
Abstract
Ethnobotanical surveys were conducted to locate culturally important, regionally scarce, and disappearing medicinal plants via a novel participatory methodology which involves healer-expert knowledge in interactive spatial modeling to prioritize conservation efforts and thus facilitate health promotion via medicinal plant resource sustained availability. These surveys, conducted in the Maya Mountains, Belize, generate ethnobotanical, ecological, and geospatial data on species which are used by Q’eqchi’ Maya healers in practice. Several of these mountainous species are regionally scarce and the healers are expressing difficulties in finding them for use in promotion of community health and wellness. Based on healers’ input, zones of highest probability for locating regionally scarce, disappearing, and culturally important plants in their ecosystem niches can be facilitated by interactive modeling. In the present study, this is begun by choosing three representative species to train an interactive predictive model. Model accuracy was then assessed statistically by testing for independence between predicted occurrence and actual occurrence of medicinal plants. A high level of accuracy was achieved using a small set of exemplar data. This work demonstrates the potential of combining ethnobotany and botanical spatial information with indigenous ecosystems concepts and Q’eqchi’ Maya healing knowledge via predictive modeling. Through this approach, we may identify regions where species are located and accordingly promote for prioritization and application of in situ and ex situ conservation strategies to protect them. This represents a significant step toward facilitating sustained culturally relative health promotion as well as overall enhanced ecological integrity to the region and the earth.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
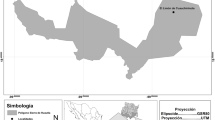
The Maya Mountains of Belize, Central America (Figure 1), support a large area of varied tropical rainforest ecosystems. Given model conservation practices, supportive natural history, and local ecosystem views, the rainforests are well intact and diverse in the remote and rugged, rainforest-laden Maya Mountains range of southern Belize. Indeed, they are host to some of the most intact regions of tropical rainforest in all of Central America (Meerman and Clabaugh, 2007). Floral diversity of the area is promoted by a multitude of environmental features inclusive of high levels of regional precipitation (Hartshorn et al., 1984; Balick et al., 2001; Pesek et al., 2006a; Meerman and Clabaugh, 2007) and unique geomorphologic variation, including elevation differentials (Dixon, 1956; Bateson and Hall, 1977; Abramiuk and Meurer, 2006).

Location of the Maya Mountains study site within the southeastern Maya area.
There are four predominant floral ecosystem types in the Maya Mountains region (Meerman and Clabaugh, 2007). They rise in the southern half of Belize and extend roughly 60 miles in a southwest–northeast direction and then extend westward at the northernmost end (Hartshorn et al., 1984; Balick et al., 2001). The range is predominantly granitic-volcanic and it ascends steeply from the coastal limestone plains which border it to the east. The chain peaks around 1000 m and then descends to a western plateau of generally 500 m in elevation. The prevailing winds from the Caribbean drop their precipitation on the southern slopes, contributing much to the floristic diversity and species differential between the northern and southern slopes (Balick et al., 2001). Annual rainfall ranges from approximately 431.8 cm (170 inches) in the south to 127 cm (50 inches) in the north (Hartshorn et al., 1984; Balick et al., 2001). North of the divide, there are two predominant ecosystems of the mountains: lowland broad-leaved moist forest and submontane broad-leaved moist forest. The regions to the south of the divide are dominated by the other two of the four main floral ecosystem types of the Maya Mountains: submontane broad-leaved wet forest and lowland broad-leaved wet forest (Meerman and Clabaugh, 2007). As exemplified by the above precipitation differential, there is a major difference between the northern and southern ecosystems; it is that of greater rainfall and correlative supported floristic diversity within the southern ecosystem types. The present study focuses effort on the last of the three floral ecosystems listed above. These are the southernmost three.
Several recent surveys of the mountains have produced new records of botanical species which may well include those new to science (Ironmonger et al., 1995; Pesek et al., 2006a). A biodiversity hotspot (Myers et al., 2000), the remote and rugged nature of the Maya Mountains also provides for rich cultural traditions in the area. This finding is in keeping with the fact that traditional cultures persist most notably internationally in mountainous regions (Stepp et al., 2005). This intact cultural diversity is host to a rich, intact traditional healing system among the indigenous Q’eqchi’ Maya of the southern Maya Mountains (Treyvaud-Amiguet et al., 2005).
In exemplary fashion, the Government of Belize has entered into an agreement for comanagement of the area with the Maya inhabitants. This partnership supports sustainable land use and management, a much needed and innovative exercise of import given our global need for answers to complex questions on conservation of biological and cultural diversity. It is a well-known fact that the biodiversity of earth is being lost at a staggering rate (World Wildlife Fund, 2004; Hanski, 2005), and that the rate of destruction of our rainforest ecosystems is unsustainable (United Nations Food and Agricultural Organization, 2000). It is also well-known that the destructive operators arise from complex human need in resource exploitation and exchange, inclusive of economic underpinnings. Additionally, cultural diversity is disappearing at an alarming rate, along with the areas that nurtured these learned patterns of behavior.
Languages can be taken as a marker of cultural diversity and indigenous peoples knowledge inclusive of traditional healing, ecosystems concepts, and an understanding of nature’s utility and natural harmony. Language is disappearing along with the loss of the world’s natural areas and biodiversity. Language diversity trends can be extrapolated to quantify the disappearance of cultural diversity (Maffi, 2001a, b; Buenz, 2005; Sutherland, 2003). Globally, languages are disappearing quickly. Of the 15,000 languages spoken 70 years ago, only 6000 are still spoken today (Grimes, 1996; Davis, 1999; Maffi, 2001a; Buenz, 2005; Sutherland, 2003).
It has been advanced by Jeffrey McNeely, chief scientist, IUCN, that preservation of tropical forests and the cultures which call them home can only be accomplished together. One strategy with much promise in accomplishing this is strategic application of certain types of traditional knowledge, for example, the promotion of respect and select integration of traditional healing in national health care systems (World Health Organization, 2002; Arnason et al., 2004a, b; Pesek et al., 2006a, 2007).
Traditional knowledge
The main form of knowledge to which people ascribe is knowledge based on observation and experience through engaging with the environment (Conklin, 1954; Berlin, 1992). This engagement with the environment can consist of formulaic, systematic empirical testing, which is a trademark of the scientific method, or it can constitute a form of testing that is less formulaic, but based on observation, and is systematic nonetheless. The knowledge gained via these means goes by many names, including common knowledge, cultural knowledge, and traditional knowledge.
Traditional knowledge, however it is acquired, has one critical feature that it shares throughout the world, and that is that traditional knowledge has been tested. This is especially apparent in the world of plants and the human uses of plants. Much of our traditional knowledge of plants and their uses comes from generations of trials involving human subjects who came into contact with these plants in one way or another. Some of this traditional botanical knowledge stretches several millennia, as the case with the Maya has shown.
Our knowledge web and how it is affected
Traditional knowledge is integral to human adaptation to the environment. In order to understand better how traditional knowledge is vital to our survival, we must be clear on the fundamental meaning of knowledge. Although the topic of knowledge has been one of the most controversial topics in Western philosophy, from a cognitive psychological perspective, knowledge can best be defined as a series of mental concepts and their interrelationships which are stored in memory.
Knowledge is affected through association and disassociation of concepts; this goes for much of our memory in general. Sever the interrelationships between one concept and another, and one of these concepts will no longer be accessible through the other. Constantly retrieve these concepts from memory together and the neurophysiological underpinnings of the interrelationship between concepts will be strengthened (Hebb, 1949).
Many mental concepts have external counterparts, that is, we often have stored in memory mental concepts of material things in the environment. With regard to concepts with external counterparts, these concepts are often accessed by being prompted by their external counterparts. Cross-cultural tests of the knowledge bases of Tzeltal Maya children and American children support this view. The disparities between the knowledge bases of Tzeltal Maya children and American children were attributed to exposure to different plants and animals (Dougherty, 1978). This research suggests that our knowledge is in large part structured information pervading the local environment, external to our neurophysiological wiring and functioning as a kind of memory store. This, of course, is a compelling argument against destroying the environment since it suggests that by destroying the environment, we are in effect erasing our external memory stores—and with it our knowledge.
Q’eqchi’ Maya traditional healing knowledge
For varied reasons, including remoteness of mountainous setting and the intact nature of regional rainforests, components of Q’eqchi’ Maya traditional knowledge continue to be maintained. Not only do the Q’eqchi’ maintain an intact traditional healing system to this day (Treyvaud-Amiguet et al., 2005), they are proactive in the conservation of this rich heritage, as well as the interdependent biodiversities (Arnason et al., 2004a, b; Pesek et al., 2006a 2007). The reasons for this include aforementioned effectors, as well as the fact that Q’eqchi’ Maya populations preferentially gravitate to traditional healers for healthcare and so their healers actively access their knowledge within their traditional ecosystems settings. These healers serve as primary care providers, very much in keeping with global estimates that 80% of the world’s population relies on traditional healers for primary healthcare (Farnsworth et al., 1985; World Health Organization, 2002).
The material counterparts of Maya medicinal plant concepts are disappearing (i.e., the medicinal plants and rainforest niches themselves), and thus, so is the knowledge associated with these plants, the other plants with which they are associated in the wild, and the sustainable uses to which these plants could be put (e.g., culturally relative health promotion).
The Maya have acquired a heritage that goes back more than three millennia. We see evidence of a distinctive cultural tradition brewing in the Maya lowlands as early as the Middle Formative period at such sites as Cuello, in what is now Belize (Housley et al., 1991; Hammond et al., 1992). For the ancient Maya of the lowlands, knowledge of the local plant life and how the Maya conceptualized their environment was essential for their survival. For thousands of years, they acquired, by necessity, a knowledge base in this bountiful rainforest region, and today, we must learn from these traditions, and others, in constructive ways.
Medicinal plants and maya concepts of medicinal plants
According to researchers in the cognitive sciences, humans like many other animals maintain cognitive maps of the environment in which they move (Hirtle and Jonides, 1985; Herman et al., 1987). For the past and present Maya, these cognitive maps would have represented the regions in the natural environment in which they collected their medicinal plants. In other words, cognitive maps are assumed to have functioned as concepts underwriting the traditional knowledge web of the Maya, at least regarding where specific medicinal plants can be found. The Q’eqchi’ Maya healers, for example, go precisely to the area where they remember there being specific medicinal plants or areas where they understand certain plants may be present. In fact, they view their surroundings with regard to traditional ecosystems concepts and then use these concepts in navigating as they collect medicinal plants. Specifically, the Q’eqchi’ Maya healers note four environmental categories in which medicinal plants grow: cool areas under high forest with much humus, warm areas under low forest, hot and warm areas on rocks and cliffs, and riversides. These concepts have profound implications on a variety of fronts, including: subsistence and use patterns, sustainable resource exploitation and resource exchange practices, propagation of knowledge, targeted conservation, and culturally relative health promotion.
The notion of a cognitive map is a very useful way of representing the concepts by which the Maya, past and present, move through the natural environment to their medicinal plant destinations. As such, we use the notion of a cognitive map as an heuristic to account for the concepts that make up Q’eqchi’ Maya traditional knowledge regarding where medicinal plants are located.
Propagation of concepts of medicinal plants
Research into understanding how concepts are acquired or, in our specific case, how cognitive maps are acquired is ongoing. Whereas there is no clear understanding of this process, some researchers have proposed underlying processes and mechanisms for explaining the manner in which concepts propagate (Sperber, 1979; Lake, 1998).
Thus far, the hypothesis accounting for the propagation of mental concepts which has been researched the most extensively is the cultural evolutionary approach (Shennan, 2002). Here, concepts propagate through populations of human individuals much like infectious diseases. Indeed, models that are used to account for how “ideas” spread are based on epidemiological models (Sperber, 1979; Cullen, 2000). In this scenario, a concept, or in our case a cognitive map, is subject to the same evolutionary laws by which biological organisms abide.
Biological evolution is a change through time; this change is caused by certain traits that are selected due to their role in increasing the fitness of the individual organisms in a given population. If laws governing how mental concepts change and gain priority status in the human mind are similar to the laws governing biological evolution, then we can use mechanisms for evolution, namely natural selection and mutation, to approximate the mechanisms by which mental concepts pervade the minds of human individuals. Based on this premise, in the present study, we rely on an evolutionary computational technique that reproduces the process by which cognitive maps can be seen to form in the human mind. These cognitive maps are expressed by displaying them externally so that individuals, such as the Q’eqchi’ Maya healers that are conceiving of them, can manipulate them in a manner according to their understanding. We then can use these “externalized” cognitive maps to predict plant locations. These can then serve to structure and prioritize community-based conservation initiatives and sustainable resource management scenarios—in effect, culturally relative conservation of both culture and biodiversity—and this then enables the continuance of culturally relative health promotion while facilitating greater ecological integrity.
Interactive spatial evolutionary computation-based predictive modeling of medicinal plant occurrences
Modeling the geographical distribution of a medicinal plant species is a key issue when considering its conservation and sustainable use. To gain an understanding of the distribution of medicinal plants in a specific topographic landscape, certain obvious facts regarding the species must be carefully and accurately assessed—for example, locations where the species exists or could exist, areas where they could thrive, and what specifically are their requirements for survival. Often, for a threatened plant species, this type of conceptual knowledge is embedded in the cultural knowledge of indigenous people of a particular region.
Exploring relationships between traditional knowledge, ecosystems, geospatial particulars, mathematical modeling techniques, and empirical observations for predicting species distribution is complex. Inferential procedures that provide robust and reliable predictions of species geographic distributions thus become critical to biodiversity analyses, conservation, and sustainable resource use planning. In the present work, a habitat suitability model for medicinal plant species is developed by combining spatial predictive modeling with an evolutionary computation method known as Interactive Evolution (IE) (Takagi, 2001). A variation on genetic algorithms (Holland, 1975), IE maximizes interactive spatial evolutionary computation-based predictive modeling by enabling an analyst to combine advanced mathematical geospatial and pattern recognition modeling techniques, conceptual knowledge, available empirical data and expert-interactive dynamic data visualization techniques to predict species distributions.
As a result, IE is not only a robust alternative to conventional modeling methods (Davis and Goetz 1990; Cressie, 1993; Carpenter et al., 1997; Guisan and Zimmermann, 2000; Royle et al., 2001), but an improvement upon them given the application of an interface of conceptual knowledge (i.e., expert input and shaping via varied intellects, worldviews, and cognitive maps, e.g., those of Q’eqchi’ Maya healers).
IE is particularly well suited to mathematical modeling problems whose potential solutions can be visually expressed, as is the case with spatial data analysis. It makes use of the fact that the positions of particular attribute values on a map induce associative processes in the analyst, thereby allowing the analyst to draw upon analogies, cultural and traditional knowledge, and in particular their cognitive maps.
While traditional genetic algorithms (Holland, 1975) and many other mathematical optimization techniques in spatial data analysis use an explicit analytic expression for a fitness (utility) function to be evaluated by the computer, with IE, the analyst performs this step based on visual perception and association. The strength of IE is that the user does not have to state or even understand an explicit fitness criterion—the user needs only to be able to apply it. This also frees the user from user specifications, design efforts, or having to know the algorithmic details. This feature of IE combined with the power of the geospatial mathematical modeling techniques can be used very effectively in predicting and testing hypotheses about species distributions and habitat specificity.
Using IE, the main features of habitat suitability characteristics for each plant species can be seen to represent a chromosome. Moreover, each chromosome can be seen to be comprised of genes, denoted by specific traits (features) of the species distribution. For example, a gene can be defined by an area where the species has been observed and recorded, as well as by a set of possible locations where the species can occur. Rather than provide a stringent mathematical derivation of these possibilities, however, these possibilities will be based on the analyst’s understanding, as well as on a set of environmental variables for the region of interest (e.g., precipitation, elevation, soil type, temperature, moisture content, humus, co-occurring species, etc.). Then for each chromosome, we calculate the species activity areas using density estimation, such as kernel density estimation or mixture modeling adapted from the theory of spatial point processes (Cressie, 1993).
The estimated spatial densities are then projected onto identical geographical maps of the same region, each map having a unique probability distribution of the plant species. These maps are represented by contour maps where the color/grayscale and distribution shape denote the probabilities of locating plant species. These maps provide a common language which can allow ethnobotanists, ecologists, and other expert-users (e.g., Q’eqchi’ Maya healers) to understand one another more comprehensively. Moreover, they can then be used for postulating hypotheses regarding preferences as well as assessments regarding the predictability of the models.
Field work methodology
Our study methodologies and Informed Consent protocol were approved by the Institutional Review Board of Cleveland State University, Cleveland, Ohio. This research was undertaken with Informed Consent of the Maya participants whose identities are not revealed due to ethical guidelines in research. This research was carried out with the much appreciated support of the Government of Belize via permits granted by the Forest Department. Additionally, as has been the case with our prior work, in order to provide intellectual property protection, we do not discuss specific plant species along with their use associations.
The Informed Consent and support of the healers and locals guides was sought through a series of workshops initiated by Belize Indigenous Training Institute (BITI). BITI, a Belizean NGO incorporated in 1998 by indigenous groups with the assistance of the Inuit Circumpolar Conference (Inuit Canada, Greenland, Alaska, and Siberia), is governed by a Board of Directors comprised of representatives from indigenous peoples’ cultural councils including Q’eqchi’ Council of Belize, the Toledo Maya Cultural Council, the National Garifuna Council, and the Xunantunich Organization. The organization provides practical training in areas that will lead to income generation and employment at the community level, as well as provision of capacity building training and work in the area of traditional knowledge and cultural heritage. The Q’eqchi’ Healers Association (QHA), an organized group of traditional Q’eqchi’ Maya healers, is one of the community associations affiliated with the umbrella organization BITI.
Our expedition team varied in number and specialty over the course of our work, but each expedition consisted of a core team of participatory researchers including an ethnobotanist, an ethnographer of Maya descent, three Q’eqchi’ Maya bushmasters, two representative and well-known senior practicing Q’eqchi’ Maya traditional healers and spiritual guides (four other healers joined us often, however, these two healers on our core team were on every expedition), and a navigator and mapping specialist.
The healers and bushmasters input was implemented in the development of objectives at the outset. They were trained in map interpretation and issues such as site destinations and site monitoring. Provisional maps of the natural vegetation communities using remotely sensed data were composed and evaluated for routes and vegetative overlays. The principal mapping tools employed early on consisted of existing topographic, ecosystem, and vegetation maps; remote sensing imagery, primarily Landsat Thematic Mapper and aerial photographs; DTED; and GIS data. From these and other sources, a preliminary vegetation classification of the habitat was developed.
The study area in which the ethnobotanical surveys were conducted and examined for medicinal plants were within the aforementioned remote and rugged, heavily forested region of the Maya Mountains in Belize 16°S, 17° N, −88°E, −89.25°W (Figure 1). The study was carried out over the course of 4 years, which were comprised of a spring and fall field stint ranging from 2 to 4 weeks, and then followed up with data analysis and processing.
The indigenous ecosystems concepts of the Q’eqchi’ Maya were utilized for selecting regions to collect medicinal plants. These data were collected in a spatially nonrandom scheme by letting the Maya healers (domain experts) choose specific locations within the region, which they felt were deserving of sampling. The healers were asked to move freely though portions of the region until they found locations where they would traditionally collect medicinal plants. After these positions had been located by the healers, the healers then identified the different plants around the area in 30 m × 30 m transect grids. Within these grids, when specific plants of focus were identified, 1 m × 10 m transect grids were analyzed around them in detail for environmental descriptors used in computing species distribution for each grid cell detailed below. For the present study, we continued our research focus on the 53 species of medicinal plants elucidated on our Rapid Ethnobotanical Survey (RES) (Pesek et al., 2006a) and ultimately developed detailed ecological and geospatial data on three of these species over a total of 11 transects. These three species were selected as representative of the Q’eqchi’ healing plants for the following reasons: (i) they were among those initially determined by the healers to be regionally scarce and disappearing (Pesek et al., 2006a); (ii) they were noted by the healers to be particularly important plants; (iii) there was at least one species from each of their two caloric categories (i.e., hot and cold); (iv) they rely on these plants heavily in practice (i.e., used in treatment of commonly seen ailments or used to treat specific conditions for which it is the most or only efficacious remedy) (Table 1); (v) they represent a diversity in habitat preference; (vi) they had a relative frequency significant for data collections.
Covariate data and applications methodology
In order to quantitatively characterize Q’eqchi’ Maya ecosystems concepts so that we could incorporate these concepts into our predictive model of plant spatial occurrence, we used several representative environmental descriptors that influence the patterns of occurrence of a given medicinal plant. Environmental descriptors in the form of raster-based covariates were used to model plant occurrence in the study area, and they were obtained from two main sources: (1) Digital Terrain Elevation Data (DTED) Level 1, obtained from the National Imagery and Mapping Agency (NIMA); and (2) Landsat 5 Thematic Mapper images, with a pixel size of 30 m × 30 m and with band combination 4, 5, 3, from the Biodiversity & Environmental Resource Data System (BERDS) repository (Meerman and Clabaugh, 2007). This band combination mapping allows for the strongest contrast between vegetative variants as follows: forested land (shown in shades of orange-brown), herbaceous formations (shown in shades of green), scarcely covered soils, such as plowed fields, lava streams, or bare rocks (represented by different shades of blue) (Meerman and Clabaugh, 2007). In addition to the raster-based covariates, we also used vector-based covariates. These vector-based covariates, also obtained from the BERDS repository (Meerman and Clabaugh, 2007), describe aquatic ecosystem types, UNESCO classified ecosystem types, and geologic land types.
All raster- and vector-based covariates were rasterized or aggregated to a grid cell size of 30 m2 of the DTED Level 1. Each environmental descriptor was then defined over the remaining area—a 256 × 256 grid. Since our fieldwork was localized in the mountains, grid cells belonging to the coastal and littoral water region of Belize were removed, resulting in approximately 42,000 prediction site possibilities.
Experimental design and results
For application of our model, we chose the three representative species of medicinal plants on which we amassed detailed ecological and geospatial data; these species are further represented as Medicinal Plant (MP) 6, MP 22, and MP 25 (Table 2). The data locations are sparse for these plant species, suggesting that they are regionally rare—this finding is consistent with the healers initial reports. Indeed, the prediction domain for these species make up only 0.11% of the prediction grid.
The environmental descriptors used in computing species distribution for each grid cell are: (1) terrain-related features: (i) location of grid cells—northing and easting, (ii) elevation, (iii) slope, (iv) aspect; (v) three spectral variables, the digital values of Thematic Mapper Bands 4, 5, and 3; and (2) binary features derived by examining 10 unique UNESCO-recognized ecosystems (Meerman and Clabaugh, 2007), 4 Q’eqchi’ Maya ecosystems, and 10 geologic ecosystems (Meerman and Clabaugh, 2007). The binary features are equal to 1 if the grid cell belongs to patches of land associated with the above-listed ecosystem types, and 0 otherwise.
Thirty-two environmental descriptors were used to train the predictive model. For each grid cell in the data set where one of the three medicinal plant species was found, we measured the values of all 32 environmental descriptors. This means that each medicinal plant now had an associated set of 32 distinctive measurements. This set of values could then function as a kind of “address” that we could use to locate sets of values in other grid cells with medicinal plants. The locations of the grids with medicinal plants were generated using a supervised learning algorithm and were reproduced probabilistically on a map. The probability intensity ranges anywhere from 0 to 1 (1 being 100% probability of species occurrence).
The model accuracy was assessed statistically by testing for independence between predicted occurrence and actual occurrence using cross-validation (Davis and Goetz 1990). Approximately 60% of the original data were randomly withheld in an iterative fashion, and the model output was aggregated to test for independence. The probability of correctly classifying each of the medicinal plants was then calculated. For the MP25 plant, the model correctly classified 72% of the test sites; for the MP22 plant, it classified 63% of the test sites correctly; and for the MP6 plant, it correctly classified 61% of the sites.
The probabilistic map of the species occurrence (Figure 2) accurately predicted 69% of the field observations and was able to avoid major classification distortions caused by the northeastern region and coastal areas. These are areas of mangrove and littoral forest as well as lowland savannah, which are not suitable habitat for the three species of medicinal plants considered.
A number of well-known spatial pattern recognition techniques such as Bayes decision theory, neural networks, maximum entropy, or classification trees can be used to produce probabilistic maps of species occurrence (Davis and Goetz, 1990; Carpenter et al., 1997; Guisan and Zimmermann, 2000; Royle et al., 2001). However, none of these approaches can be used to easily integrate a domain user’s knowledge into the modeling feedback loop—something we do with IE and the healer-expert users. Nevertheless, the existing approaches were used as a first step in the present predictive modeling approach. Indeed, with some modifications, we found the neural network-based approach known as adaptive resonance theory (Carpenter et al., 1997) particularly useful in constructing our supervised learning algorithm. One modification was to evolve the parameters of the basic predictive modeling algorithm (e.g., parameters that influence learning ability and learning rate of classification). Another modification was to select and modify the environmental descriptors of the training data, within ranges that reflect the indigenous conceptions of plant habitat, so that probabilistic maps can be produced of species occurrences which coincide with that of the healers’ cognitive maps. For example, according to the healers, MP22 is found on hillsides (i.e., a slope we deem to be between 10° and 30°) facing north to northeast (i.e., an aspect we deem to be between 0° and 67°) within one of their four ecosystem types. Such flexibility allows us to search for nonrandom associations between environmental descriptors of medicinal plant loci of known occurrence all over the study area.
The computational model devised and generated multiple maps displaying the probabilities of species occurrence (Figure 3). These maps display probabilities of species occurrence which are then subjectively evaluated for merging by domain users according to their conceptions of the species ecology and known characteristics of species habitat (their expert understanding). This merging is accomplished by picking the maps that appear closest to how the domain user envisions the terrain. By choosing the maps, the domain expert is combining the attributes of the maps in a manner not unlike what we have described for the manner in which cognitive maps are created in the human mind. Consistent with the view that cognitive maps are subject to evolutionary principles just as biological organisms, the expressions of these cognitive maps in the form of the computer-generated maps are treated as if they are individuals of a reproductive pair. In selecting the maps to be combined, the expert user is in effect “mating” the maps. The outcome of this crossover produces a new line of maps. The algorithm iterations are repeated until the user is satisfied that one of the maps displaying the probabilities of plant occurrences resembles the one the user has envisaged (Figure 2). In addition, during the process of map evaluations and selections, the domain user can directly enhance the algorithm accuracy by allowing the expert user to interact with each of probabilistic maps by including or excluding pixels or region(s) on the maps where they believe species do or do not exist. That is, the predictive model can not only learn from positive examples (e.g., data presently available) but can also learn from negative examples—loci where user believes (knows) the medicinal plants either can exist and have not been predicted there or cannot exist but have been predicted there. This additional modification generated by the expert during the model run serves to increase the accuracy of the model so that the probabilities of correct classification are increased.

Probabilistic map of three medicinal plant species (MP 6 light gray, MP 22 dark gray, MP 25 black) occurrence after 13 generations of algorithm runs and combined with the expert users evaluations of the spatial predictive models of the species and associated corrections to these models.

IE-based spatial predictive modeling tool display. The user is presented with four possible outcomes of the basic spatial predictive model of distributions of three species of medicinal plants (MP 6 light gray, MP 22 dark gray, MP 25 black). No predictions were made where habitat characteristics differed from those encountered at training sites. The display grid can be easily reconfigured to accommodate for more than four probabilistic maps of species occurrences to be evaluated by an expert user. The expert user selects a model or multiple models to survive into the next generation based on varied nonquantifiable measures like traditional knowledge, visual or intuitive cues, or other experience. The automatic generation of variation is accomplished by mutation and recombination operators that constantly propose new variants to the presently existing population of spatial predictive models on the screen.
Discussion and future directions
We can effectively learn from the traditional knowledge of indigenous peoples (Pesek et al., 2006b), and this, in turn, has untold broad impacts (Arnason et al., 2004a, b; Pesek et al., 2006a, 2007). The present study is a step toward identifying regions where certain medicinal plants are likely to be, and to argue for the appropriate prioritization in applying conservation strategies based on accurate computational output of traditional ecosystems concepts. We can work with the healers to better record and understand their broad notions, such as ecosystems concepts and the very basis of their knowledge to more specific venues, e.g., their pharmacopoeias, the biological activity of their floras, and the cultural context of their healing. These activities have potential to generate fresh revenue streams for the local Maya and Belize, and for conservation of biodiversity and culture via culturally relative methods. At the same time, they will aid in disseminating plant healing knowledge to younger generations of Maya before it is lost, as well as, via concurrent programming, effect culturally relative wellness promotion and community health. Innovative applications in conservation can also alleviate the pressure on harvesting plants from the wild. Plants and their healing properties make up a significant part of Maya culture; when these plants disappear, so do the cultural traditions that once utilized the plants.
As ethnobotanical, ecological, and spatial data are accrued in the course of our fieldwork, there will be new information and ecosystems services data being continually refined in our reference collection and in our predictive model. Our findings can be applied in different ways, including in terms of ecosystems services in support of culturally relative conservation arguments which host significant broad impacts in terms of ecosystems health, culturally relative healthcare, and the development of a model in sustainability for us as a global people. At the same time, we can use these applications in facilitating both in situ and ex situ conservation techniques, and sustainably cultivate these plants according to traditions and ecologies.
References
Abramiuk MA, Meurer W (2006) A preliminary geoarchaeological examination of ground stone artifacts in and around the Maya Mountains, Toledo District, Belize. Latin American Antiquity 17:335–354.
Arnason J, Cal V, Assinewe V, Poveda L, Waldram J, Cameron S, et al. (2004a) Visioning Our Traditional Health Care: Workshop on Q’eqchi’ Healers Center, Botanical Garden and Medicinal Plant Biodiversity Project in Southern Belize. Final report to IDRC, Ottawa, Canada, 48 pp
Arnason J, Catling P, Small E, Dang P, Lambert J (editors) (2004b) Biodiversity & Health: Focusing Research to Policy, Ottawa, Canada: NRC Research Press.
Balick M, Nee M, Atha D (2001) Checklist of Vascular Plants of Belize, with Common Names and Uses, New York: New York Botanical Garden Press.
Bateson J, Hall I (1977) The Geology of the Maya Mountains, Belize (Overseas Memoir 3), London, UK: Her Majesty’s Stationery Office.
Berlin B (1992) Ethnobiological Classification: Principles of Categorization of Plants and Animals in Traditional Societies, Princeton, NJ: Princeton University Press.
Buenz E (2005) Country development does not presuppose the loss of forest resources for traditional medicine use. Journal of Ethnopharmacology 100:118–123.
Carpenter GA, Gjaja MN, Gopal S, Woodcock CE (1997) ART neural networks for remote sensing: vegetation classification from Landsat TM and terrain data. IEEE Transactions on Geoscience and Remote Sensing 35:308–325.
Conklin H (1954) The Relation of Hanunóo Culture to the Plant World. Unpublished PhD dissertation in anthropology, New Haven, CT: Yale University.
Cressie N (1993) Statistics for Spatial Data: Revised Edition, New York: John Wiley and Sons.
Cullen BS (2000) Contagious Ideas: On Evolution, Culture, Archaeology, and Cultural Virus Theory Oxford, UK: Oxbow Books.
Davis W (1999) Clouded Leopard, Vancouver, BC: Douglas and MacIntyre Publishing Group.
Davis F, Goetz S (1990) Modeling vegetation pattern using digital terrain data. Ecology 4:69–80.
Dixon C (1956) Geology of Southern British Honduras with Notes on Adjacent Areas, Belize City, Belize: Government Printing Office.
Dougherty J (1978) Salience and relativity in classification. American Ethnologist 5:66–79.
Farnsworth N, Akerele O, Bingel A, Soejarto D, Guo Z (1985) Medicinal plants in therapy. Bulletin of the World Health Organization 63:965–981.
Grimes B (1996) Ethnologue: Languages of the World, 13th ed., Dallas: Summer Institute of Linguistics.
Guisan A, Zimmermann N (2000) Predictive habitat distribution models in ecology. Ecological Modelling 135:147–186.
Hammond N, Clarke A, Belli FE (1992) Middle preclassic Maya buildings and burials at Cuello, Belize. Antiquity 66:955–964.
Hanski I (2005) Landscape fragmentation, biodiversity loss and the societal response. EMBO Reports 6:388–392.
Hartshorn G, Nicolait L, Hartshorne L, Bevier G, Brightman R, Cal J, et al. (1984) Belize, Country Environmental Profile, Trejos Hnos, San Jose, Costa Rica
Hebb DO (1949) The Organization of Behavior: a Neuropsychological Theory, London: Chapman and Hall.
Herman JF, Blomquist SL, Klein CA (1987) Children’s and adults cognitive maps of very large unfamiliar environments. British Journal of Developmental Psychology 5:61–72.
Hirtle SC, Jonides J (1985) Evidence of hierarchies in cognitive maps. Memory and Cognition 3:208–217.
Holland JH (1975) Adaptation in Natural and Artificial Systems, Ann Arbor, MI: University of Michigan Press.
Housley RA, Hammond N, Law IA (1991) AMS radiocarbon dating of preclassic Maya burials at Cuello, Belize. American Antiquity 56:514–519.
Ironmonger S, Leisnerm R, Sayre R (1995) Plant records from the natural forest communities in the Bladen Nature Reserve, Maya Mountains, Belize. Caribbean Journal of Science 31:30–48.
Lake MW (1998) Digging for memes: the role of material objects in cultural evolution. In: Cognition and Material Culture: The Archaeology of Symbolic Storage, Renfrew C, Scarre C (editors), Cambridge, UK: University of Cambridge, McDonald Institute for Archaeological Research, pp 77–88.
Maffi L (2001a) Language, knowledge, and indigenous heritage rights. In: On Biological and Cultural Diversity, Maffi L (editor), Washington, DC and London, UK: Smithsonian Institution Press, pp 412–432.
Maffi L (editor) (2001b) On Biological and Cultural Diversity, Washington, DC and London, UK: Smithsonian Institution Press.
Meerman JC, Clabaugh J (editors) (2007) Biodiversity & Environmental Resource Data System (BERDS). Available: http://www.biodiversity.bz (v. October 2007)
Myers N, Mittermeier R, Mittermeier C, da Fonseca G, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858.
Pesek T, Cal M, Cal V, Fini N, Minty C, Dunham P, et al. (2006a) Rapid ethnobotanical survey of the Maya Mountains range in southern Belize: a pilot study. Trees for Life Journal 1:1–12.
Pesek T, Helton L, Nair M (2006b) Healing across cultures: learning from traditions. EcoHealth Journal 3:114–118.
Pesek T, Cal V, Fini N, Cal M, Rojas M, Sanchez P, et al. (2007) Itzama: revival of traditional healing by the Q’eqchi’ Maya of southern Belize. HerbalGram 76:34–43.
Royle J, Link W, Sauer J (2001) Statistical mapping of count survey data. In: Predicting Species Occurrences: Issues of Scale and Accuracy, Covello, CA: Island Press
Shennan S (2002) Genes, Memes, and Human History: Darwinian Archaeology and Cultural Evolution, London: Thames and Hudson Ltd.
Sperber D (1979) Explaining Culture: A Naturalistic Approach, Oxford, UK: Blackwell Publishers Ltd.
Stepp J, Castaneda H, Cervone S (2005) Mountains and biocultural diversity. Mountain Research and Development 25:223–227.
Sutherland W (2003) Parallel extinction risk and global distribution of languages and species. Nature 423:276–279.
Takagi H (2001) Interactive evolutionary computation: fusion of the capacities of EC optimization and human evaluation. Proceedings of the IEEE 89:1275 –1296.
Treyvaud-Amiguet V, Arnason J, Maquin P, Cal V, Sanchez-Vindaz P, Poveda L (2005) A consensus ethnobotany of the Q’eqchi’ Maya of southern Belize. Economic Botany 59:29–42.
United Nations Food and Agricultural Organization (2000) Global Forest Resources Assessment, 2000. FAO Forestry Paper 140. http://www.fao.org/forestry/fo/fra/main/ (accessed August 3, 2005)
World Health Organization (2002) WHO Traditional Medicine Strategy 2002–2005, Geneva, Switzerland: World Health Organization.
World Wildlife Fund (2004) Living Planet Report 2004, Gland, Switzerland: World Wildlife Fund.
Acknowledgments
This work was supported by Cleveland State University, College of Science, and the Naturaleza Foundation.
Author information
Authors and Affiliations
Corresponding author
Additional information
T.P. designed study, conducted data collections and analysis, and largely wrote the paper. M.A. advised and assisted with study design, data collections and analysis and contributed heavily to the paper in the knowledge and cognitive sections and the writing thereof. D.G. developed algorithms for spatial evolutionary computation-based predictive modeling and applied them in study design, performed spatial statistical analysis and contributed heavily to sections on interactive spatial predictive modeling, study design, results and the writing thereof. N.F. assisted with study design, data collections, analysis, and mapping. J.M. assisted with study design and covariate data. V.C. assisted with on site coordination. All authors discussed results and commented on the manuscript.
Rights and permissions
About this article
Cite this article
Pesek, T., Abramiuk, M., Garagic, D. et al. Sustaining Plants and People: Traditional Q’eqchi’ Maya Botanical Knowledge and Interactive Spatial Modeling in Prioritizing Conservation of Medicinal Plants for Culturally Relative Holistic Health Promotion. EcoHealth 6, 79–90 (2009). https://doi.org/10.1007/s10393-009-0224-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10393-009-0224-2