inc (SAS) (2004) SAS/STAT User’s Guide, Version 8. SAS Institute Inc., SAS Campus Drive, Cary, NC, USA
Schemel LE, Cox MH (2007) Floodwater chemistry in the Yolo Bypass during winter and spring 1998. US Geological Survey Open File Report 2007–1025
Schemel LE, Sommer TR, Muller-Solger AB, Harrell WC (2004) Hydrologic variability, water chemistry, and phytoplankton biomass in a large floodplain of the Sacramento River, CA, USA. Hydrobiologia 513:129–139
Smith EM, Kemp WM (2001) Size structure and the production/respiration balance in a coastal plankton community. Limnol Oceanogr 46:473–485
Sobczak WV, Cloern JE, Jassby AD, Muller-Solger AB (2002) Bioavailability of organic matter in a highly disturbed estuary: the role of detrital and algal resources. Proc Natl Acad Sci USA 99:8101–8105
Sommer T, Harrell B, Nobriga M, Brown R, Moyle P, Kimmerer W, Schemel L (2001a) California’s Yolo Bypass: evidence that flood control can be compatible with fisheries, wetlands, wildlife, and agriculture. Fisheries 26:6–16
Sommer TR, Nobriga ML, Harrell WC, Batham W, Kimmerer WJ (2001b) Floodplain rearing of juvenile Chinook salmon: evidence of enhanced growth and survival. Can J Fish Aquat Sci 58:325–333
Sommer TR, Harrell WC, Kurth R, Feyrer F, Zeug SC, O’Leary G (2004a) Ecological patterns of early life stages of fishes in a large river-floodplain of the San Francisco estuary. Am Fish Soc Symp 39:111–123
Sommer TR, Harrell WC, Mueller Solger AB, Tom B, Kimmerer W (2004b) Effects of flow variation on channel and floodplain biota and habitats of the Sacramento River, California, USA. Aquat Conserv Mar Freshwater Ecol 14:247–261
Tockner K, Pennetzdorfer D, Reiner N, Schiemer F, Ward JV (1999) Hydrological connectivity, and the exchange of organic matter and nutrients in a dynamic river-floodplain system (Danube, Austria). Freshwater Biol 41:521–535
United States Environmental Protection Agency (US EPA) (1983) Methods for Chemical Analysis of Water and Wastes. Technical Report EPA-600/4-79-020. United States Environmental Protection Agency, Washington, DC, USA
United States Geological Survey (USGS) (1985) Methods for determination of inorganic substances in water and fluvial sediments. United States Geological Survey. Open file report 85-495
Unrein F (2002) Changes in phytoplankton community along a transversal section of the Lower Parana floodplain, Argentina. Hydrobiologia 468:123–134
Utermöhl H (1958) Zur Vervollkommung der quantitativen Phytoplankton-methodik. Mitt Int Ver Theor Angew Limnol 9:1–38
Van den Brink FWB, Van Katwijk WM, Van der Velde G (1994) Impact of hydrology on phyto- and zooplankton community composition in floodplain lakes along the Lower Rhine and Meuse. J Plank Res 16:351–373
Van den Brink FWB, De Leeuw JPHM, Van der Velde G, Verheggen GM (1993) Impact of hydrology on the chemistry and phytoplankton development in floodplain lakes along the Lower Rhine and Meuse. Biogeochemistry 19:103–128
Vollenweider RA (1974) A manual on methods for measuring primary production in aquatic environments. International biological program handbook, vol 12. Balckwell Scientific Publications, Oxford
Ward AK, Wetzel RG (1980) Photosynthetic responses of blue-green algal populations to variable light intensities. Archiv Hydrobiol 90:129–138
Acknowledgments
This research was funded by research grants from the Sacramento-San Joaquin River Interagency Ecological Program Special Studies Program and the California Bay-Delta Authority (CALFED). Technical assistance was provided by W. Harrell and his staff.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lehman, P.W., Sommer, T. & Rivard, L. The influence of floodplain habitat on the quantity and quality of riverine phytoplankton carbon produced during the flood season in San Francisco Estuary. Aquat Ecol 42, 363–378 (2008). https://doi.org/10.1007/s10452-007-9102-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10452-007-9102-6