
Abstract
The Western honey bee (Apis mellifera) is responsible for ecosystem services (pollination) worth US$215 billion annually worldwide and the number of managed colonies has increased 45% since 1961. However, in Europe and the U.S., two distinct phenomena; long-term declines in colony numbers and increasing annual colony losses, have led to significant interest in their causes and environmental implications. The most important drivers of a long-term decline in colony numbers appear to be socioeconomic and political pressure on honey production. In contrast, annual colony losses seem to be driven mainly by the spread of introduced pathogens and pests, and management problems due to a long-term intensification of production and the transition from large numbers of small apiaries to fewer, larger operations. We conclude that, while other causal hypotheses have received substantial interest, the role of pests, pathogens, and management issues requires increased attention.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Summary
There has been significant concern over managed honey bee colony declines and losses given their role as crop pollinators and concurrent concern regarding perceived global pollinator declines. Political and socioeconomic factors have contributed to colony declines in Europe and the U.S. over the past half-century. Yet the overall global number of colonies has increased 45% since 1961. Although many environmental and anthropogenic factors remain under investigation for their role in annual honey bee colony losses, the introduction of pests and pathogens, and large-scale shifts in management practices may be significant, under-researched drivers of colony losses in Europe and North America.
Introduction
There has been significant concern over managed honey bee colony declines and losses, coupled with a growing interest in the long-term fate of global pollinator populations. The Western or European honey bee (Apis mellifera) is a semi-free-ranging managed agricultural species (i.e., managed wildlife species) that pollinates US$215 billion worth of crops worldwide (vanEngelsdorp et al. 2008; Gallai et al. 2009). While the number of colonies has increased 45% globally since 1961, the proportion of pollinator-dependent crops has risen by 300% (Aizen and Harder 2009). To perform the ecosystem service of pollinating agricultural crops honey bees are now managed intensively (particularly in the U.S.) as mobile pollinators, with millions moved annually by truck across the country (Sumner and Boriss 2006).
However, in recent decades both long-term (multi-year) declines and annual losses in honey bee colonies have occurred throughout Europe and the U.S. In this paper, we review the literature on patterns of bee declines and losses, their hypothesized causes, and the environmental, socioeconomic, and agricultural changes likely involved.
Long-Term Colony Declines
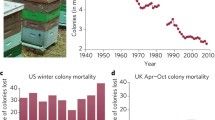
Long-term declines in honey bee colony numbers (i.e., the steady decrease in the overall number of colonies over time) in Europe and the U.S. appear to be associated with political and socioeconomic factors. They have involved more than a 25% decrease of European colonies since the mid-1980s and nearly a 60% decrease in the U.S. (Fig. 1) (Potts et al. 2010a). The collapse of the U.S.S.R. in the 1990s reduced governmental financing of beekeeping in former Soviet countries of eastern Europe, causing a 50% reduction in managed colony numbers there (Aizen and Harder 2009). In western Europe, higher production costs, competition from cheaper imported honey, and increased affordability of sugar-based products have led to a 30% decrease in beekeepers and a 25% decrease in colony numbers since 1985 (Aizen and Harder 2009; Potts et al. 2010a). Honey demand and prices had already fallen in the U.S. at the end of WWII, making beekeeping less profitable. Declines were compounded by rapid expansion of honey importation from China, Argentina, and Vietnam during the 1960s leading to reduced demand for domestic honey (Daberkow et al. 2009; vanEngelsdorp and Meixner 2010). Other factors include suspension of federal support of honey prices in the U.S. in 1996 and the cessation of exports from the U.S. to Canada in 1987 which led to a loss of 250,000 colonies a year in California alone (vanEngelsdorp and Meixner 2010).

Long-term declines in the number of honey-producing honey bee colonies (from 5.9 million in 1947 to 2.3 million in 2008), as reported by the USDA National Agricultural Statistical Service (NASS), and adapted from Potts et al. (2010a). Colony numbers reflect the highest year-to-year increases since 1944 between 2008 and 2010.
Additionally, long-term declines in the U.S. may be artificially heightened by the counting methods used by the USDA National Agricultural Statistics Service (NASS). A widely cited reduction by one million colonies during the 1980s (Fig. 1) coincides with NASS discontinuing the inclusion of colonies from operations with five or fewer hives (vanEngelsdorp and Meixner 2010). Thus, key political and socioeconomic factors including the process of data collection have played a primary role in the steady decline in honey bee colony numbers in Europe and the U.S.
Annual Colony Losses
In contrast to long-term colony declines, annual colony losses are designated as colonies that fail each year and (most frequently reported during the overwintering period) are often replenished by beekeepers “splitting” colonies. As colony losses are only officially counted once per year, they frequently do not include colonies lost during other seasons. This paucity of data on non-overwintering losses leads to a significant bias in the assessment of honey bee health. In the U.S. annual colony losses doubled from the average expected 15 to 30% in 2006–2007 and have remained elevated (vanEngelsdorp et al. 2007). A series of sudden and severe annual losses without evidence of dead bees or hive invaders, but the retention of brood and a queen, was reported with these elevated losses in 2006 and termed “colony collapse disorder” (CCD) (vanEngelsdorp et al. 2009b). The causes and significance of CCD remain unclear (Fig. 2). However, the number of U.S. operations reporting CCD declined in recent annual surveys (vanEngelsdorp et al. 2008, 2011), with no reports in neighboring Canada despite annual losses at a similar scale to the U.S. since 2006 (Currie et al. 2010). CCD was reported by beekeepers as the eighth most important contributor to colony losses in 2009 (vanEngelsdorp et al. 2010; Williams et al. 2010). In 2010 and 2011, only 4 and 7% of beekeepers, respectively, listed CCD as a cause for their losses (vanEngelsdorp et al. 2011, 2012). Regardless of prevalence, some authors have questioned the validity of CCD as a clearly defined syndrome, especially when CCD is defined solely by colony loss with the absence of dead bees (the primary clinical sign investigated in the national beekeeper surveys) rather than the full accepted definition (vanEngelsdorp et al. 2007, 2008, 2010, 2011). However, in many cases adequate disease and health monitoring of an individual colony prior to its decline is absent. Many European countries have also reported elevated annual losses in recent years but similarly the lack of data standardization and the capacity of beekeepers to replenish colonies masks trends (Potts et al. 2010b; van der Zee et al. 2012).

The causes of U.S. honey bee overwintering losses from 2007 to 2011, self-reported by beekeepers through surveys conducted by the USDA, the Apiary Inspectors of America and online surveys through beekeeper associations, summarized by vanEngelsdorp et al. Respondents had the option to report multiple causes for colony loss each year; each cause is reported as percent of total responses given. Double dagger includes colonies reported to have been weak in the fall and usual overwintering losses. Dagger Primarily V. destructor; however, in some years reported as “invertebrate pests” including tracheal mites (Acarapis woodi) and the small hive beetle (Aethina tumida). Asterisks defined only by the loss of a colony with the absence of dead bees—and thus true cases of CCD are likely to be overestimated in all years.
Hypothesized Causes of Annual Colony Losses
Pathogens and Pests
Pathogens and pests are known to cause annual colony losses and have been linked, hypothetically, to long-term declines in bee populations. Prior to the 1970s, diseases of bee larvae (brood diseases) were the most economically important threats, e.g., American Foulbrood (Paenibacillus larvae). These continue to cause significant losses and have global ranges (vanEngelsdorp and Meixner 2010). However in the 1970s and 1980s the parasitic mite Varroa destructor was introduced to honey bees in Europe and in North America respectively (Rosenkranz et al. 2010). This is now a pan-global pest with the exception of Australia, several isolated islands, and possibly some Central African countries (Potts et al. 2010a). This pest is a key factor in recently elevated colony losses in Canada (Currie et al. 2010; Guzman-Novoa et al. 2010) and much of Europe (Aston 2010; Chauzat et al. 2010; Gajger et al. 2010; Le Conte et al. 2010; Mutinelli et al. 2010; Neumann and Carreck 2010; Vejsnaes et al. 2010; Potts et al. 2010b), whereas regions in which V. destructor is not a significant problem have not experienced similar losses (Neumann and Carreck 2010).
The V. destructor mite transmits pathogenic viruses (Gisder et al. 2009) including deformed wing virus (DWV), which results in overt infection significantly correlated with colony loss (Genersch et al. 2010). Three other viruses, acute bee paralysis virus, Israeli acute paralysis virus, and Kashmir bee virus, also increase in virulence when transmitted by V. destructor and all are recognized as causes of colony loss in regions affected by the mite (Ribiere 2008; Highfield et al. 2009; Genersch and Aubert 2010). However, lack of standardized surveillance by beekeepers, inconsistency among studies, and the complexity of transmission dynamics hampers an assessment of their true significance. For example most studies report viral presence or absence, but do not collect sufficient clinical data over time to assess the relative contribution of a virus in a given colony loss event. Interactions between viruses, honey bee immunology, and strain variation may play a role in how an individual or colony responds to viral infection. Recent analysis of the emergence of V. destructor in the Hawaiian archipelago suggests that introduction of the mite leads to increased prevalence and intensity of infection by DWV, and the dominance of a single virulent strain (Martin et al. 2012). The rise of this virulent strain in Hawaii lagged the introduction of the mite and may explain the lag in honey bee losses relative to the introduction of the vector elsewhere. This strongly suggests that the consequences of both Varroa introduction and the viruses it harbors are significant for global honey bee losses.
The microsporidian Nosema apis is the cause of bee dysentery. A close relative, N. ceranae from the Asian honey bee, has recently emerged in A. mellifera and may have replaced N. apis throughout much of its range (Klee et al. 2007, Chen et al. 2008). N. ceranae has been cited as the cause of significant colony losses in Spain (Higes et al. 2007, 2008), though such virulence has not been demonstrated elsewhere (Gisder et al. 2010). Further research is needed to understand the role of N. ceranae in honey bee losses and any possible additive or synergistic effects it may have with other stressors.
The recent, pan-global emergence of honey bee diseases correlates with broad trends in disease emergence (Daszak et al. 2000). These include a growing number of diseases spread by globalized trade and travel—a process termed “Pathogen Pollution” (Cunningham et al. 2003). The anthropogenic introduction of exotic pathogens and pests is the most common cause of disease emergence in crop plants (Anderson et al. 2004), and is responsible for the emergence of a wide range of diseases affecting livestock (e.g., H5N1 avian influenza, Rift Valley fever—Kilpatrick et al. 2006a; Al-Afaleq and Hussein 2011) and humans (e.g., West Nile virus—Kilpatrick et al. 2006a, b; Al-Afaleq and Hussein 2011). Honey bee pathogens have emerged and spread through trade in bees, bee products, and equipment [e.g., the introduction of tracheal mite (Acarapis woodi) and V. destructor to the Americas (Mutinelli 2011), and the spread of V. destructor and N. ceranae from the Asian honey bee (A. ceranae) to A mellifera]. Recently, the World Organisation for Animal Health (or Office Internationale des Epizooties) has made a number of bee pathogens notifiable under regulations approved by country signatories of the World Trade Organization (Mutinelli 2011). This enables restriction of trade in bees and bee products from countries that have identified these pathogens in their bee populations.
Agrochemical Drivers
It has been hypothesized that the application of pesticides, in particular neonicotinoid insecticides, plays a significant role in honey bee colony losses. Neonicotinoids are extensively applied to some crops commercially pollinated by honey bees (Johnson et al. 2010). Neonicotinoids are systemic insecticides that can be applied to seeds and absorbed into the growing plant tissue, making them highly effective, but allowing residues to be present in pollen and nectar (Thompson 2010; Creswell 2011). The application of pesticides directly to seed has environmental advantages over widespread spray treatments such as reduced run-off, less frequent non-target species toxicity (Thompson 2010), and reduction of pesticide use per hectare. Acute honey bee mortality events caused by the accidental drift of dust or spray from pesticide applications have occurred (Krupke et al. 2012). While drift reducing technologies have helped reduce the risk (Thompson 2010), a recent study revealed colony losses may still occur due to pesticide drift (Tapparo et al. 2012).
Exposure to pesticides may also result in sublethal effects on the health of individual honey bees that could lead to reduced survival. Recent studies suggest that low-level pesticide exposure can impair immune system functioning, learning ability, memory, foraging behavior, and odor discrimination in honey bees (Desneux et al. 2007; Yang et al. 2008; Alaux et al. 2010; Williamson and Wright 2013). Further, chronic exposure to pesticides that would otherwise have no harmful effects for healthy colonies may interact with pathogens to produce harmful consequences for colonies already vulnerable to disease (Thompson 2003; Vidau et al. 2011; Aufauvre et al. 2012). One study suggested that individual honey bee vulnerability to Nosema infection is enhanced by the presence of the neonicotinoid imidacloprid, but the parent colonies failed to show increased Nosema levels due to exposure in this study (Pettis et al. 2012).
Simulated exposure events on free-ranging foragers showed that exposure of honey bees to the neonicotinoid thiamethoxam at low levels caused mortality, perhaps due to homing failure, and that this may contribute to colony loss (Henry et al. 2012). However, the bioavailability of the active ingredient in the treated bees was not measured in this study, and is a critical issue to investigate in general. Studies have detected low levels of neonicotinoids (1–10 μg kg−1) in the pollen and nectar of treated plants throughout the blooming period (Creswell 2011) and some scientists argue that bees likely ingest nectar repeatedly through several flower visits, thus increasing their exposure (Yang et al. 2008; Creswell 2011). Other research has shown that chronic exposure to the neonicotinoid imidacloprid at similar sublethal doses has negligible effects at the colony level (Nguyen et al. 2009). Field exposure of bees to flowering canola (Brassica napus) grown from seed treated with the neonicotinoid clothianidin had negligible impact on acute mortality, longevity, brood development, and over-winter survival (Cutler and Scott-Dupree 2007). However, control and treatment fields in that study were in close proximity (295 m), with a small radius (1 km) surrounding the test areas for which alternative foraging options were controlled. Elevated levels of the crop fungicide chlorothalonil have been reported in entombed pollen (pollen presumably sealed off within the hive as a protective measure) (vanEngelsdorp et al. 2009a), and the presence of entombed pollen has been associated with increased colony mortality. However, larvae and adult bees that were fed diets supplemented with entombed pollen did not experience increased mortality (vanEngelsdorp et al. 2009a). Similarly, a laboratory study (Girolami et al. 2009) found extremely high concentrations of neonicotinoids in the guttation drops of corn plants and demonstrated that when bees consumed this fluid, death occurred within a few minutes. The guttation drops were supplemented with a sugar solution to stimulate consumption and there is continued debate as to whether honeybees use guttation fluid as a source of water in relevant crop species. Other studies did not find detrimental effects when conducting similar research with sugar beets, and when alternative drinking sources were available as might occur in nature (Keppler et al. 2010; OPERA 2011). While several recent studies have used models to suggest colony level effects (Henry et al. 2012; Whitehorn et al. 2012), a confirmed causal relationship between environmental pesticide residues and CCD or significant colony loss has yet to be demonstrated (Johnson et al. 2010). The most difficult challenge facing experimental researchers lies in deducing whether sublethal effects on individuals play a role in colony loss under real field conditions.
Beekeepers commonly use miticides to reduce the impact of varroasis. Miticides are placed directly in the hive, and are lipophilic and accumulate in the wax, particularly in the comb where the brood develops (Johnson 2009). Exposure may be exacerbated by synergism, which occurs between low doses of the miticides coumaphos and tau-fluvalinate for example due to binding competition for P450, a class of enzymes important for detoxification (Johnson 2009). Some fungicides found in hives inhibit ergosterol biosynthesis and thus synergize the toxicity of pyrethroid insecticides to honey bees (Johnson 2009). Finally, bees exposed to the antibiotic oxytetracycline which is applied within the hive, were significantly more sensitive to coumaphos and tau-fluvalinate (Hawthorne and Dively 2011). Effects of additive exposure to cholinergic pesticides such as imidacloprid and coumaphos have also been found (Palmer et al. 2013). Further, recent studies examining in-hive chemicals have detected a high diversity of miticides and agricultural pesticides in bee colonies (vanEngelsdorp et al. 2009b; Mullin et al. 2010). Although high levels of intoxicants were detected, no direct correlation between pesticides found in hives and colony loss was found (aside from higher levels of coumaphos in healthy colonies) (vanEngelsdorp et al. 2009b). Although recent studies are beginning to address how bees process these toxins by determining what factors affect the ability of honey bees to upregulate P450 detoxification genes in response to in-hive chemicals (Johnson et al. 2012), additional research into the potential additive and synergistic effects is still needed.
Other Causal Hypotheses
Some authors have proposed that multiple drivers act together, or secondarily to broader changes that may affect the ability of honey bees to resist declines, or that they directly cause immunosuppression. These broader changes include poor genetic diversity, weather events, stress, or inadequate nutrition (Potts et al. 2010a). There has been little research on whether long distance transportation of colonies causes stress, but it has not been linked to losses (vanEngelsdorp et al. 2007, 2008, 2010, 2011, 2012) Agricultural intensification, the increase of monoculture acreage, and loss of natural habitat may affect honey bee nutrition, and theoretically colony health; however, this has not been fully investigated (Brodschneider and Crailsheim 2010).
What is the Most Likely Causal Candidate for Annual Colony Losses?
As with any large-scale biological phenomenon, competing and often conflicting hypotheses are difficult to resolve. However, the evidence from the current state of the field suggests that some individual drivers appear to be more important in honey bee annual losses than others. For example, surveys of beekeepers and published reports suggest elevated colony losses in much of Europe and Canada are attributed to V. destructor and its associated pathogens, as well as known environmental and managerial challenges such as weather events and starvation (Aston 2010; Chauzat et al. 2010; Gajger et al. 2010; Le Conte et al. 2010; Mutinelli et al. 2010; Neumann and Carreck 2010; Vejsnaes et al. 2010; Potts et al. 2010b). Similarly, the majority of losses in the U.S. have been attributed to one or more known causes such as weather events, starvation, queen failure, and mites by beekeepers in recent surveys (vanEngelsdorp et al. 2010, 2011, 2012). However, these reports do not take into consideration the presence or absence of rigorous health surveillance in these colonies prior to the loss.
The hypothesis that honey bees have become immunosuppressed, making them more susceptible to pathogens or pests, parallels similar arguments used for wildlife population declines that were later attributed to single causal factors, e.g., amphibian declines due to the emerging disease chytridiomycosis (Daszak et al. 2003). Caution needs to be applied to this hypothesis, particularly when so little is known of honey bee innate and social immunity. Honey bees have approximately one-third of the immunity genes of other insects, but as colonial insects they also rely on social immunity where colonial behavior protects the colony (Evans and Spivak 2010). Additionally, balanced nutrition including pollen and propolis has recently been demonstrated to induce detoxifying enzymes in individual honey bees (Mao et al. 2013). Therefore, studies that experimentally show an effect on immunity in individual honey bees cannot be readily extrapolated to an effect on the colony in the field.
Perhaps the most parsimonious hypothesis for the cause of annual losses is that the relatively recent introduction of Varroa mites, and the resultant expansion of the range, prevalence, or intensity of infection of viral strains they harbor, has had a dramatic impact on honey bees. Thus, rather than immunosuppression of honey bee immunity as the underlying condition, it may be the relative naivety of bee colonies to Varroa and viral strains causing their decline and susceptibility to other factors. This hypothesis is supported by recent findings that the increase in prevalence and dominance of virulent viral strains lags behind Varroa introduction and leads to a complex, but probably significant, relationship between pest introduction, pathogen emergence, and honey bee colony losses (Martin et al. 2012). In a similar scenario, arguments that disease-induced amphibian population declines were only possible due to immunosuppression by another driver have now been largely refuted. In this case, the international trade in amphibians has now been widely accepted as the primary driver of the emergence of this disease (Daszak et al. 2003; Fisher et al. 2012). This is consistent with the increasing importance of trade and travel in the emergence and spread of diseases in humans, livestock, and wildlife globally (Smolinski et al. 2003; Karesh et al. 2005; Fevre et al. 2006; Hosseini et al. 2010) and in other types of biological invasions (Levine and D’Antonio 2003).
Critical Research and Policy Gaps
New Approaches to Investigating Causal Factors
Investigating the causes of long-term population declines and die-offs in wildlife or free-ranging domesticated species requires long-term data on all hypothesized causal drivers. In cases where multiple drivers may be involved, a framework to deduce the importance, or strength, of each driver is required. Similar problems abound in disease ecology, where pathogens responsible for population declines are affected by climate, land use change, trade, pollutants, and the interaction of a myriad of other drivers. In these cases, traditional hypothesis testing may be supplemented by modeling, risk analysis and strong inference methods that provide a measure of the relative role of each driver (Plowright et al. 2008). A macroecology approach (Gaston and Blackburn 2000; Blackburn and Gaston 2004) involving a broad view of ecological systems based on model selection could complement experimental studies on honey bee colony losses. For example, it would be possible to make statistical comparisons among different potential hypotheses at a broader scale (Fig. 3). Each hypothesis could be ranked and weighted giving a quantifiable measure of support. When similar levels of support for each hypothesis is found, model averaging can be used to make robust estimations and predictions (Burnham and Anderson 2002; Johnson and Omland 2004).

A complementary analysis framework to study honey bee losses. This macroecological approach uses a top-down perspective and statistical methodologies (e.g., model selection) to contrast and to explain observed patterns at different temporal and spatial scales.
For honey bee colony losses, long-term datasets on bee colony dynamics, land use change, Varroa destructor prevalence, bee pathogen distribution and prevalence, pesticide exposure, and a range of other hypothesized causes are not uniformly available across the geographic range of reported losses. Furthermore, there are differences among regions in the manner data have been collected, and honey bee losses defined and reported. While specific tests of some hypotheses have been published, these are relatively few and are still insufficient to deduce cause. More importantly, there are a large number of hypothesized drivers that have not yet been studied at a scale relevant to understanding the ecology of bee losses.
Varroa destructor Research
Studies to deduce the role of V. destructor in colony losses would require information on the introduction of Varroa into new regions. At the individual apiary level, there are rarely adequate monitoring history and records available of Varroa levels in each colony over time prior to a loss event. Rather, investigation into the role of Varroa in losses is usually attempted after the fact. Until more rigorous health monitoring techniques are implemented, mathematical modeling of the spread of Varroa at a regional level might provide insight, and models could be back-tested by using data on first reports of the pest from states, provinces, and counties following introduction. Metapopulation models which explicitly estimate the connectivity among colonies may provide a way to estimate the seasonal spread of pathogens, their ability to cause the extensive colony loss in some regions, and their likely patterns of persistence. Testing this hypothesis might be aided by the presence of regions where Varroa is absent (e.g., Victoria, British Columbia) and by studies that monitor the impact of the mite over a number of years on longitudinal experimental colonies.
Intensification of Honey Bee Production
The ratio of commercial beekeeper to (small-scale) hobbyist operations has increased in many countries due to the increasing demand for pollination services to commercial crops (Daberkow et al. 2009; Potts et al. 2010b) and rising competition from international honey markets (Le Conte and Navajas 2008). Given the current condition of the U.S. honey market, American beekeepers are less economically rewarded to produce honey and provide their bees with a varied diet compared to providing pollination services. Bees devoted to pollination services are primarily exposed to monoculture nutritional sources (Le Conte and Navajas 2008; Champetier et al. 2010). Between 1987 and 2002 in the U.S., there was a 40% decline in beekeepers owning fewer than 25 colonies, and a 66% increase in colonies managed by commercial beekeepers (Daberkow et al. 2009). Thus, the intensification of bee production in the U.S. is similar to much earlier intensification of livestock production, e.g., the movement from backyard poultry production to commercial rearing in the early twentieth century. This process led to the emergence of agricultural pathogens historically (e.g., Salmonella enteritidis) and currently (e.g., influenza A/H5N1). However, there are important differences between honey bee management and intensively managed livestock. These include a paucity of regulations on veterinary care, disease testing and quarantine of traded animals, use of treatments, nutrition studies to develop dietary supplements, and health and welfare inspection. The manner in which health care, management plans, nutrition modification, and record-keeping are applied in beekeeping operations may need to adapt to the changing levels of intensification to avoid further declines in bee health and increased annual losses. Trained bee health consultants (e.g., extension workers, state apiculturists, veterinarians) may help in this transition. These workers could assist beekeepers in adapting management schemes based on local environmental and disease threats, size of operation, and diagnostic and treatment needs.
Pesticides
There has been a great deal of work on the role of pesticides in bee colony loss leading to significant concern over their effects. Despite this concern and a recent European policy changes on the use of neonicotinoid pesticides, scientific studies examining the overall impact of neonicotinoid pesticides on managed honey bee colony loss are yet to demonstrate colony level losses in field settings and at field doses. Critical issues to be addressed are the lack of comparable losses in some regions where pesticides are used, uncertainty about appropriate field doses, and the methods used in experimental studies. Further research on sublethal and additive or synergistic effects of agricultural chemicals and their residues, using field-based colony level data, and application of spatial analysis methodologies to past and ongoing losses may help elucidate the role of pesticides in losses.
Longitudinal Epidemiological Monitoring and Disease Investigations
The lack of standardized reporting within and among countries makes it difficult to track patterns of colony loss on local or regional scales. Longitudinal epidemiological monitoring that includes long-term studies of patterns of colony loss, disease incidence and pathogen prevalence in single populations, with standardized sampling and data collection protocols would allow effective comparisons among different geographic areas experiencing losses. The COLOSS network (Prevention of Honeybee Colony Losses—http://coloss.org/), the German Honey Bee Monitoring Program (Genersch et al. 2010), the Bee Informed Project (www.beeinformed.org), and USDA’s Managed Pollinator Coordinated Agricultural Project (CAP—www.beeccdcap.uga.edu), have begun efforts along these lines.
Environmental and Ecological Implications
The full range of environmental implications of honey bee colony declines and losses is unclear and some suggest that the role of the managed honey bee in pollinating native and even some crop plants is overestimated (Winfree et al. 2007; Ollerton et al. 2012; Garibaldi and Winfree 2013). Thus, it is difficult to assess how honey bee colony losses relate to global native pollinator declines. Honey bees are an introduced species in many parts of their range (e.g., North America), and they are extensively managed throughout. They do, however, have free contact with wild native pollinators, allowing pathogen exchange, resource competition, and exposure to similar environmental hazards. Yet stressors may or may not affect managed honey bees differently than native or non-managed species. Despite evidence of transmission, the impact of Varroa mites and viruses on native bees at the population level remains unclear. Similarly, pesticides have the potential to affect native pollinators (Potts et al. 2010a), particularly bumble bee (Bombus terrestris) foraging behavior and reproductive success (Gill et al. 2012; Laycock et al. 2012; Whitehorn et al. 2012) although bumble bee life cycle characteristics differ from that of the honey bee. Regardless of the true extent of native pollinator declines (Levine and D’Antonio 2003), it is uncertain if managed honey bee declines and losses are related to native or free-ranging pollinator declines. Regardless, past lessons learned from impacts of agricultural intensification on wildlife should be applied when investigating the potential health links between honey bees and native pollinators, while accounting for the unique nature of managing a semi-free-ranging species.
Conclusion
Causes of long-term declines of honey bees in the U.S. and Europe are most likely due to the loss of beekeepers to the industry and intense competition for bee products. More research is needed on honey bee annual losses, but the global spread of pathogens and pests through the trade in bees and bee products likely plays a key role. These losses compound two critical environmental problems: the role of anthropogenic movement and manipulation of wildlife and their habitat and the role of agricultural intensification on managed species. In many ways, the state of research on honey bee losses mirrors the situation of livestock production in the early twentieth century, with poor coordination of best management practices (including lack of adequate knowledge of health threats and treatments) and a low appreciation of the impact of rapid demographic change on a managed population.
For this issue, some lessons can be learned from amphibian population declines, which were first brought to global attention in the late 1980s and considered enigmatic by their occurrence in presumed pristine locations (Blaustein and Wake 1990). Despite several initial hypotheses of driver interactions (Kiesecker and Blaustein 1995; Davidson et al. 2001; Pounds 2001), the identification of an emerging disease, chytridiomycosis (Berger et al. 1998) led to a marked shift in emphasis. The emergence of this disease, driven by the global trade in amphibians is now widely accepted as the major driver of enigmatic global amphibian population declines (Stuart et al. 2004; Mendelson et al. 2006; Fisher et al. 2012) and has led to the adoption of global policies to reduce its spread (Mendelson et al. 2006; Schloegel et al. 2010). Applying these lessons to the current state of honey bee health suggests that filling research gaps, particularly in management impacts and techniques, will complement additional studies on environmental factors such as nutrition, pesticides, pests, and pathogens to formulate comprehensive solutions to what may become an economically and environmentally costly phenomenon.
References
Aizen, M. A. and L. D. Harder (2009). “The global stock of domesticated honey bees is growing slower than agricultural demand for pollination.” Current Biology 19: 915-918.
Al-Afaleq, A. I. and M. F. Hussein (2011). “The status of Rift Valley fever in animals in Saudi Arabia: a mini review.” Vector-Borne and Zoonotic Diseases 11(12): 1513-1520.
Alaux, C., F. Ducloz, D. Crauser and Y. Le Conte (2010). “Diet effects on honeybee immunocompetence.” Biology Letters 6(4): 562-565.
Anderson, P. K., A. A. Cunningham, N. G. Patel, F. J. Morales, P. R. Epstein and P. Daszak (2004). “Emerging infectious diseases of plants: pathogen pollution, climate change and agrotechnology drivers.” Trends in Ecology & Evolution 19(10): 535-544.
Aston, D. (2010). “Honey bee winter loss survey for England, 2007-8.” Journal of Apicultural Research 49: 111–112.
Aufauvre, J., D. G. Biron, C. Vidau, R. Fontbonne, M. Roudel, M. Diogon, B. Vigues, L. P. Belzunces, F. Delbac and N. Blot (2012). “Parasite-insecticide interactions: a case study of Nosema ceranae and fipronil synergy on honeybee.” Scientific Reports 2: 326.
Berger, L., R. Speare, P. Daszak, D. E. Green, A. A. Cunningham, C. L. Goggin, R. Slocombe, M. A. Ragan, A. D. Hyatt, K. R. McDonald, H. B. Hines, K. R. Lips, G. Marantelli and H. Parkes (1998). “Chytridiomycosis causes amphibian mortality associated with population declines in the rain forests of Australia and Central America.” Proceedings of the National Academy of Sciences of the United States of America 95(15): 9031-9036.
Blackburn, T. and K. Gaston (2004). “Special Feature: Macroecology.” Basic and Applied Ecology 385-387.
Blaustein, A. R. and D. B. Wake (1990). “Declining amphibian populations: a global phenomenon.” Trends in Ecology & Evolution 5: 203–204.
Brodschneider, R. and K. Crailsheim (2010). “Nutrition and health in honey bees.” Apidologie 41: 278–294.
Burnham, K. P. and D. R. Anderson (2002). “Model selection and multimodel inference: a practical information-theoretic approach.” Springer, New York.
Champetier A, Sumner D, Wilen JE (2010) The Bioeconomics of Honeybees and Pollination.
Chauzat, M. P., A. C. Martel, S. Zeggane, P. Drajnudel, F. Schurr, M. C. Clement, M. Ribiere-Chabert, M. Aubert and J. P. Faucon (2010). “A case control study and a survey on mortalities of honey bee colonies (Apis mellifera) in France during the winter of 2005-6.” Journal of Apicultural Research 49(1): 40-51.
Chen, Y., J. D. Evans, B. I. Smith and J. S. Pettis (2008). “Nosema ceranae is a long-present and wide-spread microsporidian infection of the European honey bee (Apis mellifera) in the United States.” Journal of Invertebrate Pathology 97: 186-188.
Creswell, J. E. (2011). “A meta-analysis of experiments testing the effects of a neonicotinoid insecticide (imidacloprid) on honey bees.” Ecotoxicology 20: 149–157.
Cunningham, A. A., P. Daszak and J. P. Rodríguez (2003). “Pathogen pollution: defining a parasitological threat to biodiversity conservation.” Journal of Parasitology 89: S78-S83.
Currie, R. W., S. F. Pernal and E. Guzmán-Novoa (2010). “Honey bee colony losses in Canada.” Journal of Apicultural Research 49: 104–106.
Cutler, G. C. and C. D. Scott-Dupree (2007). “Exposure to clothianidin seed-treated canola has no long-term impact on honey bees.” Journal of Economic Entomology 100: 765–772.
Daberkow, S., P. Korb and F. Hoff (2009). “Structure of the US beekeeping industry: 1982-2002.” Journal of Economic Entomology 102(3): 868-886.
Daszak, P., A. A. Cunningham and A. D. Hyatt (2000). “Emerging infectious diseases of wildlife - threats to biodiversity and human health.” Science 287: 443-449.
Daszak, P., A. A. Cunningham and A. D. Hyatt (2003). “Infectious disease and amphibian population declines.” Diversity and Distributions 9: 141–150.
Davidson, C., H. B. Shaffer and M. R. Jennings (2001). “Declines of the California red-legged frog: Climate, UV-B, habitat, and pesticides hypotheses.” Ecological Applications 11(2): 464-479.
Desneux, N., A. Decourtye and J. M. Delpuech (2007). “The sublethal effects of pesticides on beneficial arthropods.” Annual Review of Entomology 52: 81–106.
Evans, J. D. and M. Spivak (2010). “Socialized medicine: Individual and communal disease barriers in honey bees.” Journal of Invertebrate Pathology 103: S62-S72.
Fevre, E. M., B. M. D. C. Bronsvoort, K. A. Hamilton and S. Cleaveland (2006). “Animal movements and the spread of infectious diseases.” Trends in Microbiology 14(3): 125-131.
Fisher, M. C., D. A. Henk, C. J. Briggs, J. S. Brownstein, L. C. Madoff, S. L. McCraw and S. J. Gurr (2012). “Emerging fungal threats to animal, plant and ecosystem health.” Nature 484(7393): 186-194.
Gajger, I. T., Z. Tomljanovic and Z. Petrinec (2010). “Monitoring health status of Croatian honey bee colonies and possible reasons for winter losses.” Journal of Apicultural Research 49: 107–108.
Gallai, N., J. M. Salles, J. Settele and B. E. Vaissiere (2009). “Economic valuation of the vulnerability of world agriculture confronted with pollinator decline.” Ecological Economics 68(3): 810-821.
Garibaldi, L. A., I. Steffan-Dewenter, R. Winfree, M. A., Aizen, R. Bommarco, S. A. Cunningham, C. KKremen, L. G. Carvalheiro, L.D. Harder, O. Afik, I. Bartomeus, F. Benjamin, V. Boreux, D. Cariveau, N. P. Chacoff, J. H. Dudenhöffer, B. M. Freitas, J. Ghazoul, S. Greenleaf, J. Hipólito, A. Holzschuh, B. Howlett, R. Isaacs, S. K. Javorek, C. M. Kennedy, K. M. Krewenka, S. Srishnan, Y. Mandelik, M. M. Mayfield, I. Motzke, T. Munyuli, B. A. Nault, M. Otieno, J. Petersen, G. Pisanty, S. G. Potts, R. Rader, T. H. Ricketts, M. Rundlöf, C. L. Seymour, C. Schüepp, H. Szentgyörgyi, H. Taki, T. Tscharntke, C. H. Vergara, B. F. Viana, T. C. Wanger, C. Westphal, N. Williams, and A. M. Klein (2013). “Wild Pollinators Enhance Fruit Set of Crops Regardless of Honey Bee Abundance”. Science 339(6127): 1608-1611.
Gaston, K. J. and T. M. Blackburn (2000). Pattern and process in macroecology, Wiley- Blackwell.
Genersch, E. and M. Aubert (2010). “Emerging and re-emerging viruses of the honey bee (Apis mellifera L.).” Veterinary Research 41(6): 54
Genersch, E., W. von der Ohe, H. Kaatz, A. Schroeder, C. Otten, R. Buchler, S. Berg, W. Ritter, W. Muhlen, S. Gisder, M. Meixner, G. Liebig and P. Rosenkranz (2010b). “The German bee monitoring project: a long term study to understand periodically high winter losses of honey bee colonies.” Apidologie 41(3): 332-352.
Gill, R. J., O. Ramos-Rodriguez and N. E. Raine (2012). “Combined pesticide exposure severely impacts individual- and colony-level traits in bees.” Nature 491: 105-109.
Girolami, V., L. Mazzon, A. Squartini, N. Mori, M. Marzaro, A. Di Bernardo, M. Greatti, C. Giorio and A. Tapparo (2009). “Translocation of Neonicotinoid Insecticides From Coated Seeds to Seedling Guttation Drops: A Novel Way of Intoxication for Bees.” Journal of Economic Entomology 102(5): 1808-1815.
Gisder, S., P. Aumeier and E. Genersch (2009). “Deformed wing virus: replication and viral load in mites (Varroa destructor).” Journal of General Virology 90(2): 463-467.
Gisder, S., K. Hedtke, N. Möckel, M.-C. Frielitz, A. Linde and E. Genersch (2010). “Five-year cohort study of Nosema spp. in Germany: Does climate shape virulence and assertiveness of Nosema ceranae?” Applied and Environmental Microbiology 76(9): 3032-3038.
Guzman-Novoa, E., L. Eccles, Y. Calvete, J. McGowan, P. G. Kelly and A. Correa-Benitez (2010). “Varroa destructor is the main culprit for the death and reduced populations of overwintered honey bee (Apis mellifera) colonies in Ontario, Canada.” Apidologie 41(4): 443-450.
Hawthorne, D. J. and G. P. Dively (2011). “Killing them with kindness? In-hive medications may inhibit xenobiotic efflux transporters and endanger honey bees.” PLoS ONE 6(11): e26796.
Henry, M., M. Beguin, F. Requier, O. Rollin, J. F. Odoux, P. Aupinel, J. Aptel, S. Tchamitchian and A. Decourtye (2012). “A Common Pesticide Decreases Foraging Success and Survival in Honey Bees.” Science 336(6079): 348-350.
Higes, M., P. García-Palencia, R. Martín-Hernández and A. Meana (2007). “Experimental infection of Apis mellifera honeybees with Nosema ceranae (Microsporidia).” Journal of Invertebrate Pathology 94: 211-217.
Higes, M., R. Martin-Hernandez, C. Botias, E. Garrido Bailon, A. V. Gonzalez-Porto, L. Barrios, M. Jesus del Nozal, J. L. Bernal, J. J. Jimenez, P. Garcia Palencia and A. Meana (2008). “How natural infection by Nosema ceranae causes honeybee colony collapse.” Environmental Microbiology 10(10): 2659-2669.
Highfield, A. C., A. El Nagar, L. C. M. Mackinder, L. M. L. J. Noel, S. J. Martin and D. C. Schroeder (2009). “Deformed Wing Virus Implicated in Overwintering Honeybee Colony Losses.” Applied and Environmental Microbiology 75(22): 7212-7220.
Hosseini, P., S. H. Sokolow, K. J. Vandegrift, A. M. Kilpatrick and P. Daszak (2010). “Predictive power of air travel and socio-economic data for early pandemic spread.” PLoS ONE 5(9): e12763.
Johnson, J. B. and K. S. Omland (2004). “Model selection in ecology and evolution.” Trends in Ecology & Evolution 19(2): 101-108.
Johnson, R. M. (2009). “Managed pollinator CAP Coordinated Agricultural Project: A national research and extension initiative to reverse pollinator decline when varroacides interact.” American Bee Journal 149: 1157–1159.
Johnson, R. M., M. D. Ellis, C. A. Mullin and M. Frazier (2010). “Pesticides and honey bee toxicity - USA.” Apidologie 41(3): 312-331.
Johnson, R. M., L. Dahlgren, B. D. Siegfried, and M. D. Ellis (2012). “Acaricide, fungicide and drug interactions in honey bees (Apis mellifera). PLoS ONE 8(1): e54092.
Karesh, W. B., R. A. Cook, E. L. Bennett and J. Newcomb (2005). “Wildlife trade and global disease emergence.” Emerging Infectious Diseases 11(7): 1000-1002.
Keppler, J., R. Becker, R. Spatz and F. Dechet (2010). Systemische insektizide Beizmittel – Auftreten und Relevanz von Guttation für die Entwicklung von Honigbienenkolonien. Deutsche Pflanzenschutztagung. J. K.-I. BfKeVG. Quedlinburg, Germany.
Kiesecker, J. M. and A. R. Blaustein (1995). “Synergism between UV-B radiation and a pathogen magnifies amphibian embryo mortality in nature.” Proceedings of the National Academy of Sciences of the United States of America 92: 11049–11052.
Kilpatrick, A. M., A. A. Chmura, D. W. Gibbons, R. C. Fleischer, P. P. Marra and P. Daszak (2006a). “Predicting the global spread of H5N1 avian influenza.” Proceedings of the National Academy of Sciences of the United States of America 103: 19368-19373.
Kilpatrick, A. M., L. D. Kramer, M. J. Jones, P. P. Marra and P. Daszak (2006b). “West Nile virus epidemics in North America are driven by shifts in mosquito feeding behavior.” PloS Biology 4(4): e82.
Klee, J., A. M. Besana, E. Genersch, S. Gisder, A. Nanetti, D. Q. Tam, T. X. Chinh, F. Puerta, J. M. Ruz, P. Kryger, D. Message, F. Hatjina, S. Korpela, I. Fries and R. J. Paxton (2007). “Widespread dispersal of the microsporidian Nosema ceranae, an emergent pathogen of the western honey bee, Apis mellifera.” Journal of Invertebrate Pathology 96(1): 1-10.
Krupke, C. H., G. J. Hunt, B. D. Eitzer, G. Andino and K. Given (2012). “Multiple routes of pesticide exposure for honey bees living near agricultural fields.” PLoS ONE 7(1): e29268.
Laycock, I., K. M. Lenthall, A. T. Barratt and J. E. Creswell (2012). “Effects of imidacloprid, a neonicotinoid pesticide, on reproduction in worker bumble bees (Bombus terrestris).” Ecotoxicology 21(7): 1937-1945.
Le Conte, Y., M. Ellis and W. Ritter (2010). “Varroa mites and honey bee health: can Varroa explain part of the colony losses?” Apidologie 41(3): 353-363.
Le Conte, Y. and M. Navajas (2008). “Climate change: impact on honey bee populations and diseases.” Revue scientifique et technique (International Office of Epizootics) 27: 485-497, 499-510.
Levine, J. M. and C. M. D’Antonio (2003). “Forecasting biological invasions with increasing international trade.” Conservation Biology 17(1): 322-326.
Mao, W., M. A. Shuler and M. R. Berenbaum (2013). “Honey constituents up-regulate detoxification and immunity genes in the western honey bee Apis mellifera.” Proceedings of the National Academy of Sciences of the United States of America 110(22): 8763-8764.
Martin, S. J., A. C. Highfield, L. Brettell, E. M. Villalobos, G. E. Budge, M. Powell, S. Nikaido and D. C. Schroeder (2012). “Global honey bee viral landscape altered by a parasitic mite.” Science 336(6086): 1304-1306.
Mendelson, J. R., K. R. Lips, R. W. Gagliardo, G. B. Rabb, J. P. Collins, J. E. Diffendorfer, P. Daszak, R. Ibanez, K. C. Zippel, D. P. Lawson, K. M. Wright, S. N. Stuart, C. Gascon, H. R. da Silva, P. A. Burrowes, R. L. Joglar, E. La Marca, S. Lotters, L. H. du Preez, C. Weldon, A. Hyatt, J. V. Rodriguez-Mahecha, S. Hunt, H. Robertson, B. Lock, C. J. Raxworthy, D. R. Frost, R. C. Lacy, R. A. Alford, J. A. Campbell, G. Parra-Olea, F. Bolanos, J. J. C. Domingo, T. Halliday, J. B. Murphy, M. H. Wake, L. A. Coloma, S. L. Kuzmin, M. S. Price, K. M. Howell, M. Lau, R. Pethiyagoda, M. Boone, M. J. Lannoo, A. R. Blaustein, A. Dobson, R. A. Griffiths, M. L. Crump, D. B. Wake and E. D. Brodie (2006). “Biodiversity - Confronting amphibian declines and extinctions.” Science 313(5783): 48.
Mullin C. A., M. Frazier, J. L. Frazier, S. Ashcraft, R. Simonds, D. vanEngelsdorp, J. S. Pettis. (2010). “High levels of miticides and agrochemicals in North American apiaries: Implications for honey bee health. PLoS ONE 5(3): e9754.
Mutinelli, F. (2011). “The spread of pathogens through trade in honey bees and their products (including queen bees and semen): overview and recent developments.” Revue Scientifique et Technique - Office International des Epizooties 30: 257–271.
Mutinelli, F., C. Costa, M. Lodesani, A. Baggio, P. Medrzycki, G. Formato and C. Parrini (2010). “Honey bee colony losses in Italy.” Journal of Apicultural Research 49: 119–120.
Neumann, P. and N. L. Carreck (2010). “Honey bee colony losses.” Journal of Apicultural Research 49: 1–6.
Nguyen, B. K., C. Saegerman, C. Pirard, J. Mignon, J. Widart, B. Tuirionet, F. J. Verheggen, D. Berkvens, E. De Pauw and E. Haubruge (2009). “Does imidacloprid seed-treated maize have an impact on honey bee mortality?” Journal of Economic Entomology 102(2): 616-623.
Ollerton, J., V. Price, W. S. Armbruster, J. Memmott, S. Watts, N. M. Waser, O. Totland, D. Goulson, R. Alarcon, J. C. Stout and S. Tarrant (2012). “Overplaying the role of honey bees as pollinators: a comment on Aebi and Neumann (2011).” Trends in Ecology & Evolution 27(3): 141-142.
OPERA (2011). Bee health in Europe - Facts & figures. U. C. d. S. Cuore. Italy.
Palmer MJ, Moffat C, Saranzewa N, Harvey J, Wright GA, Connolly CN (2013) Cholinergic pesticides cause mushroom body neuronal inactivation in honeybees. Nature Communications 4(1634). doi:10.1038/ncomms2648
Pettis, J. S., D. vanEngelsdorp, J. Johnson and G. Dively (2012). “Pesticide exposure in honey bees results in increased levels of the gut pathogen Nosema.” Naturwissenschaften 99(2): 153-158.
Plowright, R. K., S. H. Sokolow, M. E. Gorman, P. Daszak and J. E. Foley (2008). “Causal inference in disease ecology: investigating ecological drivers of disease emergence.” Frontiers in Ecology and the Environment 6(8): 420-429.
Potts, S. G., J. C. Biesmeijer, C. Kremen, P. Neumann, O. Schweiger and W. E. Kunin (2010a). “Global pollinator declines: trends, impacts and drivers.” Trends in Ecology & Evolution 25(6): 345-353.
Potts, S. G., S. P. M. Roberts and R. Dean (2010b). “Declines of managed honey bees and beekeepers in Europe.” Journal of Apicultural Research 49: 15–22.
Pounds, J. A. (2001). “Climate and amphibian declines.” Nature 410(6829): 639-640.
Ribiere M, Ball B, Aubert M (2008) Natural history and geographic distribution of honey bee viruses. In: Virology and the Honey Bee, Aubert M, Ball B, Fries I et al. (editors), Luxembourg: European Communities, pp 15–84.
Rosenkranz, P., P. Aumeier and B. Ziegelmann (2010). “Biology and control of Varroa destructor.” Journal of Invertebrate Pathology 103: S96-S119.
Schloegel, L. M., P. Daszak, A. A. Cunningham, R. Speare and B. Hill (2010). “Two amphibian diseases, chytridiomycosis and ranaviral disease, are now globally notifiable to the World Organization for Animal Health (OIE): an assessment.” Diseases of Aquatic Organisms 92(2-3): 101-108.
Smolinski, M. S., M. A. Hamburg, J. Lederberg and Committee on Emerging Microbial Threats to Health in the 21st Century (2003). Microbial threats to health: Emergence, detection, and response. Washington D.C., The National Academies Press
Stuart, S. N., J. S. Chanson, N. A. Cox, B. E. Young, A. S. L. Rodrigues, D. L. Fischman and R. W. Waller (2004). “Status and trends of amphibian declines and extinctions worldwide.” Science 306(5702): 1783-1786.
Sumner, D. A. and H. Boriss (2006). Bee-conomics and the leap in pollination fees. Agricultural and Resource Economics Update, University of California Gianni Foundation of Agricultural Economics. 9: 9-11.
Tapparo, A., D. Marton and C. Gioro (2012). “Assessment of the environmental exposure of honeybees to particulate matter containing neonicotinoid insecticides coming from corn coated seeds.” Environmental Science & Technology 46: 2592–2599.
Thompson, H. (2010). “Risk assessment for honey bees and pesticides - recent developments and ‘new issues’.” Pest Management Science 66: 1157–1162.
Thompson, H. M. (2003). “Behavioural effects of pesticides in bees - Their potential for use in risk assessment.” Ecotoxicology 12(1-4): 317-330.
van der Zee, R., L. Pisa, S. Andonov, R. Brodschneider, J. D. Charriere, R. Chlebo, M. F. Coffey, K. Crailsheim, B. Dahle, A. Gajda, A. Gray, M. M. Drazic, M. Higes, L. Kauko, A. Kence, M. Kence, N. Kezic, H. Kiprijanovska, J. Kralj, P. Kristiansen, R. M. Hernandez, F. Mutinelli, B. K. Nguyen, C. Otten, A. Ozkirim, S. F. Pernal, M. Peterson, G. Ramsay, V. Santrac, V. Soroker, G. Topolska, A. Uzunov, F. Vejsnaes, S. Wei and S. Wilkins (2012). “Managed honey bee colony losses in Canada, China, Europe, Israel and Turkey, for the winters of 2008-9 and 2009-10.” Journal of Apicultural Research 51(1): 91-114.
vanEngelsdorp, D., D. Caron, J. Hayes, R. Underwood, M. Henson, K. Rennich, A. Spleen, M. Andree, R. Snyder, K. Lee, K. Roccasecca, M. Wilson, J. Wilkes, E. Lengerich, J. Pettis and Bee Informed Partnership (2012). “A national survey of managed honey bee 2010-11 winter colony losses in the USA: results from the Bee Informed Partnership.” Journal of Apicultural Research 51(1): 115-124
vanEngelsdorp, D., J. D. Evans, L. Donovall, C. Mullin, M. Frazier, J. Frazier, D. R. Tarpy, J. Hayes and J. S. Pettis (2009a). “Entombed pollen”: A new condition in honey bee colonies associated with increased risk of colony mortality.” Journal of Invertebrate Pathology 101(2): 147-149.
vanEngelsdorp, D., J. D. Evans, C. Saegerman, C. Mullin, E. Haubruge, B. K. Nguyen, M. Frazier, J. Frazier, D. Cox-Foster, Y. P. Chen, R. Underwood, D. R. Tarpy and J. S. Pettis (2009b). “Colony Collapse Disorder: A descriptive study.” PLoS ONE 4(8): e6481.
vanEngelsdorp, D., J. Hayes, R. M. Underwood, D. Caron and J. Pettis (2011). “A survey of managed honey bee colony losses in the USA, fall 2009 to winter 2010.” Journal of Apicultural Research 50(1): 1-10.
vanEngelsdorp, D., J. Hayes, R. M. Underwood and J. Pettis (2008). “A survey of honey bee colony losses in the US, fall 2007 to spring 2008.” PLoS ONE 3(12): e4071.
vanEngelsdorp, D., J. J. Hayes, R. Underwood and J. S. Pettis (2010). “A survey of honey bee colony losses in the United States, fall 2008 to spring 2009.” Journal of Apicultural Research 49(1): 7-14.
vanEngelsdorp, D. and M. D. Meixner (2010). “A historical review of managed honey bee populations in Europe and the United States and the factors that may affect them.” Journal of Invertebrate Pathology 103: S80-S95.
vanEngelsdorp, D., R. Underwood, D. Caron and J. Hayes (2007). “An estimate of managed colony losses in the winter of 2006-2007: A report commissioned by the Apiary Inspectors of America.” American Bee Journal 147(7): 599-603.
Vejsnaes, F., S. L. Nielsen and P. Kryger (2010). “Factors involved in the recent increase in colony losses in Denmark.” Journal of Apicultural Research 49: 109–110.
Vidau, C., M. Diogon, J. Aufauvre, R. Fontbonne, B. Vigues, J. L. Brunet, C. Texier, D. G. Biron, N. Blot, H. El Alaoui, L. P. Belzunces and F. Delbac (2011). “Exposure to Sublethal Doses of Fripronil and Thiacloprid Highy lncreases Mortality of Honeybees Previously Infected by Nosema ceranae.” PLoS ONE 6(6).
Whitehorn, P. R., S. O’Connor, F. L. Wackers and D. Goulson (2012). “Neonicotinoid pesticide reduces bumblebee colony growth and queen production.” Science 336(6079): 351-352.
Williams, G. R., D. R. Tarpy, D. VanEngelsdorp, M. P. Chauzat, D. L. Cox-Foster, K. S. Delaplane, P. Neumann, J. S. Pettis, R. E. L. Rogers and D. Shutler (2010). “Colony Collapse Disorder in context.” Bioessays 32(10): 845-846.
Williamson, S. M. and G. A. Wright (2013). “Exposure to multiple cholinergic pesticides impairs olfactory learning and memory in honeybees.” Journal of Experimental Biology 216(10): 1799-1807.
Winfree, R., N. M. Williams, J. Dushoff and C. Kremen (2007). “Native bees provide insurance against ongoing honey bee losses.” Ecology Letters 10(11): 1105-1113.
Yang, E. C., Y. C. Chuang, L. Chen and L. H. Chang (2008). “Abnormal foraging behavior induced by sublethal dosage of imidacloprid in the honey bee (Hymenoptera: Apidae).” Journal of Economic Entomology 101: 1743–1748.
Acknowledgments
We acknowledge the beekeepers who took our survey as well as Jordan Levinson and Chia-Yi Hou of EcoHealth Alliance for assistance. Research was supported by a National Science Foundation Human and Social Dynamics “Agents of Change” award (SES-HSD-AOC “Human-related factors affecting emerging infectious diseases,” BCS—0826779), and by funding from the MARS Foundation, the Margaret A. Cargill Foundation, and Bayer Crop Science. Funders had no role in the data collection, analysis, nor in the preparation of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Smith, K.M., Loh, E.H., Rostal, M.K. et al. Pathogens, Pests, and Economics: Drivers of Honey Bee Colony Declines and Losses. EcoHealth 10, 434–445 (2013). https://doi.org/10.1007/s10393-013-0870-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10393-013-0870-2