
Abstract
In wolves, most offspring mortality occurs within the first 6–8 months of their life. As wolf pups pass this entire period at either the den or rendezvous sites, their selection by wolf packs may affect pup survival and recruitment. Rendezvous sites are important for pup survival as they are used during summer and early autumn, when intense human activity may increase pup mortality. Adult wolves and pups can be killed by livestock guarding dogs during summer and intentionally or accidentally during large game hunting in autumn. This study describes factors related to rendezvous site selection in order to enhance their protection and management. We studied the rendezvous site selection of 30 wolf packs in central and northern Greece between 1998 and 2010, after locating 35 sites using the simulated howling survey method and telemetry. We considered a series of environmental and anthropogenic predictors of wolf rendezvous site selection at two spatial scales. At the landscape-population scale, wolves selected rendezvous sites below 1,200 m asl, with large inter-site distance (mean, 12.9 km), and avoided partially forested or open habitats, indicating preference for covered, spaced areas with seasonally stable resources. At the home range scale, wolves selected rendezvous sites away from forest roads and villages, close to water sources, and in areas with low forest fragmentation, indicating avoidance of human presence and disturbance. In the summer of 2011, we used an ensuing resource selection model (RSF, AUC = 0.818) to successfully locate seven new rendezvous sites outside our previous survey area, verifying the utility of prediction maps (all new sites were at areas with 0.8–1 model probability). Rendezvous prediction maps can be used to reduce field effort when monitoring wolf populations, assess livestock predation risk, design protected areas, and reduce human disturbance on reproductive wolf packs.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Successful reproduction and pup survival are critical demographic components related to wolf population dynamics and species recovery (Boitani 2000; Fuller et al. 2003). Many studies identified resource availability as the main factor determining wolf reproductive success (e.g., Fuller 1989b; Fuller et al. 2003), while others showed wolf human-caused mortality to be an important inhibitor of wolf reproduction and population recovery (e.g., Liberg et al. 2012). Homesite selection by wolf packs can be closely related to both these factors. It can directly influence access to food resources by reproductive wolf packs (Frame et al. 2008) as well as disturbance of nurturing adults and pups from humans (Frame et al. 2007; Argue et al. 2008; Habib and Kumar 2007; Nonaka 2011). The highest risk of pup mortality occurs during the first 6 months of their age (Fuller et al. 2003) when still occupying homesites (i.e., dens and rendezvous sites). Dens are heavily used during spring for parturition and after birth of pups, while rendezvous sites are selected during summer and early autumn after den abandonment in early summer (Packard 2003). During this entire pup rearing period, the breeding pair and other pack members move continuously to and from dens and rendezvous sites in order to feed and protect the offspring (Alfredeen 2006; Ruprecht et al. 2012). In Greece, the period of rendezvous site use coincides with increased human activities, such as widespread grazing of livestock accompanied by guarding dogs and the onset of wild boar (Sus scrofa) hunting. The increased wolf movements and human presence in this period results in higher encounter frequency between wolf packs and livestock guarding dogs and/or hunters and, as a consequence, a high rate of wolf pup mortality (Iliopoulos 1998, 1999, 2000; Iliopoulos and Petridou 2012). Excessive human disturbance close or inside to homesites may also lead to their abandonment and the translocation of pups (Person and Ingle 1995; Weaver et al. 1996; Frame et al. 2007; Habib and Kumar 2007; Argue et al. 2008).
During environmental impact assessments (EIAs) or the development of management plans for large construction (e.g., highways) or resource extraction (e.g., mining, timber harvest) projects, it is important that local wolf dens and rendezvous sites are identified so as to minimize the activities’ impact on local wolf populations (e.g., Paquet and Darimont 2002; Lesmerises et al. 2013). In Greece, the typically short period of field surveys afforded to EIAs for such projects means that wolf homesite detection may be incomplete. Moreover, stochastic events such as wolf pack breeder loss which may cease wolf reproduction for some years in an area (Brainerd et al. 2008) may hinder homesite detection during field surveys. The ability to predict the location of wolf homesites using environmental and anthropogenic parameters would help address such problems, contributing to the improved development of wolf-sensitive action timetables. Moreover, such prediction models would be particularly useful in identifying priority sites for wolf conservation initiatives, assessing livestock predation risk (Gula 2008; Treves et al. 2011), and improving the efficiency of wolf population monitoring schemes over large areas (Ausband et al. 2010; Stenglein et al. 2010).
While dens are especially important for pups, in human modified landscapes rendezvous sites may play an equally important role in wolf pup survival (Paquet and Darimont 2002; Fritts et al. 1995). Recent research has shown topographic landscape features, vegetation structure and habitat type, presence of water, human disturbance, and protection status of an area to be good predictors of wolf rendezvous site (e.g., Norris et al. 2002; Theuerkauf et al. 2003; Capitani et al. 2006; Person and Russell 2009; Unger et al. 2009; Ausband et al. 2010; Kaartinen et al. 2010). Until now, in Europe, rendezvous site selection has been studied only in Poland (Theuerkauf et al. 2003) and Italy (Capitani et al. 2006). To fill this gap in Greece, we examined between 1998 and 2010 the rendezvous site selection patterns of thirty wolf packs—a large proportion of the estimated 100 packs in the country (Iliopoulos 2009). The analysis aims to identify the most important landscape attributes, resource availability variables, and anthropogenic landscape features affecting rendezvous site selection at the (a) landscape-population and (b) home range scale. Additionally, we test the hypothesis that wolves select rendezvous sites primarily in areas where pups will be safer from human-caused mortality, predicting that distance from roads, human settlements, and less fragmented forest landscapes would be positively related to rendezvous site selection. Finally, we examine the robustness of the home range resource selection model and its implications for management purposes, using a posteriori original field data.
Materials and methods
Study area
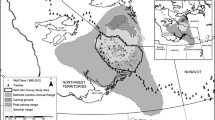
Surveys for locating wolf rendezvous site occurred in two regions in central and northern Greece covering a total area of 6,700 km2 (Fig. 1). The a posteriori test of the generated rendezvous site predictive model was conducted at three small “out of sample” areas with a total surface of 300 km2 (Fig. 1). Across all these areas, elevation ranges from 100 to 2,500 m and temperature from −20 to 38 °C. Evergreen oak (Quercus coccifera) scrublands dominate areas up to 700 m elevation, deciduous oak (Quercus sp.) forests are abundant between 600 and 1,100 m, conifer forests of fir (Abies borisii regis) and black pine (Pinus nigra) above 1,000 m, and beech (Fagus sylvatica)–fir mixed forests the 1,000–1,800 m zone. Scrublands and forests cover 65 % of the study area, with remaining being natural grasslands and agricultural or mixed agricultural–forest areas. Average human population density, excluding cities, is 15 inhabitants/km2 (data for 1999: Ministry of Interior). Village density ranges from 0.6 to 5.4 settlements/100 km2. Livestock grazing is practiced throughout the study area, and shepherd dogs are widely used for livestock protection from large carnivores (i.e., wolf and brown bear Ursus arctos). The 100–500 m elevation zone is primarily used as wintering area of nomadic livestock flocks from October to May; these flocks are moved to the 1,200–2,400 m elevation zone. At the 500–1,000 m zone, grazing is practiced by residing free-ranging livestock flocks year-round. Average density of sheep and goats is 67.21, cattle 2.55, and free-ranging pigs 2.9 heads/km2. The forest road network is extensive, with a mean density of 2.43 ± 0.63 km/km2. Hunting for birds and mammals lasts 6 months, beginning from August 20 and is practiced with trained hunting dogs for birds and hare (Lepus europaeus) and also with drive hunts for wild boar. Hunting is prohibited inside the Northern Pindos National Park, Vikos-Aoos National Park core areas, and 47 smaller reserves covering in total 12 % of the study area.

Map of the study areas showing a) main sampling areas (dashed polygons, 6,700 km2) monitored during 1998 and 2010 to locate rendezvous sites used for analysis and creation of a resource selection model and b) “out of sample” areas (double line polygons, 300 km2) monitored during summer of 2011, which were used to check the robustness of the RSF model. Solid line polygons indicate home ranges (minimum convex polygons) of radio-collared packs studied between 1998 and 2011. Black dots represent rendezvous sites (n = 35) of 30 different wolf packs that were used for modeling rendezvous site selection
Rendezvous site surveying
Surveying for wolf rendezvous site took place between 1998 and 2010, and the ensuing model was tested in the field in 2011. Six out of the 30 packs were tracked using satellite and radio-telemetry, permitting direct investigation of putative rendezvous sites. Each year between July and September, rendezvous site surveying and confirmation was carried out by the “simulated howling survey method” in a saturated census (Harrington and Mech 1982). This time is preferred as the wolf packs are still sedentary (Packard 2003), the distinction of pup howls from those of adults is feasible (Harrington and Asa 2003), and the response rate is high (Harrington and Mech 1982). We applied the method in all 30 pack areas, where and when recent field data confirmed wolf presence, so as to minimize false-absence records, a common problem in used versus unused studies (Boyce et al. 2002). Field evidence included tracks of two or more wolves, markings, direct sightings of wolves or wolf pups, livestock damage reports, and records from camera traps. When a wolf pack response was elicited, the azimuth of the howl(s) was recorded and the distance to responding wolves estimated during daylight revisits. A site (n = 35) was classified as a rendezvous site based on presence of pups when (a) a wolf pup response was heard at least twice during the same summer period (n = 29 sites) and (b) only adult wolves were heard but pup presence was verified by other field evidence, such as captured or dead pups, direct sightings, and images of pups from camera traps (n = 6 sites). To distinguish among located neighboring rendezvous sites, we repeated the simulated howling protocol during the same night. Nevertheless, in all cases, inter distance was longer than the threshold of 5 km suggested by Capitani et al. (2006) or 6.4 km by Ausband et al. (2010).
We included a rendezvous site in the analysis only once, even if it had been reused by a pack over multiple years. For each rendezvous site (n = 35), we randomly selected a non-overlapping unused site (Hawth’s tools 3.7 for ArcGIS 211 9.3), at distances ≤5 km, in order to minimize the probability to be located outside of the respective wolf pack home range. Both used and unused sites were considered as circular surfaces, with a radius of 800 m (area of 2 km2), as average size of known rendezvous sites from collared wolf packs during this study was 2.2 ± 0.39 km2 (n = 8 wolves). “Unused” sites located over lakes, heavily disturbed areas, and purely agricultural or open habitat areas were excluded from all analyses.
Data analysis
We initially analyzed the selection of rendezvous sites at the landscape-population scale (second order selection; Johnson 1980) by comparing the use of rendezvous sites versus the availability of three major variables in the study area: (a) livestock summer density (June-October) in three classes: 0–32, 33–79, and >80 heads/km2; (b) elevation zones in four classes: 100–500, 501–800, 801–1,200, and 1,201–2,500 m, classified with the quantile method (ArcGis 9.3, ESRI); and (c) Corine landuse types grouped in six classes: evergreen scrublands, deciduous forests, conifer forests, transitional scrubland to forest, mixed agricultural–forest habitats, and natural grasslands. It was not possible to include more variables in the analysis, as detailed spatial data at this scale were lacking. We determined the selection ratio (SR) for each variable class (units = 1 km2) as follows (Manly et al. 2002):
where o i is the proportion of the ith class sampled inside rendezvous sites and π i is the proportion within the study area. W i >1 indicates preference, while W i <1 indicates avoidance of the i class. We calculated Bonferroni-corrected confidence intervals (Manly et al. 2002) as follows:
where 1 is the number of classes per variable, α = 0.1, and SE is the standard error of W i . The SE(W i ) equation takes into account variation in resource selection from all different rendezvous sites (Rogers and White 2007). If confidence intervals overlap with 1, i class is used according to availability.
We then analyzed rendezvous site selection at the home range scale (third order selection; Johnson 1980) by comparing rendezvous and unused sites with multivariate logistic regression (Hosmer and Lemeshow 1989; Capitani et al. 2006; Ausband et al. 2010). For each rendezvous and control site pair, we measured habitat and landscape characteristics (Table 1) at a resolution of 10 × 10 m, using Aster satellite images, DTM military maps, and Corine landuse maps in ArcGis 9.3 (ESRI). Forest roads and active farms were mapped in the field. Livestock density was spatially adjusted according to the actual location of active farms. Perennial water sources were mapped from military maps of 1:50,000 scale (Hellenic Geographical Military Service). Forest fragmentation indices were estimated with Patch Analyst 4 (Rob Rempel, Centre for Northern Forest Ecosystem Research, Canada) for ArcView3.2 (ESRI). We regrouped Corine landuse maps to describe vegetation form, independent of habitat type. Categorical covariates entered models as “dummy” variables.
We conducted analysis including all variables in a full model and sequentially removed covariates with lowest b/SE ratio, until variable reduction ceased to improve the model performance (Arnold 2010). Selected covariates were further combined to create candidate models. Variables correlated at Pearson’s r > |0.5| were tested separately. We selected as best model the most parsimonious, statistically significant model (Hosmer and Lemeshow χ 2 test), with the largest R 2 Nagelkerke, R 2 Cox and Snell values (Boyce et al. 2002) and overall percent of correct classification of RS. For each variable i included in the model, we estimated the R i statistic (Field 2005) which describes the relative importance of each variable with the following formula:
where b i is the slope of variable i, SE i is the standard error of b i , df the degrees of freedom of i, and LL is the log likelihood.
Performance was evaluated with integration of area under curve (AUC) after the calculation of the ROC (receiver operating characteristic curve) according to (Boyce et al. (2002). All statistical analyses were performed with SPSS 13 (SPSS Inc, Chicago, USA).
For a preliminary, out of sample, testing of model robustness, we calculated rendezvous site suitability at three smaller new areas of 300 km2 (Fig. 1). We created a grid surface covering the test areas with a cell size of 70 × 70 m. We then created raster surfaces with the same extent and cell size to visualize ecogeographical variables that entered the final logistic regression model (see Table 4). Variable raster surfaces had a buffer zone of 1,000 m around the study area boundaries so as to account for no-data values during calculations. At each cell, we assigned a unique value derived by each variable raster surface. An 800-m-radius moving window (FRAGSTATS 4.1, McGarigal et al. 2002) was used to estimate forest fragmentation covariate values for each grid cell considered in the final model (TCAI index). We calculated the probability of each reference grid cell constituting the center of a rendezvous site using the logistic regression equation derived from the final model. Thus, each cell of the test study areas was assigned a probability value ranging from 0 to 1. To produce the final suitability rendezvous site maps, we rerun the moving window procedure at a radius of 800 m (corresponded to a rendezvous site of 2 km2 in size) around each grid cell in order to smooth the probability maps by averaging probability values of neighboring cells included at that specific radius. Calculated rendezvous site suitability ranged accordingly from 0 to 1. We then reclassified probability with the equal interval method that creates classes within an equal probability range of 0.1 to produce prediction maps with ten suitability zones (Fig. 2). All analyses were performed using ArcGiS 9.3 (ESRI).

Rendezvous site prediction map (70 × 70m grid cell size) in the three “out of sample” test study areas monitored in summer of 2011, after application of the home range-scale logistic regression equation, derived from resource selection analysis of 35 rendezvous sites used by 30 wolf packs in the main study area. Probability classes (n = 10) were created with the equal-probability interval method. Closed circles of 800 m radius (2 km2) represent new rendezvous sites (n = 7) used by four wolf packs, while black dots represent their center. All new rendezvous sites located in the field had their center in high suitability areas (0.8–1) and their overall average suitability was 0.736 ± 0.096 (SD)
During the summer 2011, we carried out a saturated census to locate rendezvous sites currently occupied or/and used during the last 2–3 years as to increase sample size of rendezvous sites after verifying previous use when possible. We initially undertook a preliminary field assessment census during July 2011 to define areas where wolf packs were present. During this assessment, we mapped all available direct and indirect information indicating presence of wolf packs, such as spatially localized wolf signs (scats, tracks, markings, diggings), and cross referenced information on wolf group howling and pup sightings provided by local shepherds and hunters. During August and September 2011, we undertook a saturated simulated howling survey (Harrington and Mech 1982) in the three new study areas where current wolf presence was mapped during preliminary census. We used the same two evaluation criteria for classification of sites as rendezvous ones, when wolf howling was heard as previously described. When a wolf response was not elicited at a site but other strong indications of wolf reproduction were evident (e.g., farmer reports on pup sighting, howling and presence of dens), we visited—if possible—the sites for identification of field signs suggesting use by wolf pups. Field surveys were conducted independently from GIS model interpretation.
We calculated percent of use of each suitability class i, inside new RS and the percent of standardized selection ratio B i derived from w i as follows:
Results
During the 1998–2010 survey, we identified a total of 35 rendezvous sites belonging to the 30 different wolf packs studied. We were able to monitor reuse pattern in ten of these rendezvous sites, belonging to nine more intensively studied packs, by checking if those were repeatedly occupied by adult wolves and pups. Checks were performed with the same protocol for at least two different summers per site (range = 2–6) but not necessarily during consecutive years. Use and reuse history of these ten rendezvous sites is presented in Table 2. Seven of these ten rendezvous sites incorporated also an active den inside or at a relatively close distance (<2 km). Dens were located with the use of telemetry data and presence of wolf pups verified with simulated howling in six cases and camera trap pictures in one case. Average inter-rendezvous site distance between different wolf packs was 12.9 ± 1.7 km (range = 10.7–16.6 km, n = 12 pairs) and did not significantly differ between sub-portions of the study area (Kruskal–Wallis test, H = 2.682, df = 3, exact P = 0.509, n = 4).
At the landscape-population scale, wolves did not select any livestock density zone but avoided establishing rendezvous sites at altitudes >1,200 m and mixed agricultural–forest habitat or grasslands. There was a nonsignificant tendency for selecting the 501–800 m zone and the evergreen scrubland zone (see Table 3).
At the third order selection (home range scale), four variables entered the final model (Table 4). Wolves selected rendezvous sites farther from forest roads (P = 0.007, mean ± SD = 435 ± 326 m, range = 73–1614 m), closer to a permanent water source (P = 0.012, mean ± SD = 500 ± 434 m, range = 0–1,707 m), in less fragmented forested areas (P = 0.040), and farther from villages (P = 0.019, mean ± SD = 2,819 ± 1,234 m, range = 1,047–6,026 m). The model correctly predicted 80 % of rendezvous sites and according to the estimated AUC value (0.818, SE = 0.051) can be considered a “useful application” (see Boyce et al. 2002).
In summer 2011, we identified seven new rendezvous sites of 2 km2 area each in the “out of sample” study areas (Fig. 2). Although total surface of the “out of sample” evaluation areas was small (300 km2), the rendezvous sites were spaced at large distances apart from each other (>10.2 km). Identified rendezvous sites corresponded to four different wolf packs. Five of these seven sites where currently occupied by wolves during the 2011 field census and successfully located with the simulated howling survey method (Fig. 2). One of the sites was occupied by wolves the previous year (2010). Its actual use by a wolf pack was verified after inspection of many diggings, spatially concentrated food remains still present in the area, and/or other objects carried by wolves. Moreover, a wolf den was demonstrated in the field by a local shepherd, 1.4 km away from the rendezvous site, who had discovered the den and seen the pups in early June 2010. Although we did not find any signs of current wolf activity in the seventh rendezvous site, there were cross-referenced reports related to pup rearing from several farmers in adjacent areas, who characterized the site as a “traditional” wolf homesite. Indications of rendezvous use included wolf group howling and many wolf pup sightings during summers between 2007 and 2009. Relevant information was considered adequately sufficient as to finally include also this site in the analyses.
All new rendezvous sites were located in high-suitability classes (mean ± SD = 0.736 ± 0.096, range = 0.6–0.86), as derived from probability prediction maps (Fig. 2). The predominant probability class was the 0.8, while standardized percent selection ratios (%B i ) were highly correlated to suitability levels (r s = 0.988, P < 0.01, Fig. 3). All rendezvous sites had their centers in areas with estimated probability >0.8, which totally covered 9.9 % of the “out of sample” study areas.

Percentage distribution (dark gray bars) of the total surface of the new rendezvous sites (n = 7) located in the “out of sample areas” at each probability class, after assigning predicted probability values to each rendezvous site cell (70 × 70m) derived from rendezvous site probability prediction maps, and standardized %B i values (light gray bars). B i is the standardized value (range, 0–100 %) of each probability class selection ratio w i inside the new rendezvous sites (see “Materials and methods” for equation used)
Discussion
Our analyses of rendezvous site selection produced some interesting results that have been recorded for the first time in Greece. At the population-landscape scale, wolves avoided altitudes >1,200 m, while there was a nonsignificant tendency to prefer the medium altitude zone (501–800 m) to establish rendezvous sites. In Greece, high altitudes (>1,200 m) are characterized by more food sources, but this is highly seasonal and food abundance peaks only during summer. On the other hand, lower altitudes (<1,200 m), and especially the 501–800 m zone, are characterized by less fluctuation in food sources, as livestock, currently the main wolf prey in Greece (Papageorgiou et al. 1994; Migli et al. 2005; Iliopoulos et al. 2009), graze year round in such areas. Although, some authors found that proximity of homesites to summer food sources increases pup survival, no direct links between homesite location and food abundance were recorded (Heard and Williams 1992; Heard et al. 1996; Frame et al. 2008). Breeding wolves may establish homesites even in areas far from abundant food resources and increase travel distance to feed offspring (Walton et al. 2001; Frame et al. 2008). In contrast, it seems that stability of food resources is more important and has been linked to den and rendezvous site selection (Ciucci and Mech 1992).
This is further supported by the lack of any significant influence of mean livestock density on rendezvous site selection. According to our findings, rendezvous sites were not related to prey abundance and were not located in proximity to each other, as shown by the low variation of the in-between rendezvous site distances, a pattern also encountered in northern Italy (Capitani et al. 2006). In fact, concentration of wolf rendezvous sites at areas with high livestock densities would result in respectively high inter-pack aggression and intra-specific mortality (Ciucci and Mech 1992). Appropriate spacing, to combine optimum distance from scattered food sources and reduced inter-pack aggression (Mech et al. 1998; Mech and Harper 2002), could be an important factor of rendezvous site spatial distribution apart for environmental and anthropogenic factors.
Regarding habitat types, wolves did not particularly select any specific forest habitat but generally avoided semi-forested areas and open habitats to establish rendezvous sites (w i = 0.52–0.57). Forest habitats offer greater safety from human disturbance and are usually selected for homesite establishment, even when open habitats dominate the landscape (Cortes 2001). In addition, evergreen scrublands were slightly preferred (w i = 1.38), although this preference was not statistically significant as dense or young forests appear to provide excellent hiding cover for wolves at homesites (Kaartinen et al. 2010).
At the home range scale, proximity to forest roads emerged as the most important factor (R = 0.232), determining rendezvous site selection. As road proximity implies increased human disturbance, rendezvous sites were located away from forest roads at a high probability, corroborating results from studies in Italy (Ciucci et al. 1997; Capitani et al. 2006), Poland (Theuerkauf et al. 2003) and in a, heavily disturbed by logging, area in the USA (Person and Russell 2009). In effect, Karlsson et al. (2007) found that simple human presence in an area may trigger wolves to abandon it. Nonaka (2011) found that adult breeding wolves always moved away from dens up to 5,600 m after experimental disturbance at homesites and, in most cases, pups were translocated to a new den. Frame et al. (2007) found that pups were moved by adults to secondary dens in 50 % of all studied cases, following experimental disturbance. Nevertheless, this response was more related to pup age at the time of disturbance, as also found in India (Habib and Kumar 2007). Although wolves do not seem to tolerate human disturbance close to homesites (Frame et al. 2007; Argue et al. 2008; Habib and Kumar 2007; Nonaka, 2011), no effects between experimental human disturbance and survival of wolf pups were found (Frame et al. 2007; Argue et al. 2008; Person and Russell 2009). Wolves reused disturbed homesites at a similar frequency to undisturbed homesites, the years following disturbance (Frame et al. 2007; Argue et al. 2008; Person and Russell 2009). It is reasonable to assume that higher mobility of wolf pups in summer and early autumn makes them less vulnerable to human disturbance close to rendezvous sites as they can flee more easily from approaching humans compared to younger pups.
Nevertheless, in all aforementioned studies, experimental human disturbance did not include mortality of wolf pups or adult wolves. In contrast to experimental disturbance, in other cases when local residents are involved, presence of humans close to homesites may result in direct killing or removal of pups from dens (Jedrzejewska et al. 1996). Forty percent of wolves, aged less than 1 year in Greece, were killed inside homesites (Iliopoulos 1998), using the most commonly method, drive hunts. In fact, drive hunts are responsible for 42 % of overall human-caused mortality of adult wolves and 35 % of that of wolf pups (Iliopoulos 1998). The method consists of a hunting group walking inside wolf resting areas, with or without hunting dogs, and driving wolves in specific stalking points, similar to those for wild boar. This method is facilitated by forest roads which permit a quick census of wolf tracks around resting sites and an easier access of the hunting team. Road density was also highly correlated to increased harvest of wolves in southern Alaska (Person et al. 1996). In Bialowieza primeval forest, where a negative relation was similarly found between proximity to roads and rendezvous site selection (Theuerkauf et al. 2003), wolf pup human-caused mortality inside homesites has historically constituted an important factor of wolf mortality (Jedrzejewska et al. 1996). In our case, road avoidance by reproductive wolf packs, when selecting for rendezvous sites, may consist an adaptive strategy against hunting of breeding wolves and offspring in Greece, rather than a response to mere human presence and temporal disturbance.
The response to a particular disturbance seems to also depend on disturbance history, a critical concept in understanding the behavior of long-lived animals that learn through social transmission (Curatolo and Murphy 1986). Although this is the case for areas with increased anthropogenic impact and a long history of wolf persecution, in other areas where legal protection is implemented and human populations are low, no relations between distance from forest roads and rendezvous site selection has been detected (Unger et al. 2009). In such areas, some wolves seem to tolerate more human presence (Thiel et al. 1998).
Distance from villages was also related to anthropogenic disturbance, but with lower influence (R = 0.151) on rendezvous site selection. Wolves selected sites that were farthest from villages, as also reported by Theuerkauf et al. (2003) and Capitani et al. (2006). However, in Greece, this factor was the least important because wolves frequent the vicinities of villages for food, such as garbage or dead livestock (Iliopoulos, unpublished data).
Proximity to water appeared to be the second most important factor (R = −0.209). Similar findings have been also reported by Unger et al. (2009) and Ausband et al. (2010). Proximity to water sources reduces the need for pups to travel longer distances and therefore expose themselves to fatal danger in order to drink water, which is essential for their survival, especially during summer and autumn dry months.
Another critical factor for rendezvous site selection at the home range scale was forest fragmentation (R = 0.172). In our study sites, wolf packs selected for homogenous, less fragmented forested areas (see also Theuerkauf et al. 2003). In larger, more homogenous forests (higher TCAI index), potential predators (e.g., humans, livestock guarding dogs) need to travel longer distances inside vegetated areas to reach the centers of rendezvous sites and are subject to easier auditory detection by wolves as has been also experimentally demonstrated (Wam 2003; Karlsson et al. 2007).
Rendezvous sites appeared to be rather traditional, as a high rate of reuse was detected: the most notable cases involved reuse after 12 years and consecutive use for 5 years, while in total eight out of ten sites were reused for at least 2 years (Table 2). Wolf pack #5 used the same den and rendezvous site for at least five consecutive summers (2002–2007), whereas, according to local farmer testimonies, wolves bred there even earlier. After illegal killing of at least one breeder wolf in winter 2008, wolf reproduction ceased in that area. Forest network use at this homesite was restricted to public vehicles, with a system of metal bars set by the local hunting club to reduce hunting pressure. Moreover, livestock grazing was prohibited by forest service to protect black pine reforestations. Although we lack data on wolf reproduction prior to forest road closing in that area, consistent reuse of the den, and the associated rendezvous site, could be linked to low human presence. Another interesting reuse case of a rendezvous site that also encompassed a den in the same area was by wolf pack #1. Wolf reproduction was located for the first time during the onset of the study in spring and summer 1999. Although both radio-collared breeding wolves were illegally killed by wild boar hunters in fall 1999, reuse of the homesites was recorded after 12 years most probably by wolves unrelated to that first breeding pair.
Reuse rates of rendezvous sites in our study area are in partial contrast to results from Poland (Theuerkauf et al. 2003) or Finland (Kaartinen et al. 2010), where wolves used different homesites. Rendezvous site reuse has been associated with breeder persistence (Capitani et al. 2006) or with proximity to important or limited resources (Ballard and Dau 1983; Ciucci and Mech 1992). In our case, recorded reuse, apart from breeder persistence, may be also related to shortage of high suitability habitat for establishing new rendezvous sites. In fact, prediction maps at test areas showed that only 9.9 % could be suitable for rendezvous site core areas. This estimate may be even smaller, considering the variance not explained by the suitability model. As wolves appeared to select against human disturbance, and our study sites were located in areas with high human presence and very high forest road density, site reuse appeared as an ecologically safe option (see also Fernández et al. 2012).
Conclusions and management implications
The current study on wolf rendezvous site selection in northern and central Greece showed that avoidance of human presence and disturbance accounted more than factors related to habitat types or availability of prey. Wolf preference to livestock in our study area and the lack of any spatial association of rendezvous sites with prey availability are not contradictory, as free-ranging livestock also implies high human presence and disturbance. This strategy, adopted by wolves in Greece, may be a response to the long history of human persecution, still ongoing and widespread, and is consistent with countries with similar histories, such as Poland and Italy. This was more evident at the home range scale. On the other hand, at the landscape scale, wolves established rendezvous sites in a way to minimize intra-specific competition and maximize access to stable resource availability, a trend more commonly encountered among wolves (Ballard and Dau 1983; Fuller 1989a; Ciucci and Mech 1992).
Premium rendezvous site habitat appeared as a rather limited resource in our study area. This was expressed by the consecutive reuse of rendezvous sites by wolf packs and was highly related to high road density. Thus, control of forest road use could be an effective management measure to assist wolf reproduction in critical areas, such as sink areas with high human-caused mortality. A seasonal restriction on forest road use, implemented in parts of the study area to reduce hunting pressure, was also beneficial for wolves. Limited use of established road networks or careful planning of road network construction, along with maintenance of unfragmented forest patches, should always accompany any plans for establishment of protected areas in Greece, as by themselves do not seem to guarantee any particular positive effects on wolf demography. This perspective has been increasingly important for wildlife protection in national parks (Ament et al. 2008).
In the field, relatively accurate location of rendezvous sites was feasible through the howling survey method, which permitted cost-effective sampling of an increased number of different wolf packs and the creation of an RSF model suitable for large area assessment. As the RSF model robustness was adequate, it can be used with confidence to substantially concentrate wolf monitoring efforts on about 10 % of areas under consideration. A similar reduction (89 %) has been also reported by Ausband et al. (2010). Bearing this in mind, rendezvous site prediction maps can be variably useful during EIA studies. However, these maps are insufficient to predict and avoid permanent den site habitat modification or loss, caused by major infrastructure works.
In contrast, our rendezvous site prediction maps are valuable when permanent loss of rendezvous site habitat is in question. In Greece, they can be used for designing protected areas or for mapping livestock depredation risk. This is especially important since increased use of rendezvous sites partially coincides with the hunting season and human disturbance peaks or with attack peaks in autumn, respectively (Iliopoulos et al. 2009). Finally, when seasonal disturbance is considered, the assessment of rendezvous site maps should include individual and detailed disturbance time frames, as packs respond and are affected differently depending on the age of pups and the duration, season, and levels of disturbance (Habib and Kumar 2007; Argue et al. 2008; Person and Russell 2009).
References
Alfredeen AC (2006) Denning behavior and movement pattern during summer of wolves, Canis lupus, on the Scandinavian Peninsula. Grimso Institute of Wildlife Biology
Ament R, Clevenger AP, Yu O, Hardy A (2008) An assessment of road impacts on wildlife populations in US National Parks. Environ Manage 42(3):480–496
Argue AM, Mills KJ, Patterson BR (2008) Behavioural response of eastern wolves (Canis lycaon) to disturbance at homesites and its effects on pup survival. Can J Zool 86:400–406
Arnold TW (2010) Uninformative parameter and model selection using Akaike's information criterion. J Wildl Manag 74:1175–1178
Ausband DE, Mitchell MS, Doherty K, Zager P, Mack M, Holyan J (2010) Surveying predicted rendezvous sites to monitor gray wolf populations. J Wildl Manag 74:1043–1049
Ballard B, Dau JR (1983) Characteristics of gray wolf (Canis lupus) den and rendezvous sites in south-central Alaska. Can Field Nat 97:299–302
Boitani L (2000) Action plan for the conservation of wolves in Europe (Canis lupus). Nature and Environment, No 113. Council of Europe Publishing, Strasbourg
Boyce MS, Vernier PR, Nielsen SE, Schmiegelow KA (2002) Evaluating resource selection functions. Ecol Model 157:281–300
Brainerd SM, Andren H, Bangs EE, Bradley EH, Fontaine JA, Hall W, Iliopoulos Y, Jimenez MD, Jozwiak EA, Liberg O, Mack C, Meier T, Niemeyer C, Pedersen HC, Sand H, Schultz R, Smith DW, Wabakken P, Wydeven A (2008) The effects of breeder loss on wolves. J Wildl Manag 72:89–98
Capitani C, Mattioli L, Avanzinelli E, Gazzola A, Lamberti P, Mauri P, Scandura L, Viviani A, Apollonio M (2006) Selection of rendezvous sites and reuse of pup raising areas among wolves Canis lupus of north-eastern Apennines, Italy. Acta Theriol 51:395–404
Ciucci P, Mech LD (1992) Selection of wolf dens in relation to winter territories in northeastern Minnesota. J Mammal 73:899–905
Ciucci P, Boitani L, Francisci F, Andreoli G (1997) Home range, activity and movements of a wolf pack in central Italy. J Zool 243:803–819
Cortes Y (2001) Ecologia y conservacion del lobo (Canis lupus) en medios agrıcolas. PhD thesis, University Complutense of Madrid
Curatolo JA, Murphy SM (1986) The effects of pipelines, roads, and traffic on the movements of caribou, Rangifer tarandus. Can Field Nat 100:218–224
Fernández N, Selva N, Yuste C, Okarma H, Jakubiec Z (2012) Brown bears at the edge: modeling habitat constrains at the periphery of the Carpathian population. Biol Conserv 153:134–142
Field AP (2005) Discovering statistics using SPSS, 2nd edn. SAGE, London
Frame PF, Cluff DF, David SH (2007) Response of wolves to experimental disturbance at homesites. J Wildl Manag 71:316–320
Frame PF, Cluff HD, Hik DS (2008) Wolf reproduction in response to caribou migration and industrial development on the central Barrens of mainland Canada. Arctic 61:134–142
Fritts SH, Bangs EE, Fontaine JA, Brewster WG, Gore JG (1995) Restoring wolves to the northern Rocky Mountains of the United States. In: Carbyn LN, Fritts SH, Seip DR (eds) Wolves in a changing world. Canadian Circumpolar Institute, Occasional Publication No. 35, Edmonton, pp 107–125
Fuller TK (1989a) Denning behavior of wolves in north-central Minnesota. Am Midl Nat 121:184–188
Fuller TK (1989b) Population dynamics of wolves in North-Central Minnesota. Wildl Monogr 80:1–41
Fuller TK, Mech LD, Cochrane JF (2003) Wolf population dynamics. In: Mech LD, Boitani L (eds) Wolves, behaviour, ecology and conservation. University of Chicago Press, Chicago, pp 161–191
Gula R (2008) Wolf depredation on domestic animals in the polish Carpathian mountains. J Wildl Manag 72:283–289
Habib S, Kumar J (2007) Den shifting by wolves in semi-wild landscape in Deccan Plateau Maharashtra, India. J Zool 272:259–265
Harrington FH, Asa CS (2003) Wolf communication. In: Mech LD, Boitani L (eds) Wolves, behaviour, ecology and conservation. University of Chicago Press, Chicago, pp 66–103
Harrington FH, Mech LD (1982) An analysis of howling response parameters useful for wolf-pack censusing. J Wildl Manag 6:686–693
Heard DC, Williams TM (1992) Distribution of wolf dens on migratory caribou ranges in the Northwest Territories, Canada. Can J Zool 70:1504–1510
Heard DC, Williams TM, Melton DA (1996) The relationship between food intake and predation risk in migratory caribou and implications to caribou and wolf population dynamics. Rangifer Spec Issue 2:37–44
Hosmer DW, Lemeshow S (1989) Applied logistic regression. Wiley, New York
Iliopoulos Y (1998) Monitoring of wolf population in Greece. Population estimates and human caused mortality. Project report. Project LIFE “Lycos”, NAT 97/GR/04249 Arcturos, EC DGXI, Ministry of Agriculture (in Greek)
Iliopoulos Y (1999) Monitoring of wolf population in Central Greece. Estimates of wolf population and human caused mortality. First annual interim report. Project LIFE “Lycos”, NAT 97/GR/04249 Arcturos, EC DGXI, Ministry of Agriculture (in Greek)
Iliopoulos Y (2000) Monitoring of wolf population in Central Greece. Estimates of wolf population and human caused mortality. Second annual interim report 2000. Project LIFE “Lycos” NAT 97/GR/04249, Arcturos, EC DGXI, Ministry of Agriculture (in Greek)
Iliopoulos Y (2009) Canis lupus, wolf. In: Legakis A, Maragou P (eds) Red data book of threatened animals of Greece. Hellenic Zoological Society, Athens, pp 389–390 (in Greek)
Iliopoulos Y, Petridou M (2012) Preliminary assessment on large carnivore-livestock conflicts in newly established populations of wolf and brown bear in the National Park of Oiti-Central Greece. Management authority of Oiti NP, Callisto NGO (in Greek)
Iliopoulos Y, Sgardelis S, Koutis V, Savvaris D (2009) Wolf depredation on livestock in central Greece. Acta Theriol 54:11–22
Jedrzejewska B, Jedrzejewski W, Bunevich AN, Minkowski L, Okarma H (1996) Population dynamics of wolves Canis lupus in Bialowieża Primeval Forest (Poland and Belarus) in relation to hunting by humans, 1847–1993. Mamm Rev 26:103–126
Johnson DH (1980) The comparison of usage and availability measurements for evaluating resource preference. Ecology 61:65–71
Kaartinen S, Luoto M, Kojola I (2010) Selection of den sites by wolves in boreal forests in Finland. J Zool 281:99–104
Karlsson J, Eriksson M, Liberg O (2007) At what distance do wolves move away from an approaching human? Can J Zool 85:1193–1197
Lesmerises F, Dussault C, St-Laurent MH (2013) Major roadwork impacts the space use behaviour of gray wolf. Landsc Urban Plan 112:18–25
Liberg O, Chapron G, Wabakken P, Pedersen HP, Thompson Hobbs N, Sand H (2012) Shoot, shovel and shut up: cryptic poaching slows restoration of a large carnivore in Europe. Proc R Soc Lond B 279:910–915
Manly BFJ, McDonald LL, Thomas DL, McDonald TL, Erickson WP (eds) (2002) Resource selection by animals, 2nd edn. Kluwer Academic, The Hague
McGarigal K, Cushman SA, Neel MC, Ene E (2002) FRAGSTATS: spatial pattern analysis. Program for Categorical Maps, University of Massachusetts
Mech LD, Harper EK (2002) Differential use of a wolf, Canis lupus, pack territory edge and core. Can Field Nat 116:315–316
Mech LD, Adams LG, Meir TJ, Burch JW, Dale BW (1998) The wolves of Denali. University of Minnesota Press, Minneapolis
Migli D, Youlatos D, Iliopoulos Y (2005) Winter food habits of wolves in central Greece. J Biol Res 4:217–220
Nonaka Y (2011) Response of breeding wolves to human disturbance on den sites—an experiment. Msc Thesis. Grimso forskingsstaion, Uppsala University
Norris DR, Therberge MT, Therberge JB (2002) Forest composition around wolf (Canis lupus) dens in eastern Algonquin Provincial Park, Ontario. Can J Zool 80:866–872
Packard JM (2003) Wolf behaviour, reproductive, social and intelligent. In: Mech LD, Boitani L (eds) Wolves, behaviour, ecology and conservation. University of Chicago Press, Chicago, pp 317–340
Papageorgiou N, Vlachos C, Sfougaris A, Tsachalidis E (1994) Status and diet of the wolf in Greece. Acta Theriol 39:411–416
Paquet PC, Darimont D (2002) Yeo Island wolf home site recommendation. The Heiltsuk Nation, Western Forest Products, Raincoast Conservation Society
Person K, Russell AL (2009) Reproduction and den selection by wolves in a disturbed Landscape. Northwest Sci 83:211–224
Person DK, Kirchhoff M, Van Ballenberghe V, Iverson GC, Grossman E (1996) The Alexander Archipelago wolf: a conservation assessment. United States Department of Agriculture-Forest Service. General Technical Report. PNW-GTR-384. Portland
Rogers KB, White GC (2007) Analysis of movement and habitat use from telemetry data. In: Guy CS, Brown ML (eds) Analysis and interpretation of freshwater fisheries data. American Fisheries Society, Bethesda
Ruprecht JS, Ausband DE, Mitchell MS, Garton EO, Zager P (2012) Homesite attendance based on sex, reproductive status and number of helpers in gray wolf packs. J Mammal 93:1001–1005
Stenglein JL, Waits LP, Ausband DE, Mack CM (2010) Efficient noninvasive genetic sampling for monitoring reintroduced wolves. J Wildl Manag 74:1050–1058
Theuerkauf J, Rouys S, Jedrzejewski W (2003) Selection of dens, rendezvous and resting sites by wolves in the Bialowieza Forest, Poland. Can J Zool 81:163–167
Thiel RP, Merrill S, Mech LD (1998) Tolerance by denning wolves, Canis lupus, to human disturbance. Can Field-Nat 122:340–342
Treves A, Martin KA, Wydeven AP, Wiedenhoeft JE (2011) Forecasting environmental hazards and the application of risk maps to predator attacks on livestock. BioScience 61:451–458
Unger D, Keenlance P, Kohn B, Anderson E (2009) Factors influencing homesite selection by grey wolves in northwestern Winsconsin and east-central Minnesota. In: von Wydeven AP, van Deelen TR, Heske E (eds) Recovery of gray wolves in the Great Lakes Region of the United States: an endangered species success story. Springer, New York, pp 175–189
Walton LR, Cluff HD, Paquet PC, Ramsey MA (2001) Movement patterns of barren ground wolves in central Canadian Arctic. J Mammal 82:867–876
Wam HK (2003) Wolf behavior towards people. MSc Thesis, Norwegian University of Life Sciences, Aas
Weaver JL, Paquet PC, Ruggiero LF (1996) Resilience and conservation of large carnivores in the Rocky Mountains. Conserv Biol 10:964–976
Acknowledgments
The authors would like to thank I. Hatzimichael, V. Koutis, S. Tzorgakis, P. Menounos, P. Pavlides, G. Giannatos, K. Selinides, Y. Lazarou, M. Petridou, A. Giannakopoulos, S. Riegler, H. Pilides, and T. Tragos for assisting during field work; I. Aravides, Y. Mertzanis, S. Psaroudas, and K. Papapavlou for their support throughout the study; and C. Astaras for linguistic support and helpful comments during revision of the manuscript. We also thank two anonymous referees whose comments helped to greatly improve an earlier version of the manuscript. Field work and analyses were partially funded in the framework of several conservation projects by CALLISTO NGO, E.C DG Env., Greek Ministry of Agriculture, Arcturos NGO, Egnatia odos S.A, Ergose, Argyropoulos SA, and Exergia S.A.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by C. Gortázar
Rights and permissions
About this article
Cite this article
Iliopoulos, Y., Youlatos, D. & Sgardelis, S. Wolf pack rendezvous site selection in Greece is mainly affected by anthropogenic landscape features. Eur J Wildl Res 60, 23–34 (2014). https://doi.org/10.1007/s10344-013-0746-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10344-013-0746-3