
Abstract
We investigated the movement pattern of GPS-collared red deer (Cervus elaphus) during drive hunts with loose dogs. In 46 flights of hinds (N = 9), the median flight distance was 2.5 km (range 0.4–15.0). In 28 % of the flights, the hind left its home range. Average time before returning to home range was 23 h (range 2–88). Hinds in a less forested site left their home ranges more often, fled longer distances, moved at higher speed, and returned later than hinds in a homogenous forest. Speed of movement increased with number of hunts in the less forested site. In eight flights of stags (N = 4), the median flight distance was 5.1 km (range 2.2–13.3). The start of hunting season seemed to trigger stags’ departure to their wintering areas. Knowledge of reactions to disturbance can aid game management to choose hunting methods and dogs that cause less disturbance but may also be used to deter deer from areas where they damage crops and forest plantations.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Human disturbance triggers natural anti-predatory behaviors among prey (Frid and Dill 2002; Sibbald et al. 2011) and may therefore impose short-term as well as long-term effects on behavior and spatial use (Frid and Dill 2002; Apollonio et al. 2005; Pan et al. 2011). Directed disturbance, such as hunting, is believed to impose an especially strong impact. Hunting has been shown to result in increased vigilance, altered movement patterns, shifts in timing of migration, increased home range sizes, temporary abandonment of home range, change in habitat use, and decreased diurnal activity in open areas among ungulates (Swenson 1982; Jeppesen 1987; Root et al. 1988; Vercauteren and Hyngstrom 1998; Kilpatrick and Lima 1999; Millspaugh et al. 2000; Conner et al. 2001; Viera et al. 2003; Benhaiem et al. 2008; Sunde et al. 2009; Scillitani et al. 2009). The behavioral response of ungulates to hunting can depend on habitat characteristics (Vercauteren and Hyngstrom 1998; Brown et al. 1999; Millspaugh et al. 2000; Conner et al. 2001), hunting pressure (Kilpatrick and Lima 1999; Johnson et al. 2004; Scillitani et al. 2009), as well as type of hunting method (Root et al. 1988; Williams et al. 2008; Thurfjell et al. 2013). Hunting methods that involve some kind of chase or forcing animals to abandon preferred habitat seem more likely to induce severe disturbance than hunting methods like sit-and-wait or stalking. The response to human disturbance can depend on age, sex, and social interactions (McCorquodale 2003; Apollonio et al. 2005; Williams et al. 2008), and it is thus possible that hunting will result in different behavioral responses among age classes, males and females, and various social groupings. The hunting activities can also disturb other species than the targeted species (Grignolio et al. 2011), with potentially negative effects on disturbance-sensitive species or even threatened species.
For the purposes of wildlife management and conservation, a good knowledge of disturbance effects of hunting should, thus, be of great importance. Hunters and wildlife managers should want to use hunting methods that provide good opportunities for safe and selective shots. They may also want to disregard methods causing the target species, or other species, to abandon the hunting unit or the wildlife management area. From a conservation perspective, it is important to know whether and how hunting activities interfere with preservation goals of sensitive or threatened species. Disturbance is suggested to increase damage to forestry when the deer need to seek security cover in dense plantations, where trees and plants may be heavily browsed (Gill 1992; Borkowski and Ukalski 2012). However, it is also suggested that disturbance can be used to deter animals from resources sensitive to damage (Nolte 1999).
Red deer (Cervus elaphus) seem prone to react with flight to various kinds of disturbance such as hunting, military activities, and outdoor life (Jeppesen 1987; Bateson and Bradshaw 1997; Phillips and Alldredge 2000; Sunde et al. 2009; Sibbald et al. 2011). In handbooks on red deer management (e.g., von Raesfeld and Vorreyer 1974; Wagenknecht 1986), it is emphasized that red deer should be left undisturbed in daytime resting places and in grazing zones, and authors recommend using hunting methods that minimize disturbance. The predator most likely to have had the largest impact on red deer, and shaped its anti-predatory behavior, is the wolf (Canis lupus), which usually attacks deer after a short hunt (Nelson and Mech 1991; Okarma 1995; 1997; Wikenros et al. 2009). Prolonged flights, e.g., during traditional hunts with packs of dogs in Great Britain, therefore are unnatural for red deer. Long flights also have physiological effects, signs of which begin to appear already early in the flight or after short flights (Bateson and Bradshaw 1997). This might indicate that red deer are poorly adapted to drive hunts with loose dogs.
Red deer can also react to hunting by leaving disturbed areas (Jeppesen 1987; Conner et al. 2001; Vieira et al. 2003; Sunde et al. 2009). Habitat structure is suggested as perhaps the most important factor for prey to assess predation risk and the possibility to escape an attack (Brown et al. 1999; Verdolin 2006). The structure of the landscape and type of habitat can, thus, have an impact on the consequences of disturbance, and a stronger effect can be expected in more open and fragmented habitats and landscapes (Jeppesen 1987; Vercauteren and Hygnstrom 1998; Millspaugh et al. 2000; Conner et al. 2001; Jayakody et al. 2008; Sunde et al. 2009). For wildlife managers, hunters, and landowners aiming at a careful red deer management in balance with other management goals, there may be good reasons to consider possible disturbance effects from hunting and to try to evaluate the consequences of disturbance in order to adjust hunting methods and minimize unwanted effects.
In this study, we investigated the behavior of GPS-collared red deer during drive hunts in a forest-dominated landscape in eastern Sweden. We studied movement patterns with particular interest in flight distances, frequency of flights, and elapsed time before returning to the home range as well as speed of movement after a disturbance in relation to habitat composition. Our aim is to increase knowledge on how red deer react to hunting disturbance, knowledge that we think will be useful for deer management as well as for the development of strategies to counteract damage to crops and forest plantations.
Study area
The research area in Kolmården (58°78′ N, 16°43′ E for center of area) is situated in eastern Sweden, northeast of Norrköping city, and approximately 130 km southwest of Stockholm. Kolmården has no exact borders, but is an old name of an area of deep inaccessible forests and rocky terrain, stretching over the border between the counties of Östergötland and Södermanland. The area is a largely homogenous forest landscape dominated by managed forests of spruce (Picea abies) and Scots pine (Pinus sylvestris). In the eastern parts of the area, agricultural areas stretch into the forest landscape creating a mix in the border zone. Red deer, roe deer (Capreolus capreolus), moose (Alces alces), and wild boar (Sus scrofa) are common in the area, whereas fallow deer (Dama dama) occur in low numbers. Occurrence of lynx (Lynx lynx) is sparse but permanent, and wolves occur on rare occasions.
The research area covers approximately 32,000 ha. An aerial count in February 2006 estimated the postharvest population density of red deer to 32 per 1,000 ha. The general hunting season starts on the second Monday in October and lasts until 31 January. However, during 16th August to the second Monday in October, culling hinds and calves is allowed, but only by using sit-and-wait or stalking hunting methods. Red deer hunting is organized in management areas where the hunters make three-year culling plans that must be approved by the county administrative board. In the research area, the harvests of the hunting seasons starting in 2006, 2007, and 2008 were 373, 440, and 397 deer, respectively.
Within the research area, red deer hinds were marked on the following three different estates: Stavsjö, Virå, and Valinge. Stavsjö and Virå are neighbors in the central part of the Kolmården area, and the marked females in this area had home ranges largely overlapping the mutual border. Therefore, Stavsjö and Virå are treated as one study site (hereafter referred to as Stavsjö–Virå) and Valinge as another. The two study sites differ markedly in the character of the landscape. Stavsjö–Virå represents a homogenous forest landscape with almost no agricultural areas, whereas Valinge is more fragmented with a substantial amount of agricultural fields (Allen 2012). Stavsjö–Virå consists of 84 % forest, 8 % mire, 6 % bedrock, 1 % agricultural land and 1 % buildings etc. Valinge is situated in the east, where the landscape turns to a mix between forest and agricultural land. Valinge consists of 64 % forest, 2 % mire, 8 % bedrock, 22 % agricultural land, and 5 % buildings etc. (habitat composition available from the forestry management plans of the estates).
Methods
Adult red deer hinds (randomly chosen) were tranquilized (Dan-inject rifle, immobilized with Fentanyl and Xylazine, and using Naltrexone as antidote) at feeding stations in February–March. The number of marked hinds was nine: six at Stavsjö–Virå and three at Valinge. To minimize risk for the neck outgrowing the collar, only full-grown males with an estimated age of at least 5–6 years (judged from antler size and body morphology (Hetschold and Vorreyer 1968; Krebs 1969; Drechsler 1988)) were chosen for marking. Four males were available for the present study. Three males were marked at feeding stations in February–March 2007 on two wintering sites 10 and 12 km from the rut area at Stavsjö–Virå (arrived at Stavsjö–Virå before the rut and spent the rut there), and one male was marked at Valinge during the rut 2007. The deer were fitted with a GPS collar (Vectronic Aerospace, Germany) and plastic ear tags.
The study covered the hunting seasons 2006/07, 2007/08, and 2008/09. The number of marked hinds during the three seasons was nine, eight, and five, respectively. The number of marked stags was four in 2007/08 and three in 2008/09. On dates of drive hunts inside the home ranges of marked deer, the GPS collars were programmed to start 1–2 h before estimated hunting start and to register one location per 10 or 15 min during 24 h (on non-hunting days, the collars registered 5–6 locations during 24 h). Hunters were not informed of the location of the marked deer before the hunt. The hunting method used in both study sites was drive hunt with loose dogs. During these hunts, shooters are placed out around the area that is to be hunted, whereupon dog keepers from various points enter the area and release their dogs. The size of a hunted area was generally 50–150 ha, and 3–5 areas were hunted during one hunting day. The idea is that the dogs should pursue the deer only for distances long enough to make the deer leave the hunted area. The dog-keepers therefore try to stay in contact with the dogs and if necessary directing them with commands in order to ensure that the area is systematically searched and that the dogs remain inside the hunted area. The hunting in one area lasts for about 45–90 min before it is called off. At Valinge, the number of dogs was 7–8 per drive and the number of shooters 15. The corresponding figures for Stavsjö–Virå were 2–6 and 10–15. Dog breeds commonly used were Deutscher Wachtelhund and cross-breed Deutscher Wachtelhund/Norwegian Elkhound. In part, the same dogs and dog keepers were active in both study sites. Hunters were asked to avoid shooting marked animals, and there were not any marked deer shot during the hunts.
Return times, frequency of flight behavior, and flight distances
One marked deer with functioning collar with a drive hunt inside its home range was termed as a “deer-specific hunting event” (Sunde et al. 2009) when analyzing flight distances and return times (only for hinds). The average used area per day during daytime for the hinds in the study are 0.3 km2 (Allen 2012). To exclude possible movements unrelated to the hunting activities, a clear detectable flight was chosen and defined as when a deer made a sudden move of at least 300 m during or after the hunt. Observed flight distances and return times for fleeing deer were measured in ArcView version 3.2 (Environment Systems Research Institute (ESRI), Redlands, California, USA) by adding distances between consecutive locations per individual deer. Return time was defined as the time between leaving and returning to the home range (annual home range calculated using Minimum Convex Polygon (Mohr 1947).
We used the module generalized linear mixed model and a binary logistic regression to analyze factors influencing if red deer hinds (stags were excluded due to the small sample size) fled or not during the drive hunts. Site (Valinge and Stavsjö–Virå) was entered as a fixed factor to test the effect of different habitat in the two study sites and hunt per season (2–13) and hunting season (1–3) as continuous factors (Table 1). To account for correlations between repeated observations of the same individuals, we used hind ID as a random effect.
We used a linear mixed model (LMM) to test the effect of site (Valinge and Stavsjö–Virå) on flight distances. We log-transformed the dependent variable (flight distance). Site was set as a fixed factor and hind ID as a random factor to control for a possible effect of individual, but we did not account for repeated hunts during the same hunting season or between hunting seasons in this analysis.
Speed of movement
We used a LMM to analyze factors influencing movement by red deer hinds during drive hunts. We calculated speed of movement (meter/minute) between consecutive locations by measuring the distance in ArcGIS version 9.3.1 (ESRI, Redlands, California, USA) and calculate time interval. We only used locations that were a maximum of 20 min apart in order to not include time when the GPS failed to register positions. We log-transformed the dependent variable (speed of movement). Site (Valinge and Stavsjö–Virå) was entered as a fixed factor, and hunt per season (2–13) and hunting season (1–3) as continuous factors, as well as hind-ID as a random effect (to account for correlations between repeated observations of the same individual) and two interaction effects (“site × hunt per season” and “site × hunting season”). Akaike’s information criterion (AICc) was used for ranking quality of models. We considered the model with the lowest AICc score as the best model (Burnham and Anderson 2002). All statistical analyses were conducted using SPSS Statistics version 21.0 (IBM SPSS Inc. Chicago, IL, USA).
Results
Red deer hinds
Flight distances and return times of red deer hinds were studied during 46 drive hunts: 33 at Stavsjö–Virå and 13 at Valinge. Of a total of 156 deer-specific hunting events (122 at Stavsjö–Virå and 34 at Valinge), there was a clear detectable flight at 46 (29 %) deer-specific hunting events, 26 (76 % of 34 events) at Valinge, and 20 (16 % of 122 events) at Stavsjö-Virå. In 43 (93 %) of the flights, the flight was made during the hunt. In two flights, there was a first stage of flight during hunt and a second continued flight in the evening after the hunt. One flight was entirely made during the night after the hunt. All hinds fled at least two times during the study period. Red deer hinds fled more often in Valinge compared to Stavsjö–Virå (Table 2, F1, 152 = 37.650, p < 0.001). The increasing number of hunts per season was a significant predictor of flight behavior (Table 2, F1, 152 = 9.372, p = 0.003), and there was also a tendency for a significant difference between the three hunting seasons (Table 2, F1, 152 = 3.539, p = 0.062). The distance between start of flight and stop ranged between 0.4 and 15.0 km (N = 46, median 2.5 km). With a median flight distance of 3.4 km (range 0.9–15.0 km), hinds at Valinge generally fled longer distances than hinds at Stavsjö–Virå (F1,5.25 = 7.016, p = 0.043), where the median flight distance was 2.0 km (range 0.4–7.1 km). In 28 % (N = 13, 11 at Valinge, and 2 at Stavsjö–Virå) of the flights, the hind left its home range. The average time before returning to the home range was 23 h (range 2–88, SD = 25 (The 11 flights at Valinge had an average return time of 26 h, and at Stavsjö–Virå, the return times were 2 and 11 h.)).
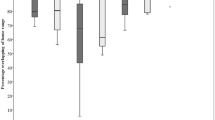
We used a total of 14,215 estimations of speed of movement of red deer hinds during drive hunts. Among the 12 models used to model variation in speed of movement, the model with the lowest AICc value included hind ID, site, hunts per season, hunting season, and the interactions “site × hunt per season” and “site × hunting season” (Table 3). Red deer hinds moved faster in Valinge compared to Stavsjö–Virå (Fig. 1, Table 4). In Valinge, the speed of movement increased with the number of hunts during the hunting season, but this effect was not evident in Stavsjö–Virå (Table 4). Speed of movement decreased during the three hunting season in Valinge but remained similar in Stavsjö–Virå (Table 4).

Speed of movement (N = 14,215) in relation to hunt per season during three consecutive hunting seasons (1–3, corresponding to years 2006/2007–2008/2009) of red deer hinds during drive hunts with loose dogs in eastern Sweden. The study site Stavsjö–Virå is an almost homogenous forest landscape, whereas Valinge is a mixed forest–agricultural landscape
Red deer stags
For the red deer stags, there were 8 flights registered during 16 deer-specific hunting events. It is unknown whether the non-fleeing stags were unaffected by dogs and hunters, or if they managed to face and withstand the dogs. Median flight distance was 5.1 km (range 1.2–13.3 km). Three flights were made entirely during the hunt, three during the night after the hunt, and two flights were made in two parts with a first part during the hunt and a second during night. During three of the flights (three different individuals) in the beginning of the hunting season, the stag left its rutting place and migrated to its winter–summer area, not to return to the rutting area until the next year’s rut. The stags that performed a seasonal migration between the summer or winter areas and the rutting area had all left the rutting area by the end of the first week of hunting, both in 2007 (three of four stags) and in 2008 (two of three stags, same migrating individuals as in 2007).
Discussion
The median flight distance of 2.5 km by hinds may appear close to the 3.5 km found by Jeppesen (1987) and the 4.0 km found by Sunde et al. (2009), both studies conducted in Denmark. That the hunting activities had an effect on the movements of the hinds was clear. Undisturbed, the same hinds rarely moved in excess of 4 km2, being active mainly during night, with daytime resting areas averaging 0.25 (Stavsjö–Virå) and 0.37 km2 (Valinge) (Allen 2012). The maximum distance of 15 km in the present study is also near the 17 km maximum recorded by Sunde et al. (2009). However, whereas Jeppesen (1987) seem to have measured flight distance from position before disturbance, the definition of a flight differed between Sunde et al. (2009) and the present study. Sunde et al. (2009) only acknowledged flights reaching outside the home ranges and then measured flight distance from nearest home range border, indicating that the actual flight distances were longer. The average flight distances recorded in Denmark and Sweden are, however, low compared to the average of 19 km that Bateson and Bradshaw (1997) measured in England, but then the use of hound packs that chase the deer for several hours is likely to impose a more persistent and severe disturbance than the Swedish methods do.
The hinds at Valinge fled more often, longer distances, and at higher speed than the hinds at Stavsjö–Virå. The number of dogs in the drives was higher at Valinge, which possibly may have affected the flight behavior. The most likely explanation, however, should be the structure of the landscape. Stavsjö–Virå represents an almost homogenous forest landscape, whereas Valinge is a mixed forest–agricultural landscape. It may be that deer are more easily disturbed in fragmented and more open landscapes (Jeppesen 1987; Millspaugh et al. 2000; Conner et al. 2001; Jayakody et al. 2008; Sunde et al. 2009). It is probably more difficult to avoid detection and to remain in smaller forest patches, and if a flight cannot be avoided, the distance to the next forest patch offering shelter is probably longer. The difference in speed of movement between the two areas could also reflect that flights conducted in open and most likely high risk habitats should be made at high speed, whereas flights in concealed habitats can be performed more cautiously. Moreover, it should also be easier to run fast in the open compared to in the forest with hindering vegetation and, as was the case in the present study areas, rocky terrain and boulders.
Jeppesen (1987) does not report number of hunts, but states that hinds left their home ranges in 10 out of 18 recorded flights in connection with hunts. Sunde et al. (2009) observed that hinds in their study left their home range in 53 % of the deer-specific hunting events. This could be compared to the hinds in the present study that only left their home ranges in 8 % of all marked deer hunts (13 out of 172) and in 28 % of the flights. However, the detailed movements of the dogs and the dog keepers following the dogs were unknown to us and not used in the analysis. The proportion-marked deer that actually came in contact with hunters or dogs was, thus, also unknown, whereby the proportion of fleeing deer and disturbed deer may be substantially higher. It is therefore difficult to decide whether a hind not making a move during or after the hunt was in close contact with dogs or hunters, but managed to stand still and avoid detection, or if the hind simply was not disturbed at all. Even so, the lower ratio of leaving home range in the present study compared to the Danish studies could be attributed to the more or less homogenous forest landscape. In Jeppesen (1987), the study area consisted of 63 % open land. In the study of Sunde et al. (2009), deer were marked in a 1,400 ha forest plantation surrounded by open, mostly agricultural, land, indicating that if the deer left the plantation, they had to move relatively long distances to reach security cover. Another difference between the present study and the Danish studies is that the Danish hinds commonly postponed their flight until dark, whereas the hinds in our research area generally left already during the hunt. Also, this could be due to the different types of landscapes. In the forested landscape, it may be possible to conduct a flight in closed habitats, but in the fragmented landscape, the fleeing deer probably need to cross open areas—something that may be highly risky during daytime. Although the sample size is small, it was interesting to see an indication of higher tendency for males to postpone the flight until after the hunt, perhaps indicating behavioral differences between the sexes.
The average return time of 23 h in the present study can be compared to Sunde et al. (2009) who recorded an average return time of nearly 5 days. It is possible that the longer return time is an effect of the more open and fragmented landscape in western Denmark. A majority of the hinds returned already within 24 h, often during the night after the hunt. This could perhaps imply that the disturbance effect of a drive hunt is not that severe. However, a factor that needs to be considered is how often red deer experience drive hunts. The estates in this study all tried to minimize the number of drive hunts per season, aiming at causing as little disturbance as possible, yet reaching their harvest goals. It is, however, not uncommon in Sweden to hold drive hunts every week (typically during weekends) throughout the hunting season.
With increasing number of hunts per hunting season, the hinds at Valinge increased their speed of movement (Fig. 1). They thereby seemed increasingly disturbed with number of hunts, even though they also seemed to get used to the disturbance with time (Fig. 1). This suggests that the number of drive hunts per season should be considered. However, this might also indicate that frequent disturbances could offer a method to decrease damage to forest plantations and crops. It would, thus, be interesting to investigate whether more frequent drive hunts cause red deer to abandon areas for longer periods, or even permanently. If deer react to repeated disturbance by leaving areas, this can be used to deter deer from forest plants or crops sensitive to damage as part of a push–pull strategy (Pyke et al. 1987; Miller and Cowles 1990; Nolte 1999). The concept of push–pull strategies is to repel (push) pest species from unsuitable resources and attract (pull) them to areas or resources where they can be eliminated, or where their presence is harmless (Cook et al. 2007).
Stags left the rutting area during or in connection to hunts during the first hunting week not to return to their rutting areas until the next year’s rut. It may, therefore, be that the high hunting activity during this first week more or less functions as a signal to the stags that it is time to depart and head for the winter area. That the timing of migration can be affected by hunting activities has been shown for red deer in North America (Conner et al. 2001; Vieira et al. 2003). One interesting question is whether this has been a successful survival strategy for stags compared to staying, i.e., that the risk of being culled is higher in the rutting area than in the winter area, and whether hunting can reinforce sexual segregation in red deer and affect timing of seasonal migration of the males (Jarnemo 2008).
Hunters may reconsider the strategy of starting the hunting season with drive hunts and instead use less disturbing hunting methods such as sit-and-wait or stalking during the first weeks of the season. The advantages of using these methods are that the stags might remain longer in the rutting areas, and that a selective harvest is easier to conduct. Management areas aiming at a sustainable harvest of a deer population with an even sex ratio and a high average age of males often express worries about the deer (especially males) moving outside the management unit and become a subject to high harvest rates and being shot before reaching fully mature age. With these objectives, management areas should carefully consider alternative hunting methods to drive hunts and try to decrease the number of drive hunts.
Red deer can cause severe damage to forest plantations and crops (Gill 1992; Verheyden et al. 2006; Reimoser and Putman 2011; Månsson and Jarnemo 2012), and the suggested counteraction generally is to decrease population density (Putman et al. 2011; Reimoser and Putman 2011). However, game management is increasingly important for rural economy in Europe (Apollonio et al. 2010; Mysterud 2010) as well as in Sweden (Mattsson et al. 2008; Liberg et al. 2010; Hillve 2012), and red deer is a species of high status among hunters (Milner et al. 2006; Mattson et al. 2008; Mysterud 2010). Game management may want to sustain high densities of deer, and use harvest techniques securing a long-term maintenance of deer and not risking deer abandoning the grounds. There are also cases where conservation efforts are made for the preservation of threatened subspecies of red deer (Hartl et al. 2003; Lovari et al. 2008; Höglund et al. 2013). Conflicting goals, e.g., forestry and agriculture versus game management or conservation, thus create a need for solutions enabling the combination of forestry and crop production with economic game management or preservation of threatened species/subspecies. Not causing too much disturbance in areas where the deer are accepted, combined with disturbance actions in damage sensitive areas, may offer one possible solution to this challenge. This, however, requires detailed knowledge of how deer react to various types of disturbances.
References
Allen A (2012) How does the landscape structure shape the movement and feeding patterns of red deer (Cervus elaphus) in Sweden? Master thesis, Imperial College London
Apollonio M, Ciuti S, Luccarini S (2005) Long-term influence of human presence on spatial sexual segregation in fallow deer. J Mamm 86:937–946
Apollonio M, Andersen R, Putman R (2010) European ungulates and their management in the 21st century. Cambridge University Press, Cambridge, UK
Bateson P, Bradshaw EL (1997) Physiological effects of hunting red deer (Cervus elaphus). Proc R Soc Lond B 264:1707–1714
Benhaiem S, Delon M, Lourtet B, Cargnelutti B, Aulagnier S, Hewison M, Morellet N, Verheyden H (2008) Hunting increases vigilance levels in roe deer and modifies feeding site selection. Anim Behav. doi:10.1016/j.anbehav.2008.03.012
Borkowski J, Ukalski K (2012) Bark stripping by red deer in a post-disturbance area: the importance of security cover. For Ecol Manag 263:17–23
Brown JS, Laundre JW, Gurung M (1999) The ecology of fear: optimal foraging, game theory, and trophic interactions. J Mamm 80:385–399
Burnham KP, Anderson DR (2002) Model selection and inference: a practical information–theoretic approach. Springer, New York
Conner M, White GC, Freddy DJ (2001) Elk movement in response to early season hunting in Northwest Colorado. J Wildl Manag 65:926–940
Cook SM, Khan ZR, Pickett JA (2007) The use of push–pull strategies in integrated pest management. Ann Rev Entom 52:375–400
Drechsler H (1988) Altersentwicklung und Altersansprache beim Rotwild. Gestalten, Gesichter, Gehabe, Geweihe, Gebisse. Hamburg, Paul Parey
Frid A, Dill L (2002) Human-caused disturbance stimuli as a form of predation risk. Ecol & Soc 6: article 11: http://www.ecologyandsociety.org/vol6/iss1/art11/print.pdf
Gill RMA (1992) A review of damage by mammals in north temperate forests: 1. Deer Forestry 65:145–169
Grignolio S, Merli E, Bongi B, Ciuti S, Apollonio M (2011) Effects of hunting with hounds on a nontarget species living on the edge of a protected area. Biol Cons 144:641–649
Hartl GB, Zachos F, Nadlinger K (2003) Genetic diversity in European red deer (Cervus elaphus L.): anthropogenic influences on natural populations. C. R. Biol 326:37–42
Hetschold W, Vorreyer F (1968) Das Ansprechen des Rotwildes. Hamburg, Paul Parey
Hillve P (2012) Den svenska jakten – ett näringsperspektiv. Rapport. Svenska Yrkesjägareföreningen, 12 pp (in Swedish)
Höglund J, Cortazar Chinarro M, Jarnemo A, Thulin CG (2013) Genetic variation and structure in Scandinavian red deer (Cervus elaphus): influence of ancestry, past hunting, and restoration management. Biol J Linn Soc 109:43–53
Jarnemo A (2008) Seasonal migration of male red deer (Cervus elaphus) in southern Sweden and consequences for management. Eur J Wildl Res 54:327–333
Jayakody S, Sibbald AM, Gordon IJ, Lambin X (2008) Red deer Cervus elaphus vigilance behavior differs with habitat type and human disturbance. Wildl Biol 14:81–91
Jeppesen JL (1987) Impact of human disturbance on home range, movements, and activity of red deer (Cervus elaphus) in a Danish environment. Dan Rev Game Biol 13:1–38
Johnson BK, Ager AA, Noyes JH, Cimon N (2004) Elk and mule deer responses to variation in hunting pressure. In: Transaction of the 69th North American Wildlife and Natural Resources Conference. The Wildlife Institute, Spokane, WA, USA, pp 625–640
Kilpatrick HK, Lima KK (1999) Effects of archery hunting on movement and activity of female white-tailed deer in an urban landscape. Wildl Soc Bull 27:433–440
Krebs H (1969) Schießen oder schonen. München, Mayer
Liberg O, Bergström R, Kindberg J, von Essen H (2010). Ungulates and their management in Sweden. In: Apollonio M, Andersen R, Putman R (eds) European ungulates and their management in the 21st century. Cambridge, Cambridge University Press, p 37–70
Lovari S, Herrero J, Conroy J, Maran T, Giannatos G, Stubbe M, Aulagnier S, Jdeidi T, Masseti M, Nader I, de Smet K, Cuzin F (2008) Cervus elaphus. In: IUCN 2008. 2008 IUCN Red List of Threatened Species. Available from www.iucnredlist.org [accessed on 8 January 2013]
Mattsson L, Boman M, Ericsson G (2008) Jakten i Sverige – Ekonomiska värden och attityder jaktåret 2005/06. Rapport från Adaptiv förvaltning av vilt och fisk. Rapport nr 1 (in Swedish)
McCorquodale SM (2003) Sex-specific movements and habitat use by elk in the cascade range of Washington. J Wildl Manage 67:729–741
Miller JR, Cowles RS (1990) Stimulo-deterrent diversion: a concept and its possible application to onion maggot control. J Chem Ecol 16:3197–3212
Millspaugh JJ, Brundige GC, Gitzen RA, Raedeke KJ (2000) Elk and hunter space-use sharing in South Dakota. J Wildl Manag 64:994–1003
Milner JM, Bonenfant C, Mysterud A, Gaillard J-M, Csanyi S, Stenseth NC (2006) Temporal and spatial development of red deer harvesting in Europe: biological and cultural factors. J Appl Ecol 43:721–734
Mohr CO (1947) Table of equivalent populations of North American small mammals. Am Midl Nat 37:233–249
Mysterud A (2010) Still walking on the wild side? Management actions as steps towards “semi-domestication” of hunted ungulates. J Appl Ecol 47:920–925
Månsson J, Jarnemo A (2012) Bark-stripping on Norway spruce by red deer in Sweden: level of damage and relation to tree characteristics. Scand J For Res DOI:10.1080/02827581.2012.701323. Published online http://www.tandfonline.com/doi/abs
Nelson ME, Mech LD (1991) White-tailed deer movements and wolf predation risk. Can J Zool 69:2696–2699
Nolte D (1999) Behavioral approaches for limiting depredation by wild ungulates. In: Launchbaugh KL, Sanders KD, Mosley JC (eds). Grazing Behavior of Livestock and Wildlife. Idaho Forest, Wildlife and Range Exp. Sta. Bull. #70, Univ. of Idaho, Moscow, ID
Okarma H (1995) The trophic ecology of wolves and their predatory role in ungulate communities of forest ecosystems in Europe. Acta Theriol 40:335–386
Okarma H (1997) Der Wolf. Ökologie, Verhalten, Schutz. Parey Buchverlag, Berlin, in German
Pan D, Teng L, Cui F, Zeng Z, Bravery BD, Zhang Q, Song Y (2011) Eld’s deer translocated to human inhabited areas become nocturnal. Ambio 40:60–67
Phillips GE, Alldredge AW (2000) Reproductive success of elk following disturbance by humans during calving season. J Wildl Manag 64:521–530
Putman R, Langbein J, Green P, Watson P (2011) Identifying threshold densities for wild deer in the UK above which negative impacts may occur. Mamm Rev 41:175–196
Pyke B, Rice M, Sabine B, Zalucki MP (1987) The push-pull strategy - behavioral control of Heliothis. Austral Cott Grow May-July:7–9
Reimoser F, Putman RJ (2011) Impact of large ungulates on agriculture, forestry, and conservation habitats in Europe. In: Putman RJ, Apollonio M, Andersen R (eds) Ungulate Management in Europe: Problems and Practices. Cambridge University Press, Cambridge, UK, pp 144–191
Root BG, Fritzell EK, Giessman NF (1988) Effects of intensive hunting on white-tailed deer movement. Wildl Soc Bull 16:145–151
Scillitani L, Monaco A, Toso S (2009) Do intensive drive hunts affect wild boar (Sus scrofa) spatial behavior in Italy? Some evidences and management implications. Eur J Wildl Res 56:307–318
Sibbald AM, Hooper RJ, McLeod JE, Gordon IJ (2011) Responses of red deer (Cervus elaphus) to regular disturbance by hill walkers. Eur J Wildl Res. doi:10.1007/s10344-011-0493-2
Sunde P, Olesen CR, Madsen TL, Haugaard L (2009) Behavioral responses of GPS-collared female red deer Cervus elaphus to driven hunts. Wildl Biol 15:454–460
Swenson JE (1982) Effects of hunting on habitat use by mule deer on mixed-grass prairie in Montana. Wildl Soc Bull 10:115–120
Thurfjell H, Spong G, Ericsson G (2013) Effects of hunting on wild boar Sus scrofa behavior. Wildl Biol 19:87–93
Vercauteren KC, Hygnstrom SE (1998) Effects of agricultural activities and hunting on home ranges of female white-tailed deer. J Wildl Manag 62:280–285
Verdolin JL (2006) Meta-analysis of foraging and predation risk trade-offs in terrestrial systems. Behav Ecol Sociobiol 60:457–464
Verheyden H, Ballon P, Bernard V, Saint-Andrieux C (2006) Variations in bark-stripping by red deer Cervus elaphus across Europe. Mamm Rev 36:217–234
Vieira ME, Conner MM, White GC, Freddy DJ (2003) Effects of archery hunter numbers and opening dates on elk movement. J Wildl Manag 67:717–728
von Raesfeld F, Vorreyer F (1974) Das Rotwild. Verlag Paul Parey, Hamburg (in German)
Wagenknecht E (1986) Rotwild. Verlag J. Neumann-Neudamm, Berlin (in German)
Wikenros C, Sand H, Wabakken P, Liberg O, Pedersen HC (2009) Wolf predation on moose and roe deer: chase distances and outcome of encounters. Acta Theriol 54:207–218
Williams SC, DeNicola AJ, Ortega IM (2008) Behavioral response of white-tailed deer subjected to lethal management. Can J Zool 86:1358–1366
Acknowledgments
We thank Ove Fransson, John Källström, Johan Palmgren, Bengt O. Röken, Håkan Svensson, and Bosse Söderberg for field work, assistance during marking and information about hunts. Johanna Månsson Wikland, Jenny Mattisson, and Henrike Hensel helped with the administration of GPS-collars. We also thank Jenny Mattisson, Henrik Andrén, Marco Festa-Bianchet, and two anonymous reviewers for valuable comments on earlier versions of the manuscript. The study was financed by The Swedish Association for Hunting and Wildlife Management, Holmen Skog AB, Ittur AB, Virå Bruk AB, Karl-Erik Önnesjös Stiftelse för Vetenskaplig forskning och Utveckling, Stiftelsen Oscar och Lili Lamms Minne, and Kolmårdens insamlingsstiftelse/Tåby Allmänning.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by C. Gortázar
Rights and permissions
About this article
Cite this article
Jarnemo, A., Wikenros, C. Movement pattern of red deer during drive hunts in Sweden. Eur J Wildl Res 60, 77–84 (2014). https://doi.org/10.1007/s10344-013-0753-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10344-013-0753-4