
Abstract
The Monte Postale and Pesciara di Bolca sites are well-known Eocene Fossil-Lagerstätten. The origin of these deposits has been previously related to a lagoon environment, protected from the open sea by a barrier formed by some kind of reef. However, no detailed description of a bioconstructed system has ever been reported in the area. This study provides the first detailed characterization of the upper Ypresian Monte Postale reef, made of in situ corals, calcareous algae, and encrusting foraminifera. Three main facies associations have been recognized: (1) a coralgal rim, roughly E/W oriented, (2) lagoon deposits in the south and (3) fore-reef sediments towards the north. The coralgal rim is represented by a wave-resistant framework made of heavily calcified organisms associated to Alveolina accumulations; the adaptation of most of these organisms to the euphotic zone promoted their growth in proximity to the sea level, allowing this structure to act as an effective barrier. In particular, during relative sea-level lowstands or highstands, the coralgal rim could have limited the water exchange between the lagoon and the open sea, reducing the oxygenation in the lagoon and allowing the formation of the Fossil-Lagerstätten. During relative sea-level transgressions, the water could have risen well above the margin, establishing normal oxygenation and hampering the fossils’ preservation. The Monte Postale succession represents one of the few examples of lower Eocene bioconstructions. The description of this biotic assemblage provides new data for the characterization of reef communities during a crucial climatic phase, near the end of the early Eocene climatic optimum global warming.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
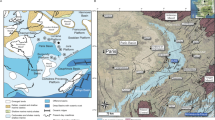
The lower Eocene Monte Postale succession is located in the eastern part of the Lessini Mountains (NE Italy), close to the Pesciara di Bolca, a worldwide famous Eocene Fossil-Lagerstätte (Fig. 1). Starting from the 16 century (Roghi et al. 2014), the Pesciara and Monte Postale localities provided a great amount of exquisitely preserved fossils. This includes mainly fishes, along with seagrass and terrestrial plant remains, reptiles, bird feathers, insects, molluscs, jellyfishes, crustaceans, worms, foraminifers, and insects (Sorbini 1972, 1999; Papazzoni et al. 2014). The ichthyofauna, extraordinarily well diversified, is represented by about 240 different taxa, and documents the earliest clearly defined coral reef fish assemblage (Bellwood 1996; Bellwood et al. 2014; Carnevale et al. 2014).

a Location map of the study area. b Close-up of the Bolca area and location of the Monte Postale and Pesciara outcrops
The Pesciara succession has been interpreted since a long time as deposited in a carbonate platform setting, within an intra-platform depression or lagoon, protected from the open sea by some kind of threshold (Sorbini 1972; Papazzoni and Trevisani 2006; Schwark et al. 2009; Trevisani 2015). Both the reef-related fish assemblage and the common occurrence of coral fragments in the Pesciara deposits suggest that this threshold may be formed by some kind of bioconstructed margin. The presence of isolated blocks of reef limestones has been briefly reported by Barbieri and Medizza (1969) from a few localities some kilometers north of the Pesciara di Bolca. In addition, a recent survey of the Monte Postale (Trevisani 2015) mentioned the presence of large benthic foraminifera–calcareous algal buildups that acted as a margin separating the Postale–Pesciara lagoon from the open sea. However, despite this evidence, no detailed description of a reef-building association, or facies analysis of reef-related sediments, have been previously reported in the Bolca surroundings.
Monte Postale represents the most complete stratigraphic record of this area (Munier-Chalmas 1891; Fabiani 1914, 1915; Dal Degan and Barbieri 2005; Trevisani 2015) and, although a valley, volcanic deposits and possibly a fault separate this succession from the nearby Pesciara (Fig. 4a), strong similarities between lithological features and fossil associations suggest that the two sites were part of the same depositional system. In order to improve and update both stratigraphical and paleontological data sets, a new geological survey of this area has been carried out. During the fieldwork, several massive to weakly stratified limestones were observed along the northern side of the Monte Postale, and their in situ position within the stratigraphic succession was ascertained. They are composed mainly of corals and calcareous algae, in growth position or preserved as broken rubble, associated to Alveolina accumulations. These sediments form a narrow, discontinuous belt, several meters thick and arranged along an E/W-directed alignment.
The main goal of this study is to provide, for the first time and within a sound biostratigraphic framework, a detailed description of the reef system associated to one of the most important Eocene Fossil-Lagerstätten in the world, and to reconstruct the environmental conditions that controlled the development of this structure. The results of this study will thus contribute to clarify the complex early Eocene paleoenvironmental setting of the Bolca area, providing the base for a more detailed paleoecological interpretation of the entire Fossil-Lagerstätten biota. Moreover, as the Monte Postale site represents one of the few examples of lower Eocene bioconstructions, the detailed analysis of the reef-building assemblage provides new information on the structure of Paleogene reefs in a time-window when these associations are poorly known (Scheibner and Speijer 2008; Zamagni et al. 2012), and during a critical climatic phase, when Cenozoic Earth surface temperatures attained their warmest state (Zachos et al. 2001; Payros et al. 2015).
Geological setting
The Lessini Mountains are located within the Southern Alps area, a structural element corresponding to the northernmost corner of the Adriatic plate (Schmid et al. 2004; Márton et al. 2011; Carminati et al. 2012; Handy et al. 2015). During the Early Jurassic, this area was occupied by a carbonate platform (the Trento Platform), confined to the West by the Lombard Basin and to the East by the Belluno Trough (Bosellini et al. 1981; Winterer and Bosellini 1981) (Fig. 2). This platform, starting from the Middle Jurassic, drowned, and became a pelagic plateau (Trento Plateau), until the Alpine collision produced a gradual uplift, with the segmentation of the plateau into several blocks. This phase was accompanied by extensive volcanic activity, possibly as a consequence of a mantle plume, with eruptive products (mainly basalts) contributing to raise parts of the sea bottom (Piccoli 1966; Barbieri et al. 1991; Zampieri 1995; Macera et al. 2008; Cannatelli 2012). Barbieri et al. (1991) recognized six volcanic stages between the late Paleocene and the middle Eocene. Some of the uplifted blocks reached the photic zone during the early Eocene and started to act as centers of initiation of shallow-water carbonates, eventually coalescing together to form a vast platform called Lessini Shelf (Fig. 2) (Doglioni and Bosellini 1987; Bosellini 1989; Luciani 1989; Bassi et al. 2008).

Paleogeographic reconstruction of the Lessini Shelf during the Paleogene (modified from Bosellini 1989). The star indicates the position of the study area
The old geological literature attributed all the lower Eocene deposits in the Lessini Mountains to an informal unit named “Calcari di Spilecco” (e.g., Malaroda in Bosellini et al. 1967; Ungaro 2001). However, in the Spilecco area, only a very short time interval is represented, namely latest Paleocene to earliest Eocene (NP9–10; Papazzoni et al. 2014). Therefore, it seems more advisable to refer most of the lower Eocene sediments in the Lessini Mountains to the Chiusole Formation (sensu Luciani 1989), made of carbonates deposited in basinal to slope settings. These deposits also contain resedimented shallow-water carbonates, whose presence gradually increases toward the raised areas of the Lessini Shelf. These shallow-water sediments have been informally indicated as “Calcari Nummulitici” (nummulitic limestones) (Bosellini et al. 1967; Carraro et al. 1969; De Zanche et al. 1977; Sarti 1980; Ungaro 2001) and in the middle Eocene they overlaid the deeper sediments of the Chiusole Formation. The Calcari Nummulitici unit, to which the Monte Postale limestones belong, is characterized by a complex geological setting. This is related to local tectonics, with a complex system of faults displacing the sedimentary successions, and to several volcanic intercalations, associated to the magmatic events that occurred during and after the carbonate deposition (Fig. 3) (Barbieri et al. 1991; Barbieri and Zampieri 1992; Zampieri 1995). Different types of lithologies have been assigned to the Calcari Nummulitici complex: limestones rich in larger foraminifera, corals and molluscs; coralline algal breccias; marly limestones and clays with freshwater and terrestrial fossils (Bosellini et al. 1967). This lithological heterogeneity and the complexity of the geological setting are the causes of a long debate on the precise definition of the Calcari Nummulitici (see for example Bosellini et al. 1967; Antonelli et al. 1990; Muscio and Tintori 2005), that still needs a thorough revision (Papazzoni et al. 2014).

Simplified geological map of the Monte Postale area; modified from Muscio and Tintori (2005)
The volcanic activity ceased during the late Eocene, a phase underlined by the deposition of deep-platform, marly sediments (Barbieri et al. 1980; Mietto 1992; Trevisani 1997). During the early Oligocene, the Lessini Shelf became a rimmed platform, as testified by a reef margin still preserved toward the eastern boundary of this area (Frost 1981; Bosellini and Trevisani 1992; Mietto 1992). During the late Oligocene, sandstones and limestones (Arenarie e Calcari di S. Urbano Formation) deposited only in the easternmost part of the Lessini Mountains (Bassi et al. 2007). The lower Miocene Marne Argillose del Monte Costi Formation constitutes the last marine deposits in the Lessini Mountains (Bosellini et al. 1967; Bassi et al. 2007, 2008; Bassi and Nebelsick 2010).
Materials and methods
The present study has been based on an extensive fieldwork, including a detailed geological survey, stratigraphic logging and sampling, together with facies analysis. Fieldwork was challenging due to the presence of a dense vegetation cover, the occurrence of minor tectonic displacements and a generally poor preservation of the outcrops.
The vegetation cover, present all over the year, considerably reduced the number and the extension of suitable outcrops. Even though, seven different sections have been measured and sampled (Fig. 4). The maximum thickness is displayed in the log E, measured on the southeastern side of the Monte Postale, facing the Pesciara outcrop (Fig. 4). Logs A1, A, B, C, D, and D1 are reduced in thickness and are related to small outcrops, maximum 15 m long and 10 m wide, arranged along the northern side of the Monte Postale (Fig. 4a). There, an alignment (approximately 180 m long and 25 m wide) of massive limestones rich in corals and calcareous algae has been identified (Fig. 4b). These coralgal limestones have been specifically selected for the study of the Monte Postale reef depositional system.

a Positions of the measured stratigraphic sections. b Measured logs with stratigraphic correlations
Although no major faults were recognized during fieldwork, several minor tectonic disturbances have been observed, related to the widespread occurrence of volcanic intrusions. Therefore, we considered the current bedding orientations and dippings of the studied outcrops as not completely reliable for the reconstruction of the original architecture of the depositional system.
Finally, the poor preservation of the outcrops greatly hampered the field observations and, in many cases, biotic components and sedimentary features have been recognized only by hammering the rock surface. To overcome this limitation, a large number of samples have been collected. From 108 samples, 16 polished sections have been prepared to integrate macroscopic field observations, whereas 132 thin-sections (6 × 4.5 cm) have been prepared for microfacies analysis.
In order to further improve the description of the reef depositional system, quantitative analyses have been applied for different purposes. To document facies distribution, not directly recognizable in the field, an average of 18 samples has been considered within each of the studied outcrops and the percentage of samples belonging to the different facies has been calculated.
Moreover, in order to document the composition of the reef-builders, relative abundances of corals were obtained considering the number of fragments or individual colonies recognized in thin-section on 64 samples. Corals have been identified at the genus level. Counts also included the solenoporacean alga Parachaetetes, which constitutes a dominant component of the reef-building biota.
In addition, to establish the diversity of the encrusting association, the percentage of each taxon has been calculated through point counting on 16 thin-sections. For each section, an average of 170 points has been considered as adequate for a reliable quantitative characterization, due to the relatively large size of most of the encrusting organisms.
Carbonate textures have been named following Dunham (1962) and Embry and Klovan (1971), whereas coral growth fabrics have been described using the terminology by Insalaco (1998). The nomenclature by Woelkerling et al. (1993), originally established for the description of coralline algae growth forms, has been here extended to characterize polygenic crusts made of various encrusting organisms.
Stratigraphic setting and age determination
Stratigraphic setting
The Monte Postale coralgal limestone can be easily identified in the field by its massive to weakly stratified appearance and by the remarkable abundance of coral colonies, calcareous algae, and encrusting organisms, preserved in growth position or as reworked fragments. Along the stratigraphic horizon represented in logs A1, A, B, C, D and D1, the coralgal limestones show a maximum thickness of 13.7 m and are frequently superimposed or interspersed by some dm thick strata of Alveolina grainstone (Figs. 4b, 5a). This massive to thick-stratified organization differs from the thinner, more marked stratification of the sediments in log E, in the upper part of log D1 and in the lower part of log C (Figs. 4b, 5b). There, Alveolina deposits can also be found, but less abundant and associated to well-stratified accumulations of fine-grained, marly wackestones. These stratified deposits are sometimes interrupted by the presence of large slumps.

a Massive coralgal limestone (CL) superimposed by thick strata of Alveolina grainstone (AG), side of log B facing NW. b Stratified succession of fine-grained, marly wackestones (SW), top of log E, outcrop facing SE. c Contact between the stratified, fine-grained wackestones (SW) and the massive coralgal limestone (CL), side of log D facing SE
In Trevisani (2015, Fig. 2), and previously in Munier-Chalmas (1891), the boundary between the massive carbonates and the stratified deposits is considered to be of tectonic origin, related to the presence of a NW/SE oriented, sub-vertical fault across the Monte Postale. In particular, this fault is supposed to separate D and D1 outcrops from the top of the E section (Fig. 4a, b). During our survey, no evidence of this structure has been observed. On the contrary, along the contact between the two lithologies any tectonic breccias have been observed and in two of the best exposed outcrops (southeast side of the D outcrop, Figs. 4a, 5c; upper part of the D1 section, Fig. 4b) the stratified deposits onlap the massive coralgal limestone. In particular, while the sharp contact on the side of the D outcrop (Fig. 5c) could be interpreted as a sequence boundary, the transition between massive and stratified carbonates along the D1 section shows stratigraphic continuity, with no evidence of erosion surfaces. For these reasons, the massive coralgal limestone and the stratified deposits are here considered as heteropic.
Dating the Monte Postale succession
The abundance of the larger foraminifer Alveolina in the Monte Postale succession allowed a reliable biostratigraphic assignment. The limestones below and above the bioconstructions contain among others Alveolina cremae and A. decastroi, markers of the SBZ 11 according to Serra-Kiel et al. (1998).
Calcareous nannofossil analysis of several marly layers sampled along the studied sections (Fig. 4) documents the Biozone CNE 5 of Agnini et al. (2014), corresponding to Zone NP13 of Martini (1971). In particular, the presence of two specimens of Discoaster cf. sublodoensis in a sample 8 m ca. above the massive coralgal limestones, according to Agnini et al. (2014) suggests to assign this portion of Monte Postale succession to the uppermost part of calcareous nannofossil Zone CNE5 (Fig. 6).

Age of the Monte Postale depositional system (dark gray bar) and time interval corresponding to the early Eocene climatic optimum (EECO sensu Luciani et al. 2016; light grey bar) refer to the shallow benthic (SBZ) (Serra-Kiel et al. 1998), calcareous nannofossils (NP) (Martini 1971) and Eocene calcareous nannofossils (CNE) (Agnini et al. 2014) biozones
Hence, based on integration of larger foraminifera and calcareous nannofossil zonations, the age of the Monte Postale depositional system is late Ypresian, spanning the interval between ~50.5 and 48.96 Ma (Vandenberghe et al. 2012; Agnini et al. 2014; Fig. 6).
Facies description
Analyses of carbonate textures and fossil associations of the Monte Postale depositional system indicate the presence of eight different facies (Table 1): coralgal boundstone, polygenic bindstone, coralgal rudstone, Alveolina–Nummulites packstone, Alveolina grainstone, laminated wackestone, non-laminated wackestone, and graded Alveolina grainstone.
Coralgal boundstone
This facies is represented by massive limestones, up to 4 m in thickness, made of scleractinian coral colonies in growth position together with a consistent component of solenoporacean algae. The coral association displays three main types of growth forms: common are massive colonies (1), up to 30 cm in size (Fig. 7a), but most frequent are platy, encrusting morphologies (2), few centimeters thick (Fig. 7b, c), whose superimposition generate a sheetstone growth fabric that constitutes large part of the coral framework. In places, small branching colonies (ramose and phaceloid) (3) are also present, with sticks rarely exceeding 0.8 cm in diameter (Fig. 7d). These different growth-forms can be frequently found at close distance within the same outcrop, showing no evident distribution pattern or zonation. A quantitative characterization of the reef-building biota is given in Fig. 7e. Even considering biases deriving from limitations in outcrop observations, the bioconstruction is dominated by the coral Actinacis (A. cognata, Fig. 7g) and solenoporacean algae of the genus Parachaetetes (Fig. 7f), representing together 40 % of the reef-building skeletal components. The growth form of Actinacis is mostly massive, but fragments of branching colonies are common in thin-sections. Parachaetetes is characterized instead by rounded or thick-branching growth forms. Other relatively common genera are represented mainly by branching colonies (ramose and phaceloid) that include Stylophora, Astrocoenia (Fig. 7h), Astreopora (Fig. 7i), Goniopora, Cladocora, and Pachygyra, although their abundance is probably overestimated, as branching corals are more easily fragmented compared to massive corals. Astreopora and Goniopora also occur as massive (globose) colonies, together with the new genus Paraleptoria (Budd and Bosellini 2015) and Siderofungia. Platy colonies are mainly represented by Actinacis and some agariciid unidentified corals. Subordinate genera are Acropora, Dendracis, Caulastrea, Euphyllia, Favia, Montastraea, and Siderastrea.

Coralgal boundstone facies. a Massive colony of Paraleptoria flexuosissima, polished section. b Platy, encrusting colony of Actinacis cognata, polished section. c Platy, tabular colony of Siderofungia sp., polished section. d Thin sticks of a branching colony of Cladocora sp. (arrows), polished section. e Relative abundance of scleractinian corals and solenoporacean algae genera. f Parachaetetes sp. plants (arrows), thin-section. g Actinacis cognata, thin-section. h Astrocoenia sp., thin-section. i Astreopora tecta, thin-section
Corals and solenoporaceans are usually coated by layered polygenic crusts up to 0.5 cm in thickness and sometimes-adjacent coral colonies are connected by the lateral growth of the encrusting organisms. The biotic composition of these crusts is the same of the polygenic bindstone described in the following paragraph. Both corals and polygenic crusts show extensive evidence of bioperforation. In places, the reef-building association also exhibits traces of abrasion, with the truncation of corals and solenoporaceans and the subsequent recolonization of the eroded surface.
Intra-reef sediment shows a wide range of textures: from a fine-grained mudstone/wackestone to a coarser, often poorly sorted packstone/grainstone. Among bioclasts, most common are fragments of corals and encrusting organisms, dasycladacean green algae, larger foraminifera (mainly Alveolina), miliolids, echinoderms, and molluscs fragments.
Polygenic bindstone
This facies is characterized by massive limestones made of sub-horizontal to irregularly arranged polygenic crusts (Fig. 8a). The superimposition of sub-millimetric to millimetric crusts generated an extremely compact, layered bindstone that can reach 80 cm in thickness. The encrusting biota is represented by a well-diversified assemblage (Fig. 9a–f), a quantitative estimation of which is reported in Fig. 8b. Most abundant are calcareous algae, mainly represented by peyssonneliaceans (Polystrata, Peyssonnelia), by a minor amount of coralline red algae (Sporolithon, Lithoporella, Lithothamnion, some unidentified Mastophoroideae) and solenoporaceans (Parachaetetes). Extremely common are also encrusting foraminifera, mainly represented by Solenomeris and by a lesser quantity of Miniacina, Acervulina, Victoriella, Placopsilina, Carpenteria, Haddonia and Planorbulina. Less common are encrusting coral colonies, 2–3 cm in thickness (Actinacis, Siderofungia and some unidentified agariciid corals), bryozoans, serpulids, vermetids and thin micritic crusts of microbial origin.

Polygenic bindstone facies. a Polished section showing the compact, sub-horizontal superimposition of the polygenic crusts. b Relative abundance of encrusting organisms and bioclastic sediment

Polygenic bindstone facies. a–f Thin-sections showing some of the main encrusting organisms. Peyssonnelia (Pe), Polystrata (Po), Acervulina (Ac), Miniacina (Mi), Placopsilina (Pl), Solenomeris (So), Victoriella (Vi), Lithothamnion (Lith), Lithoporella (Li), Sporolithon (Sp), scleractinian coral (Co)
As in the coralgal boundstone facies, abundant traces of bioerosion, together with sparse signs of abrasion occur. Small cavities among the bindstone are filled by mudstone to packstone deposits with fragments of encrusting organisms, larger foraminifera (mainly Alveolina), echinoderms, molluscs and rare dasycladacean algae.
Coralgal rudstone
Massive to weakly stratified carbonate deposits, up to some meters thick, and rich in poorly sorted skeletal fragments of the same reef-building organisms observed in the coralgal boundstone. Abundant are also dasycladacean algae and fragments of encrusting organisms (Fig. 10a); among the latter, frequent are thin Lithoporella crusts characterized by foliose, hooked and tubular growth forms (sensu Beavington-Penney et al. 2004) (Fig. 10c, d). Fragments of molluscs, larger foraminifera (mainly Alveolina, less frequently orthophragminids and Nummulites), miliolids and echinoderms are also present. Sparse in situ polygenic crusts, similar to those of the polygenic bindstone, have also been observed. These reach 1–2 cm in thickness and extend sub-horizontally among the bioclastic rudstone, binding together the skeletal fragments (Fig. 10b).

Coralgal rudstone facies. a Bioclastic deposit with abundant dasycladacean algae (arrows), thin-section. b Polygenic crusts (arrows) binding the bioclastic fragments, thin-section. c Fragment of Lithoporella crust with hooked morphology, thin-section. d Fragment of Lithoporella crust with hollow morphology, thin-section
Alveolina–Nummulites packstone
Weakly stratified biocalcarenite deposits, cropping out for a maximum thickness of 8 m and organized in decimetric layers. The prevalent texture is a packstone, rich in Alveolina, Nummulites and orthophragminids, all characterized by a moderate to good degree of preservation, together with fragments of corals, encrusting organisms and molluscs. Miliolids and rare dasycladacean algae are also present (Fig. 11a).

a Alveolina–Nummulites packstone facies, bioclastic deposit with Nummulites tests (arrows), thin-section. b Alveolina grainstone facies, bioclastic deposit with abraded skeletal grains, thin-section. c Laminated wackestone facies, alternation of millimetric to sub-millimetric laminae, thin-section. d Non-laminated wackestone facies, concentrations of micrite and small bioclasts suggesting the occurrence of bioturbation, thin-section
Alveolina grainstone
Stratified grainstone to packstone deposits, up to 7 m in thickness and organized in decimetric layers. This facies is characterized by coarse, highly abraded skeletal grains among which abundant are Alveolina, coral fragments and miliolids, associated to orthophragminids, fragments of molluscs and echinoderms. Rare are Nummulites, fragments of encrusting foraminifera and of calcareous algae (Fig. 11b).
Laminated wackestone
Extremely fine grained, sometimes marly, wackestone deposits arranged in thin, centimetric strata, with a maximum overall thickness of 7–8 m. A distinctive feature of these sediments is the occurrence of a planar lamination, more rarely undulated, with the alternation of light and darker laminae, millimetric to sub-millimetric in thickness, and related to variations in the amount of micrite. Some of these strata show the presence of slightly thicker laminae, white in color and undulated. Concerning the bioclastic grains, even if scattered macroscopic fragments of Alveolina, miliolids, corals and calcareous algae have been recognized, their extremely small size usually prevents their precise identification (Fig. 11c). Well-preserved fishes and plants remains can be found inside these strata.
Non-laminated wackestone
This facies is very similar to the laminated wackestone: centimetric layers made of extremely fine grained, sometimes marly wackestone characterized by a maximum overall thickness of 13 m. The main difference is related to the complete lack of lamination, sometimes replaced by irregularly distributed concentrations of micrite and bioclasts that suggest the occurrence of bioturbation (Fig. 11d). No macrofossils have been found within these deposits.
Graded Alveolina grainstone
Alveolina grainstone to packstone deposits are organized in 20–50 cm thick strata, characterized by the presence of an erosional base and normal graded bedding; the coarser bioclasts rarely exceed 1 cm in size. The superimposition of these levels forms deposits up to 12 m in thickness. The skeletal composition of these sediments is the same recognized in the Alveolina grainstone facies.
The Monte Postale reef system
Facies associations
Spatial distribution and stratigraphic setting of the different Monte Postale facies (Figs. 4b, 12) allow to identify three main facies associations and to interpret their depositional paleoenvironments: (1) coralgal rim; (2) fore-reef accumulations; and (3) lagoon deposits.

Spatial distribution of the Monte Postale facies associations. The reconstruction is based on the stratigraphic setting and relative abundance of the different facies within each studied outcrop
Coralgal rim: description
Coralgal boundstone and polygenic bindstone facies represent the bioconstructed portion of the Monte Postale depositional system. Field observations and quantitative analyses place these two facies in the proximal, internal part of the E/W alignment outlined by the arrangement of the studied outcrops (Fig. 12). This distribution identifies the “backbone” of the coralgal rim structure, in which the percentage of the skeletal framework varies from a maximum of 66 % of the overall lithology, in both A and A1 outcrops, to a minimum of 14 % in outcrop C. Among these bioconstructions, bioclastic deposits are also abundant and represented by the coralgal rudstone facies, largely derived from the breakage and reworking of the main reef-building organisms. The amount of this facies varies from 63 % (outcrop D1) to 22 % (outcrop A). Also the Alveolina grainstone deposits belong to the coralgal rim facies association. Stratigraphical setting of Monte Postale indicates these accumulations precede and overlie the massive coral limestone, but Alveolina grainstones can also be found laterally to the bioconstructions (see for example Fig. 4b, logs C and D1), forming bioclastic sand bodies that accumulated beside the skeletal frame.
Coralgal rim: paleoecological setting
A number of components of the coralgal rim biotic assemblage suggest deposition within extremely shallow-water conditions. The remarkable amount of Alveolina, sometimes associated to abundant miliolids, indicates a depositional depth above 40 m (Hottinger 1997). Within the calcareous algae association, the occurrence of peyssonneliaceans (Peyssonnelia and Polystrata), that represent the most important taxa within the polygenic bindstone (Fig. 8b), supports a paleodepth limited to the euphotic zone (Flügel 1979; Fagerstrom 1987). The presence of in situ solenoporacean plants (Parachaetetes) in the coralgal boundstone and polygenic bindstone, and of dasycladacean green algae, particularly concentrated within the coralgal rudstone facies, suggests a bathymetric setting restricted to the shallowest part of the euphotic zone (Flügel 1979, 2004; Fagerstrom 1987; De Castro 1997). Coralline red algae (Corallinales, Sporolithales, Hapalidiales) are generally characterized by a wider bathymetric range, that reaches the lower limit of the photic zone (Littler et al. 1985). However, within the coralgal rim assemblage, specific coralline algae growth forms have been identified, which may be indicative of a more restricted bathymetric setting. Fragments of foliose, hooked and tubular crusts, related to the genus Lithoporella, have been detected, especially concentrated within the coralgal rudstone facies (Fig. 10c, d). These morphologies, indicative of the presence of seagrass beds, are related to the development of coralline algae as epiphytic organisms on macrophytes, adapting their shape to the morphology of leaves, stems and other portions of the plants (Beavington-Penney et al. 2004; Perry and Beavington-Penney 2005; Sola et al. 2013). Preserved fragments of seagrasses have been also found within the laminated deposits of both Monte Postale and Pesciara sites (Wilde et al. 2014). In present-day tropical and temperate marine environments, seagrass meadows are typical of the shallow-water, euphotic zone (e.g., Fornos and Ahr 1997; Hemminga and Duarte 2000; Larkum et al. 2006; Mateu-Vicens et al. 2010).
Despite the evidence of extremely shallow-water setting, growth morphologies of the coralgal rim biota, as well as considerable variations in the sediment textures, indicate the presence of areas characterized by different hydrodynamic conditions. The extremely compact, layered arrangement of the polygenic bindstone (Fig. 8a), where thin, bioclastic accumulations constitute <10 % of the overall lithology (Fig. 8b), suggests that this facies developed within a high hydrodynamic setting, under the continuous effect of waves and tides. This interpretation is supported by the dominance, among coral colonies, of encrusting and massive growth forms, which are both well adapted to high-energy environments (Chappell 1980). However, within the same outcrops, coral colonies with completely different morphologies, indicative of lower hydrodynamic conditions, can also be found. This is the case, for example, of outcrop A1, where close to encrusting coral colonies and polygenic bindstone crusts there are in situ branching corals with extremely thin, delicate sticks (Fig. 7d), the latter usually adapted to very low energy settings. This can be explained by the occurrence, within the coralgal rim structure, of protected areas, sheltered from the action of waves and currents. This hypothesis is also supported by the evidence of mud-supported bioclastic accumulations and by the abundance of dasycladacean algae, whose presence is usually associated to lagoon environmental conditions (Génot 1991; Flügel 2004).
Fore-reef: description
Coralgal rudstone deposits, which are widespread within the coralgal rim, also accumulated northward from the bioconstructed margin (Fig. 12).These sediments are mixed with the Alveolina–Nummulites packstone facies (for example in outcrops A and C, Fig. 12) that gradually increases towards the more distal areas of the depositional system (up to 54 %, in outcrop D, Fig. 12). The association of coralgal rudstone and Alveolina–Nummulites packstone identifies a fore-reef tract, made of accumulations of bioclastic fragments mainly deriving from the dismantling of the coralgal rim by the action of waves and tides. These deposits may have acted as a connection between the coralgal rim and more distal, open-marine environments.
Fore-reef: paleoecological setting
The prevalent packstone texture of the fore-reef deposits suggests a depositional setting with moderate hydrodynamic conditions. The skeletal components of these deposits are quite similar to those of the coralgal rim, except for the common occurrence of Nummulites. Nummulites distribution is generally limited to the euphotic zone, but in comparison to the coralgal rim facies, they indicate a deeper, open-marine depositional setting (Hottinger 1997).
Lagoon deposits: description
The lagoon facies association crops out southward of the coralgal rim (Fig. 12) and is well exposed along the southeastern side of the Monte Postale (Figs. 4a, 5b). These deposits are characterized by the occurrence of three main facies: laminated wackestone, non-laminated wackestone and graded Alveolina grainstone. These lithologies superimpose to each other, forming alternations characterized by sharp contact surfaces (log E, Fig. 4b). Slump deposits sometimes interrupt this succession and in these cases sediments related to the three facies can be found reworked and mixed together.
Lagoon deposits: paleoecological setting
Paleoecological interpretation of this facies association follows the one proposed by Papazzoni and Trevisani (2006) for the sedimentary succession of the Pesciara di Bolca. Texture, composition, and stratigraphic arrangement of the three Monte Postale Lagoon facies show strong similarities with those described for the nearby Pesciara. In particular, the laminated wackestone can be compared with M1 and M2 microfacies of Papazzoni and Trevisani (2006) (respectively, defined as micritic, evenly laminated limestone with black laminae, and micritic, irregularly laminated limestone with white laminae); the non-laminated wackestone is similar to M3 microfacies (micritic, non-laminated limestone), while the graded Alveolina grainstone shows the same features as their F microfacies (biocalcarenite–biocalcirudite with abundant benthic fossils). Based on this comparison, the paleoecological setting of the lagoon facies association can be reconstructed as follows: quiet, low hydrodynamic conditions characterized this protected environment that allowed the deposition of very fine grained, marly wackestones. The proximity to emerged lands is suggested by the presence of terrestrial plants remains, as part of the Fossil-Lagerstätte biota whose occurrence in the Monte Postale is limited to the laminated wackestone facies. The exceptional preservation of this fossil assemblage, as well as the undisturbed, thin lamination, suggest a depositional environment with extremely reduced oxygenation. The presence of anoxic conditions at the bottom of the lagoon has been also indicated by Schwark et al. (2009) on the basis of organic geochemical analyses on the sediments of the Pesciara succession. In addition, the presence of white, undulated laminae, can be interpreted as related to algal mats of cyanobacterial origin, which is as well consistent with the occurrence of anoxic conditions (Papazzoni and Trevisani 2006). To the contrary, the non-laminated wackestone facies, with no evidence of macrofossil preservation, the lack of lamination and the frequent traces of bioturbation, suggests phases characterized by higher oxygenation levels that periodically alternated with the anoxic events. The alternation of these two facies was often interrupted by the deposition of the graded Alveolina grainstone. The erosive base and the normal graded bedding clearly indicate the latter deposits as the product of resedimentation processes, most probably related to storm events. On the basis of the skeletal composition, closely resembling the one of the Alveolina grainstone facies, it is possible to recognize these sediments as transported from the coralgal rim facies association.
Discussion
Depositional model
The coralgal rim facies association represents the core of the Monte Postale depositional system, where the growth of the reef-building organisms (mainly corals, calcareous algae, and encrusting foraminifera) and the accumulation of large benthic foraminifera (mainly Alveolina) actively contributed to the formation of a narrow, elevated structure (Fig. 13) that developed within a maximum paleodepth of a few meters. This margin was approximately oriented in a E/W direction, facing the open sea toward the north, most probably representing the border of a paleo-embayment or intrashelf seaway within the Lessini Shelf. Although the poor preservation of the studied outcrops does not allow reconstructing in detail the spatial distribution of the coralgal rim facies, there is evidence of a coexistence of areas characterized by high and by low to moderate hydrodynamic conditions. This suggests the occurrence of a complex structure, with compact, wave-resistant buildups directly facing the open sea, interconnected by more protected zones. These sheltered environments were inhabited by reef-building organisms with delicate growth forms, associated to bioclastic deposits with variable textures, filling the cavities among the reef-building organisms and contributing to strengthen the skeletal frame. The same accumulations of loose bioclastic sediments may have acted as substrate for the colonization of seagrass patches and dasycladacean green algae. The coralgal rim structure was probably crossed by channels, connecting the open sea with the rear lagoon. This was the environment in which the Alveolina grainstone facies developed, with the high degree of abrasion of its skeletal components most likely related to the continuous action of tides and waves. These accumulations played an important role in strengthening the coralgal rim, contributing to fill the gaps in the bioconstructed portion of the structure.

Reconstruction of the lower Eocene Monte Postale depositional system; the width and depth of the lagoon are not in scale
The role of this margin as an effective barrier, and thus the establishment of a protected lagoon, largely derives from the nature of the Monte Postale biotic association. The large amount of bioclastic sediments removed from the coralgal rim (Alveolina grainstone facies) and accumulated within the Lagoon succession (graded Alveolina grainstone facies) suggests the occurrence of high-energy conditions, most probably related to relatively frequent storms. The width of the Monte Postale sill probably did not exceed 20–30 m across (Figs. 12, 13), even though a larger size could be concealed by the reduced extension of the available outcrops. Such a narrow structure would not have endured extreme hydrodynamic conditions, if not for the contribution of a robust, skeletal framework created by heavily calcified organisms. In addition, the prevalence among the coralgal rim biota of taxa adapted to the euphotic zone (corals, calcareous algae, Alveolina), promoted the preferential growth of the reef assemblage within an extremely shallow-water setting. This contributed to the maintenance of the coralgal rim close to the sea level, limiting the water exchange between the lagoon and the open sea.
The reef system here described represents a single phase in the evolution of the Monte Postale succession, whose reconstruction is mainly based on the coralgal limestone alignment of the logs A1, A, B, C, D and D1 (Fig. 4b). However, considering the whole stratigraphic setting of the area, a more complex situation emerges, with evidence of at least two older reef events (base of the logs E and C; Fig. 4), and of two superimpositions of fine-grained, lagoon wackestones on the top of massive coralgal limestones (base of the log C, log D1; Fig. 4). This setting suggests a complex depositional history, characterized by several changes, with time, in the position of the reef facies associations. These shifts represent different phases in the evolution of the Monte Postale depositional system, most probably as a response to relative sea-level fluctuations. Due to the difficult outcrop situation the following reconstruction of the depositional pattern is partially speculative.
The common presence of fragments of the reef-building association throughout the lagoon sedimentary succession suggests a more or less continuous presence of the coralgal rim. Within this context, three different depositional phases of the Monte Postale margin, associated to relative sea-level fluctuations, are proposed (Fig. 14): (1) during sea-level lowstands, the coralgal rim, even if partially dismantled, could have represented an effective barrier, with the reduction or complete closure of the connections to the sea, the establishment of very low oxygen levels in the lagoon and the prevalent deposition of the laminated wackestone facies (Fig. 14a). This hypothesis agrees with the model proposed by Papazzoni and Trevisani (2006). (2) During rapid transgressions seawater could have risen well above the coralgal rim, increasing the water exchange, improving the lagoon oxygenation and promoting the deposition of the non-laminated wackestone facies (Fig. 14b). (3) During sea-level highstands, the lagoon was progressively isolated, with the gradual re-establishment of anoxic conditions (Fig. 14c). This could have been mainly related to the euphotic adaptation of most of the coralgal rim biotic assemblage, that could have likely promoted the progressive aggradation of the reef structure towards the shallower depths.

Hypothetical evolution of the Monte Postale margin and its control over the lagoon depositional environment, explanation in the text
Early Eocene coral assemblages and reef habitats
The early Paleogene (Paleocene and early Eocene) is usually considered as a transitional period for the evolution of coral reefs, placed between a time of rapid recovery and reorganization of reef ecosystems after the K/Pg crisis and a definitive flourishing phase represented by the Oligocene–Miocene (Perrin 2002; Kiessling and Baron-Szabo 2004). The early Paleogene warming trend culminated in the early Eocene climatic optimum (EECO, ~49–53 Ma; Luciani et al. 2016), the interval in which Cenozoic global temperatures reached their maximum under high atmospheric pCO2 concentrations (e.g., Zachos et al. 2001, 2008; Payros et al. 2015; Slotnick et al. 2015). Recent studies were thus concentrated on the pressure that high temperature and other stressors exerted on coral assemblages and associated bioconstructions during the late Paleocene–early Eocene greenhouse (Scheibner and Speijer 2007, 2008; Zamagni et al. 2012).
Whereas the middle–late Paleocene is characterized by a peak of the global preserved volume of coral reefs, the earliest Eocene shows a marked decline (Kiessling and Baron-Szabo 2004). This coral reef gap has been interpreted as related to rising temperatures and extreme environmental conditions that greatly affected zooxanthellate-coral development and probably favored the expansion of marginal settings where corals evolved with a limited frame-building capacity (Scheibner and Speijer 2007; Zamagni et al. 2012).
The upper Ypresian coralgal bioconstructions of Monte Postale developed near the end of the EECO interval (sensu Slotnick et al. 2015; Luciani et al. 2016). The composition of the reef-building biota reveals strong similarities with the middle Paleocene counterparts: a moderate diversity (up to 16 genera), the dominance of massive Actinacis together with a strong contribution of the solenoporacean alga Parachaetetes and the occurrence of ramose and phaceloid growth forms. This biotic association was able to form small reef structures within shallow-water and high hydrodynamic conditions when global sea temperatures just started to cool but were still high (Zachos et al. 2001). The Monte Postale coralgal reef system highlights that z-coral communities were able to adapt, and colonize shallow-water environments, during the early Eocene global warming phase, thus preserving the potential to flourish and increase their frame-building capacity from the middle–late Eocene, with more favorable climatic conditions.
Conclusions
This study provides a reconstruction of the stratigraphic architecture of the Monte Postale area, combined with new integrated biostratigraphic data and detailed facies analysis. These data allow the first detailed reconstruction of the coralgal-dominated depositional system associated to the formation of one of the most important Eocene Fossil-Lagerstätte in the world. In particular:
-
1.
On the basis of the Alveolina assemblages and calcareous nannofossils associations, the Monte Postale succession is firmly ascribed to the upper Ypresian.
-
2.
Three facies associations have been recognized: a narrow coralgal rim (1), roughly oriented E/W, separating lagoon deposits (2) in the south from fore-reef sediments (3) in the north, the latter representing a transition towards the open-marine environment. In particular, the coralgal rim reveals the occurrence of a complex environment, where scleractinian corals, calcareous algae and encrusting foraminifera, together with Alveolina accumulations, contributed to the formation of a wave-resistant, shallow-water structure interspersed by channels and protected areas.
-
3.
A close correlation has been recognized between the nature of the biotic assemblage and the possibility, by the coralgal rim, to act as an effective barrier. The abundance of heavily calcified organisms was essential in the formation of a robust, wave-resistant threshold, while the euphotic adaptation of most of the reef-related taxa contributed to keep the growth of this structure within an extremely shallow-water setting.
-
4.
Although our data do not allow a precise reconstruction of how the evolution of the coralgal rim influenced the water exchange between the lagoon and the open sea, it is possible to speculate three different depositional phases, associated to relative sea-level fluctuations. Sea-level lowstands or highstands may have limited or interrupted the water connections, supporting the deposition of laminated wackestones and the preservation of vertebrate and terrestrial plants remains. On the contrary, transgressions may have established a more open circulation, with the sedimentation of non-laminated wackestones barren in macrofossils.
-
5.
The Monte Postale succession represents one of the very few examples of lower Eocene reef depositional system. Even if further studies are necessary, the detailed description of the biotic assemblage provides new information on the structure of reef communities during a crucial climatic phase, near the end of the EECO global warming.
References
Agnini C, Fornaciari E, Raffi I, Catanzariti R, Pälike H, Backman J, Rio D (2014) Biozonation and biochronology of Paleogene calcareous nannofossils from low and middle latitudes. Newsl Stratigr 47(2):131–181
Antonelli R, Barbieri G, Dal Piaz GV, Dal Prà A, De Zanche V, Grandesso P, Mietto P, Sedea R, Zanferrari A (1990) Carta geologica del Veneto. Scala 1:250.000—una storia di cinquecento milioni di anni. Regione Veneto, SELCA, Firenze
Barbieri G, Medizza F (1969) Contributo alla conoscenza geologica della regione di Bolca (Monti Lessini). Memorie degli Istituti di Geologia e Mineralogia dell'Università di Padova 27:1–36
Barbieri G, Zampieri D (1992) Deformazioni sinsedimentarie eoceniche con stile a domino e relativo campo di paleostress (Monti Lessini). Atti Ticinensi di Scienze della Terra 35:25–31
Barbieri G, De Vecchi GP, De Zanche V, Di Lallo E, Frizzo P, Mietto P, Sedea R (1980) Note illustrative alla Carta Geologica di Recoaro alla scala 1:20.000. Mem Soc Geol 34:23–52
Barbieri G, De Zanche V, Sedea R (1991) Vulcanismo paleogenico ed evoluzione del semigraben Alpone-Agno (Monti Lessini). Rend Soc Geol Ital 14:5–12
Bassi D, Nebelsick JH (2010) Components, facies and ramps: redefining Upper Oligocene shallow water carbonates using coralline red algae and larger foraminifera (Venetian area, northeast Italy). Palaeogeogr Palaeoclimatol Palaeoecol 295:258–280
Bassi D, Hottinger L, Nebelsick JH (2007) Larger foraminifera of the Late Oligocene of the Venetian area, north-eastern Italy. Palaeontology 50:845–868
Bassi D, Bianchini G, Mietto P, Nebelsick JH (2008) Southern Alps in Italy: Venetian Pre-Alps. In: McCann T (ed) The geology of Central Europe, vol 2. The Geological Society of London, London, pp 1087–1092
Beavington-Penney SJ, Wright VP, Woelkerling WJ (2004) Recognising macrophyte-vegetated environments in the rock record: a new criterion using “hooked” forms of crustose coralline red algae. Sediment Geol 166:1–9
Bellwood DR (1996) The Eocene fishes of Monte Bolca: the earliest coral reef fish assemblage. Coral Reefs 15:11–19
Bellwood DR, Goatley CHR, Brandl SJ, Belwood O (2014) Fifty million years of herbivory on coral reefs: fossils, fish and functional innovations. Proc R Soc Lond B 281:20133046
Bosellini A (1989) Dynamics of Tethyan carbonate platforms. In: Crevello PD, Wilson JL, Sarg JF, Read JF (eds) Controls on carbonate platform and basin platform. S.E.P.M. Spec Publ 44, pp 3–13
Bosellini FR, Trevisani E (1992) Coral facies and cyclicity in the Castelgomberto Limestone (early Oligocene, eastern Lessini Mountains, northern Italy). Riv Ital Paleontol Stratigr 98(3):339–352
Bosellini A, Carraro F, Corsi M, De Vecchi GP, Gatto GO, Malaroda R, Sturani C, Ungaro S, Zanettin B (1967) Note illustrative della carta geologica d’Italia scala 1:100.000, foglio 49 Verona. Servizio Geologico d’Italia, Nuova Tecnica Grafica, Roma
Bosellini A, Masetti D, Sarti M (1981) A Jurassic, “Tongue of Ocean” infilled with oolitic sands: the Belluno Trough, Venetian Alps, Italy. Mar Geol 44:59–95
Budd AF, Bosellini FR (2015) Revision of Oligocene Mediterranean meandroid corals in the scleractinian families Mussidae, Merulinidae, and Lobophylliidae. J Syst Paleontol. doi:10.1080/14772019.2015.1102171
Cannatelli C (2012) Multi-stage metasomatism in the lithosphere beneath the Veneto Volcanic Province (VVP, northern Italy). Mineral Petrol 104(3–4):177–195
Carminati E, Lustrino M, Doglioni C (2012) Geodynamic evolution of the central and western Mediterranean: tectonic vs. igneous petrology constraints. Tectonophysics 579:173–192
Carnevale G, Bannikov AF, Marramà G, Tyler JC, Zorzin R (2014) The Pesciara–Monte Postale Fossil-Lagerstätte: 2. Fishes and other vertebrates. In: Papazzoni CA, Giusberti L, Carnevale G, Roghi G, Bassi D, Zorzin R (eds) The Bolca Fossil-Lagerstätten: a window in the Eocene world. Rend Soc Paleont It 4, pp 37–63
Carraro F, Malaroda R, Piccoli G, Sturani C, Venzo S (1969) Note Illustrative della Carta Geologica D’Italia, Scala 1:100.000. Foglio 48, Peschiera del Garda. Poligrafica & Cartevalori Ercolano, Napoli
Chappell J (1980) Coral morphology, diversity and reef growth. Nature 286:249–252
Dal Degan D, Barbieri S (2005) Rilievo geologico dell’area di Bolca (Monti Lessini orientali). Bollettino Museo Civico di Storia Naturale di Verona 27:3–10
De Castro P (1997) Introduzione allo studio in sezione sottile delle dasicladali fossili—an approach to thin-section study of fossil Dasycladales. Quaderni dell’Accademia Pontiana 22:1–261
De Zanche V, Sorbini L, Spagna V (1977) Geologia del territorio del Comune di Verona. Memorie Museo Civico di Storia Naturale di Verona 1:1–52
Doglioni C, Bosellini A (1987) Eoalpine and mesoalpine tectonics in the Southern Alps. Geol Rundsch 77:734–754
Dunham RJ (1962) Classification of carbonate rocks according to depositional texture. In: Hamm WE (ed) Classification of carbonate rocks, a symposium. AAPG Mem 1, pp 108–121
Embry AF, Klovan JE (1971) A Late Devonian reef tract on Northeastern Banks Island, N.W.T. Can Petr Geol Bull 19:730–781
Fabiani R (1914) La serie stratigrafica del Monte Bolca e dei suoi dintorni. Memorie dell’Istituto Geologico della regia Università di Padova 2:223–235
Fabiani R (1915) Il Paleogene del Veneto. Memorie dell’Istituto Geologico della regia Università di Padova 3:1–336
Fagerstrom JA (1987) The evolution of reef communities. Wiley Interscience, Wiley, New York
Flügel E (1979) Paleoecology and microfacies of Permian, Triassic and Jurassic algal communities on platform and reef carbonates from the Alps. Bull Cent Rech Expl 3:569–587
Flügel E (2004) Microfacies of carbonate rocks. Springer, Berlin
Fornos JJ, Ahr WM (1997) Temperate carbonates on a modern, low-energy, isolated ramp: the Balearic platform, Spain. J Sediment Res 67(2):364–373
Frost SH (1981) Oligocene reef coral biofacies of the Vicentin, northeast Italy. In: Toomey DF (ed) European fossil reef model. S.E.P.M. Spec Publ 30, pp 483–539
Génot P (1991) Cenozoic and recent Dasycladales. In: Riding R (ed) Calcareous algae and stromatolites. Springer, Berlin, pp 131–145
Handy MR, Ustaszewski K, Kissling E (2015) Reconstructing the Alps–Carpathians–Dinarides as a key to understanding switches in subduction polarity, slab gaps and surface motion. Int J Earth Sci 104:1–26
Hemminga MA, Duarte CM (2000) Seagrass ecology. Cambridge University Press, Cambridge
Hottinger L (1997) Shallow benthic foraminiferal assemblages as signals for depth of their deposition and their limitations. B Soc Géol Fr 168:491–505
Insalaco E (1998) The descriptive nomenclature and classification of growth fabrics in fossil scleractinian reefs. Sediment Geol 118:159–186
Kiessling W, Baron-Szabo R (2004) Extinction and recovery patterns of scleractinian corals at the Cretaceous–Tertiary boundary. Palaeogeogr Palaeoclimatol Palaeoecol 214:195–223
Larkum AWD, Orth RJ, Duarte CM (2006) Seagrasses: biology, ecology and conservation. Springer, Dordrecht
Littler MM, Littler DS, Blair SM, Norris JN (1985) Deepest known plant life discovered on an uncharted sea-mount. Science 227:57–59
Luciani V (1989) Stratigrafia sequenziale del Terziario nella Catena del Monte Baldo (provincie di Verona e Trento). Mem Soc Geol 41:263–351
Luciani V, Dickens GR, Backman J, Fornaciari E, Giusberti L, Agnini C, D’Onofrio R (2016) Major perturbations in the global carbon cycle and photosymbiont-bearing planktic foraminifera during the early Eocene. Clim Past 12:981–1007
Macera P, Gasperini D, Ranalli G, Mahatsente R (2008) Slab detachment and mantle plume upwelling in subduction zones: an example from the Italian South-Eastern Alps. J Geodyn 45(1):32–48
Martini E (1971) Standard Tertiary and Quaternary calcareous nannoplankton zonation. In: Farinacci A (ed) Proceedings of the 2nd planktonic conference, Roma, vol 2, pp 739–785
Márton E, Zampieri D, Kázmér M, Dunkl I, Frisch W (2011) New Paleocene–Eocene paleomagnetic results from the foreland of the Southern Alps confirm decoupling of stable Adria from the African plate. Tectonophysics 504:89–99
Mateu-Vicens G, Box A, Deudero S, Rodriguez B (2010) Comparative analysis of epiphytic foraminifera in sediments colonized by seagrass Posidonia oceanica and invasive macroalgae Caulerpa spp. J Foraminifer Res 40:134–147
Mietto P (1992) Monte di Malo. Aspetti Geologici. Comune di Monte di Malo & Centro Studi del Priaboniano “M° Antonio Marchioro”, Monte di Malo, Vicenza
Munier-Chalmas M (1891) Étude du Tithonique, du Crétacé et du Tertiaire du Vicentin. Série Stratigraphique, vol 1. Savy, Paris
Muscio G, Tintori A (2005) I Fossili di Bolca—tesori dalle rocce. Catalogo della mostra di Venezia. Museo di Storia Naturale, Venezia
Papazzoni CA, Trevisani E (2006) Facies analysis, palaeoenvironmental reconstruction, and biostratigraphy of the “Pesciara di Bolca” (Verona, northern Italy): an early Eocene Fossil-Lagerstätte. Palaeogeogr Palaeoclimatol Palaeoecol 242:21–35
Papazzoni CA, Bassi D, Fornaciari E, Giusberti L, Luciani V, Mietto P, Roghi G, Trevisani E (2014) Geological and stratigraphical setting of the Bolca area. In: Papazzoni CA, Giusberti L, Carnevale G, Roghi G, Bassi D, Zorzin R (eds) The Bolca Fossil-Lagerstätten: a window in the Eocene world. Rend Soc Paleont It 4, pp 19–28
Payros A, Ortiz S, Millan I, Apellaniz E (2015) Early Eocene climatic optimum: environmental impact on the North Iberian continental margin. GSA Bull 127:1632–1644
Perrin C (2002) Tertiary: the emergence of modern reef ecosystems. In: Kiessling W, Flügel E, Golonka J (eds) Phanerozoic reef patterns. SEPM Spec Publ 72, pp 587–621
Perry CT, Beavington-Penney SJ (2005) Epiphytic calcium carbonate production and facies development within sub-tropical seagrass beds, Inhaca Island, Mozambique. Sediment Geol 174:161–176
Piccoli G (1966) Subaqueous and subaerial basic volcanic eruptions in the Paleogene of the Lessinian Alps. Boll Soc Geol Ital 84:141–157
Roghi G, Dominici S, Giusberti L, Cerato M, Zorzin R (2014) Historical outline. In: Papazzoni CA, Giusberti L, Carnevale G, Roghi G, Bassi D, Zorzin R (eds) The Bolca Fossil-Lagerstätten: a window in the Eocene world. Rend Soc Paleont It 4, pp 5–17
Sarti M (1980) Frane sottomarine e debris flow in una successione carbonatico torbiditica eocenica (Val d’Avesa, Verona). Annali dell’Università di Ferrara, Sez. IX Scienze Geologiche e Paleontologiche 7:65–89
Scheibner C, Speijer RP (2007) Decline of coral reefs during late Paleocene to early Eocene global warming. eEarth Discuss 2:133–150
Scheibner C, Speijer RP (2008) Late Paleocene–early Eocene Tethyan carbonate platform evolution. A response to long- and short-term paleoclimatic change. Earth Sci Rev 90:71–102
Schmid SM, Fügenschuh B, Kissling E, Schuster R (2004) Tectonic map and overall architecture of the Alpine orogen. Eclogae Geol Helv 97:93–117
Schwark L, Ferretti A, Papazzoni CA, Trevisani E (2009) Organic geochemistry and paleoenvironment of the early Eocene “Pesciara di Bolca” Konservat-Lagerstätte, Italy. Palaeogeogr Palaeoclimatol Palaeoecol 273:272–285
Serra-Kiel J, Hottinger L, Caus E, Drobne K, Ferràndez C, Jauhri AK, Less G, Pavlovec R, Pignatti J, Samsó JM, Schaub H, Sirel E, Strougo A, Tambareau Y, Tosquella J, Zakrevskaya E (1998) Larger foraminiferal biostratigraphy of the Tethyan Paleocene and Eocene. Bull Soc Géol Fr 169:281–299
Slotnick BS, Dickens GR, Hollis CJ, Crampton JS, Strong PS, Phillips A (2015) The onset of the early Eocene climatic optimum at branch stream, Clarence River valley, New Zealand. N Z Geol Geophys. doi:10.1080/00288306.2015.1063514
Sola F, Braga JC, Aguirre J (2013) Hooked and tubular coralline algae indicate seagrass beds associated to Mediterranean Messinian reefs (Poniente Basin, Almería, SE Spain). Palaeogeogr Palaeoclimatol Palaeoecol 374:218–229
Sorbini L (1972) I fossili di Bolca. Edizioni Corev, Verona
Sorbini L (1999) I giacimenti di Bolca. In: Pinna G (ed) Alle radici della storia naturale d’Europa. Seicento milioni di anni attraverso i grandi giacimenti paleontologici. Jaca Book, Milano, pp 172–176
Trevisani E (1997) Stratigrafia sequenziale e paleogeografia del margine orientale dei Lessini Shelf durante l’Eocene superiore (Prealpi Venete, provincie di Vicenza e Treviso). Studi Trentini di Scienze Naturali, Acta Geologica 71:145–168
Trevisani E (2015) Upper Cretaceous–Lower Eocene succession of the Monte Postale and its relationship with the “Pesciara di Bolca” (Lessini Mountains, northern Italy): deposition of a fossil-fish lagerstätte. Facies. doi:10.1007/s10347-015-0431-y
Ungaro S (2001) Le biofacies paleoceniche ed eoceniche dei Monti Lessini (Veneto, Italia). Annali Università di Ferrara, Sez. Scienze della Terra 9:1–40
Vandenberghe N, Hilgen FJ, Speijer RP (2012) The Paleogene period. In: Gradstein FM, Ogg JG, Smith AG, Ogg GM (eds) The geologic time scale 2012. Elsevier, Amsterdam, pp 855–921
Wilde V, Roghi G, Martinetto E (2014) The Pesciara–Monte Postale Fossil-Lagerstätte: 3. Flora. In: Papazzoni CA, Giusberti L, Carnevale G, Roghi G, Bassi D, Zorzin R (eds) The Bolca Fossil-Lagerstätten: a window in the Eocene world. Rend Soc Paleont It 4, pp 65–71
Winterer EL, Bosellini A (1981) Subsidence and sedimentation on Jurassic passive continental margin, Southern Alps, Italy. AAPG Bull 65:394–421
Woelkerling WJ, Irvine LM, Harvey A (1993) Growth-forms in non-geniculate coralline red algae (Corallinales, Rhodophyta). Aust Syst Bot 6:277–293
Zachos JC, Pagani M, Sloan L, Thomas E, Billups K (2001) Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 292:686–693
Zachos JC, Dickens GR, Zeebe RE (2008) An early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics. Nature 451:279–283
Zamagni J, Mutti M, Košir A (2012) The evolution of mid Paleocene–early Eocene coral communities: how to survive during rapid global warming. Palaeogeogr Palaeoclimatol Palaeoecol 317–318:48–65
Zampieri D (1995) Tertiary extension in the southern Trento Platform, Southern Alps, Italy. Tectonics 14(3):645–657
Acknowledgments
The authors wish to thank the editor, Wolfgang Kiessling, for his valuable comments. The manuscript has benefited greatly from critical comments and suggestions by two anonymous reviewers. We thank S. Paladini, B. Fornaciari, A. Olivieri, M. Plessi and L. Barozzi for the assistance during fieldwork and microfacies analyses. We also thank the Cerato family for their collaboration, their useful suggestions, and great kindness. This study was funded by the Italian Ministry of Education and Research (MIUR) funds PRIN 2010–2011 (prot. 2010X3PP8J_002 and prot. 2010X3PP8J_003) project “Past ExcessCO2 worlds: biota responses to extreme warmth and ocean acidification”.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Vescogni, A., Bosellini, F.R., Papazzoni, C.A. et al. Coralgal buildups associated with the Bolca Fossil-Lagerstätten: new evidence from the Ypresian of Monte Postale (NE Italy). Facies 62, 21 (2016). https://doi.org/10.1007/s10347-016-0472-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10347-016-0472-x