
Abstract
Over the last century, human activity has caused significant changes to the activity patterns of many wildlife species. The wild boar is one species known to change its activity pattern with the intensity of human disturbance. We conducted camera trap surveys in two study sites, Shingo and Himuro, in Tochigi, central Japan. We investigated effects of two types of human disturbance on the activity pattern of a wild boar population: ‘direct’ disturbance related to hunting activity and ‘indirect’ disturbance related to daily human activity. In the hunting season, relative abundance indices (RAI) of wild boars significantly decreased, and the proportion of activity at night increased compared with the nonhunting season. RAI of wild boars at night decreased with increasing distance from the settlement, while RAI of wild boars during the day did not. Relative proportion of activity at night was higher in cameras at 0–200 m from the settlements, while no significant pattern was found in cameras far from settlements. Both direct and indirect effects of human activity had a significant effect on the activity pattern of wild boars. A decrease in human activity may result in the rapid expansion of wild boar populations, and re-evaluation of the human factor is important for more intelligent management of wild boar populations and to solve the human–wildlife conflict.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Over the last century, human activity has caused significant changes to the activity patterns of many wildlife species. Some of these changes have been related to the expansion of human activity, such as overpopulation or urban development (Markovchick-Nicholls et al. 2008; Riley et al. 2003), and some have been related to the reduction of human activity, such as depopulation or agropastoral abandonment (Rippa et al. 2011; Sirami et al. 2008). The impacts of human activities are often complex, and influence many ecological and behavioural aspects of wild animals, such as spatial usage and activity patterns.
Wildlife populations living close to human settlements are likely to be exposed to many types of human disturbance. Thus, they might adopt activity patterns and behaviours according to the intensity of this disturbance. There are two types of human disturbance that have the potential to cause changes in the activity of wildlife. One is human disturbance related to hunting activities, which is accompanied by direct interference on animals, such as through chasing or shooting (‘direct’ disturbance). For a broad range of animal species, a considerable proportion of their mortality is affected by hunting activities (Nores et al. 2008; Theuerkauf and Rouys 2008). Hunting can shape various ecological aspects of wildlife, such as population dynamics (e.g. Tod’go et al. 2008) or genetic structure (e.g. Allendorf et al. 2008). In addition, the stress and fear caused by hunting activities may lead to modification of spatial usage (e.g. Grignolio et al. 2011) and behaviour (e.g. de Boer et al. 2004) of hunted populations. Animals can change habitat selection patterns (Tolon et al. 2009, 2012; Sad’d et al. 2012), reallocate time budgets and switch activity patterns (Lima and Bednekoff 1999), and modulate vigilance levels (Sodeikat and Pohlmeyer 2007) in response to hunting activities. Because hunting is restricted in specific areas, seasons and times of day, it usually generates significant variation of risk across space and time.
On the other hand, disturbance resulting from daily human activities related to agriculture, transport or construction might also have the potential to cause considerable effects on wildlife. Many studies have observed behavioural responses (Li et al. 2011; Sibbald et al. 2011) following encounters with humans. These human disturbances, which do not directly interfere with animals (‘indirect’ disturbance), have been linked to reduced reproductive success (Phillips and Alldredge 2000) and may induce long-term population decline (French et al. 2011). However, there has been little information for cryptic species, possibly due to the fact that their behaviour makes research through direct observation difficult.
In recent years, camera traps have become important tools for research on wildlife activity, especially for cryptic species (Carbone et al. 2001; Di Bietti et al. 2006; O’Brien et al. 2003; Vine et al. 2009). Camera-based activity assessments likely offer a better approximation of population level activity patterns than individual-level, telemetry-based methods (Bridges et al. 2004). Many camera trap studies have targeted rare and endangered species, whose conservation is threatened by human disturbance (Dinata et al. 2008; Karanth 1995; Kinnaird et al. 2003). However, camera traps also have the potential to become an important tool for research into common and expanding species, which are related to human–wildlife conflicts such as damage to agriculture.
In Japan, the distribution of wild boars, Sus scrofa, has drastically expanded in the last decade, and agricultural damage ensuing from this expansion is becoming a serious problem (Kuwabara et al. 2010; Nomoto et al. 2010; Ohtsuka-Ito and Kanzaki 1998). Total economic loss caused by wild boars was 5,400,000,000 yen in 2008, mainly as a result of damage to rice, wheat, maize and potatoes. Agricultural damage from wild boars has been especially serious in rural areas with mixed forest and fields, which represents Japan’s cultural landscape (satoyama, as discussed by Nakagoshi and Hong 2001). In the satoyama landscape, changes in social and economic conditions have resulted in the growth of abandoned fields, orchards and bamboo groves, which has in turn created suitable habitat for wild boars near settlements and agricultural fields (Kodera et al. 2001). However, the agricultural damage inflicted by wild boars living in close proximity to the satoyama is problematic because agricultural activity in these areas has an important role in maintaining high biodiversity (Katoh et al. 2009). Thus, solving the human–wildlife conflict in satoyama areas remains a practical issue for farmers and wildlife managers, but is also important for the ecosystem management of these culturally valuable landscapes.
There are many studies that have investigated ways of developing effective fences to prevent agricultural damage from wild boars (e.g. Takeuchi and Eguchi 2007). However, several recent studies have emphasized the importance of considering the role played by humans in determining the efficacy of these fences (Cai et al. 2008; Honda 2005; Saito et al. 2011). The wild boar is a species known to change its activity pattern depending on the intensity of human activity, and such behavioural flexibility might influence the result of efforts aimed at preventing their damage on agriculture. When existing in a natural, undisturbed state, wild boars are regarded as being generally diurnal animals (Kurz and Marchinton 1972). However, they supposedly shift to nocturnal behaviours when faced with hunting pressure (Boitani et al. 1994; Singer et al. 1981). Additionally, there are several studies that report a possible effect of indirect human disturbance on the activity pattern of wild boars. Singer et al. (1981) reported the reaction of radio-collared animals to being closely approached by an observer, and detected a significant decrease in daily movement after disturbance, compared with the amount of movement with no disturbance. Based on their GIS-based habitat suitability model constructed from a field sign survey, Park and Lee (2003) also reported that wild boars will avoid hiking trails. Although the importance of direct and indirect human disturbance on the activity pattern of wild boars has been recognized for a long time, there is considerable difficulty in directly observing wild boars because of their cautious behaviour and preference for habitats comprising dense thickets (Kodera et al. 2001). Camera traps represent an ideal solution to this problem, making it possible to study the activity of wild boars and how this is influenced by human activity.
Our aim for this study was to examine the effect of humans on the activity pattern of wild boars, using camera traps to capture data, and with a special focus on elucidating the difference between direct and indirect human disturbance. As wild boars should display their natural behaviour when less disturbed, diurnal activity should increase when wild boars are exposed only to minor, short-term disturbance. In Japan, gun hunting is strictly regulated and permitted only in the winter hunting season during daylight hours. A nuisance control program for wild boars is conducted by trapping and is carried out year-round. While gun hunting causes stress and fear for the target animal, trapping tries to lure boars into enclosures using baits and attractants. From the perspective of activity patterns of wild boars, seasons with both gun hunting and trapping may result in more intense disturbance than seasons with only trapping. Therefore, activity patterns of wild boars may shift from diurnal in the nonhunting season towards nocturnal in the hunting season, according to the temporal variation of intense direct disturbance. Conversely, the intensity of daily human activity, which causes indirect disturbance, could be regarded as a stable disturbance in space with a constant temporal pattern. Since the intensity of disturbance will likely be highest inside settlements, any effect of indirect human disturbance tends to occur on daytime, and should be greatest near the settlement and decrease with distance. If there are any effects of indirect human disturbance, activity patterns of wild boars should shift from being more nocturnal near the settlement to more diurnal further away. Furthermore, because activity patterns of wild boars may be influenced by other environmental factors or human activities within settlements, it is important to compare these results between multiple settlements with different environments.
First, to examine the effect of direct human disturbance on wild boars, we compare their activity patterns during the hunting and nonhunting seasons. Second, we examine whether the activity patterns of wild boars show any difference according to distance from human settlements, and if there is a difference, we determine the distance away from the settlement at which the greatest behavioural difference is found. Finally, we discuss the effect of direct and indirect human disturbance on the activity pattern of wild boars in satoyama regions, and how this plays into conflicts on the territorial boundaries between humans and wildlife.
Material and methods
Study area
We carried out this study at two sites within the southwestern part of Tochigi prefecture, central Japan. The first study site, Shingo [36°24′ N, 139°32′ E, 58–416 m above sea level (a.s.l.), 46.7 km2] is located on the boundary of the cities of Ashikaga and Sano in Tochigi prefecture. The landscape of this area is composed of agricultural land use and forest with complex shapes. The forest is mainly deciduous coppice forest dominated by Konara oak Quercus serrata and Japanese chestnut Castanea crenata. Human population density of this area is 99.4–755.1 people/km2 (Ministry of Internal Affairs and Communications 2010). Percentage of area covered by forest is 27.2–79.0 %, and percentage of abandoned field within the agricultural land is 31.0–49.0 % (Ministry of Agriculture, Forestry and Fisheries 2010).
The second study site, Himuro [36°30′ N, 139°32′ E, 191–701 m a.s.l., 12.4 km2], is located at the northern part of Sano. The landscape of this area is mainly composed of forest with steep slopes, and narrow-shaped settlements are distributed along the deep valley. The forest is dominated by an evergreen coniferous plantation of Japanese cypress Cryptomeria japonica and Japanese cedar Chamaecyparis obtusa, mostly planted in 1960–1970. A small patch of deciduous coppice forest remains only on a steep slope. Human population density of this area is 21.9 people/km2 (Ministry of Internal Affairs and Communications 2010). The percentage of area covered by forest is 92.9 %, and percentage of abandoned field within the agricultural land is 61.3 % (Ministry of Agriculture, Forestry and Fisheries 2010).
The climate is temperate with an average temperature of 13.9 °C, varying from 2.8 °C in January to 25.7 °C in August. Annual precipitation averages 1,244.7 mm, with maximum rainfall in summer and minimum in winter (Sano weather station, 36°20.1′ N, 139°33.6′ E, 39 m a.s.l., 1981–2010; http://www.jma.go.jp/jma/index.html. Accessed 11 Feb 2012).
In Japan, the hunting season begins on 15 November, and finishes on 15 February the following year. Tochigi Prefectural governance has extended the hunting season until 28 February for the purpose of nuisance control. Hunters must obtain permission at the prefectural office to hunt during the hunting season. Gun hunting is allowed only during daylight hours, and is not permitted within a 300-m radius of human settlements and areas with special protection status. There is no restriction on the number of hunters on the hunting ground, and both group and solitary hunting are allowed. Gun hunters tend to use hunting dogs to chase targets in this landscape with steep and complex topography. Box and snap traps are common trapping methods; usually, box traps use baits and snap traps are set on the animal trail without baits. Since trapping requires frequent patrol and maintenance, total numbers of traps allowed for a single hunter is restricted to 30 at maximum. The main game species for hunting in our study area are sika deer (Cervus nippon) and wild boar. Local hunters with special permission have conducted a nuisance control program for wild boars in both sites all year. Usually, trapping is preferred for the nuisance control program, and gun hunting is scarcely conducted in the nonhunting season. Between April 2010 and March 2011, 540 wild boars were trapped and 39 shot by gun hunting in Shingo, and 56 were trapped and 6 shot by gun hunting in Himuro (Tochigi prefectural government, unpublished).
Camera trap survey
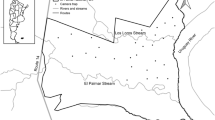
The camera trap survey was conducted from July 28, 2010 to July 28, 2011. We set up 51 camera traps at Shingo and 32 camera traps at Himuro (Fig. 1). Cameras were set in the forest (Shingo: n = 48; Himuro: n = 28) or abandoned fields with woody plants (Shingo: n = 3; Himuro: n = 4) at varying distances from the settlement. Cameras near the settlement were adjacent to various environment types, including bamboo groves, orchards, abandoned fields or cultivated fields. We calculated the distance from the settlement for each camera using the ‘Basic Forest Map in Tochigi Prefecture’ provided by the Tochigi prefectural government (unpublished) by using ArcGIS 10.0 (Esri Inc., USA). Wild boars usually move on the ground and their body size changes drastically during maturation. To capture the whole body of an adult boar in one image, we fixed cameras at a height of 180 cm, with an angle of 30° declination, on trees near animal trails. Our camera traps were infrared motion-detecting cameras (Capture Cuddeback Inc., USA), which are capable of responding to the movement of animals larger than rodents with a trigger speed of 1/3 s. The minimum time delay between two successive images was set to 1 min, and operated 24 h/day or until the batteries ran out. Because some camera traps were set in locations far from the settlement and without motorways, it was time-consuming to check all cameras during the sampling period to monitor performance or change batteries. Cameras were left in the forest for 30–40 days, and the number of trap days was calculated for each census. We identified each image of an animal to species level, recorded the time and date, and rated each image as a dependent or independent event. We defined consecutive images of the same species within 30 min as a dependent event and others as an independent event. We calculated the number of images captured per camera per day [relative abundance indices (RAI); following O’Brien et al. (2003)] as an index for relative frequency of activity. We calculated RAI in the daytime (RAIDay) and RAI in the nighttime (RAINight) separately, to evaluate the effect of human activity on wild boar activity by difference in diurnal and nocturnal activity. We defined day and night according to times of sunrise and sunset in Utsunomiya [36°34.0′ N, 139°53.0′ E]. We checked capture time and date for each image, and compared the daily sunrise and sunset times, which were provided by the National Astronomical Observatory of Japan (2011). We calculated the proportion of day and night in each day to correct the bias resulting from differences in the proportion of day and night among seasons. We counted number of images during the day (or night) and divided this by trapping effort, which was calculated from the number of trap days multiplied by the proportion of day (or night).

Location of study area, and location of camera traps at Shingo and Himuro, Tochigi prefecture, central Japan. Gray area in map indicates the settlements, and white area in the map indicates the forests. Filled circles represent location of camera traps
Analysis
We fitted generalized linear mixed models (GLMMs) to test the effect of time, seasons, distance from settlement and difference between two survey sites on the RAI of wild boars. The response variable was the number of wild boar images, and trapping effort was specified as offset, and included in the linear predictor during model fitting. Poisson distribution was specified as error distribution, and logarithm was specified as a link function. Three categorical variables with two states, one continuous variable and all possible interactions were considered as fixed effect: time (night or day), season (hunting or nonhunting), region (Shingo or Himuro) and the distance from the settlement (continuous variable). The region variable determines the possible effect of regional differences between survey sites, such as wild boar density or spatial usage pattern. To exclude possible inherent effects of other factors, such as spatial arrangement or environmental variability for each camera location, camera identity was specified as a random factor in our model. Stepwise backward model selection by AIC was performed to select the best subset of predictor variable.
To examine the difference in ratio of RAIDay and RAINight of wild boars between the nonhunting and hunting seasons (‘direct’ effect of human disturbance) and the distance from the settlement (‘indirect’ effect of human disturbance), we performed a randomization test. If the ‘direct’ or ‘indirect’ effect of human disturbance causes changes in activity patterns of wild boars, significant bias on RAIDay and RAINight should be observed. Therefore, we examined whether the observed pattern on the ratio of RAIDay and RAINight differs from the null distribution obtained from a randomization procedure. At first, we obtained observed patterns on the ratio of RAIDay and RAINight as follows. We divided the cameras into three groups according to the distance from the settlement: 0–200, 200–400 and >400 m. In each group and season, we counted the number of cameras that showed higher RAIDay than RAINight (RAIDay > RAINight). To obtain the null distribution, we randomly shuffled combinations of RAIDay and RAINight. For each trial, we counted the number of pairs with RAIDay > RAINight. We repeated the trial 100,000 times and until a 95 % CI of number of pairs with higher RAIDay than RAINight was reached. We checked whether observed values were included within the range of 95 % CI.
If significant bias on the ratio of RAIDay and RAINight was observed at a certain distance from the settlement (200 or 400 m), we used this distance as a threshold to divide the camera into two groups: ‘the camera near the settlement’ and ‘the camera far from the settlement’. We examined the relationship between active times of wild boars and humans. We classified the images by 3-h intervals, and calculated the Spearman’s rank correlation coefficient between RAI in wild boars and humans, in each camera group in two survey sites, and in hunting season and nonhunting season. All statistical analyses were performed using R 2.13.0 (R Development Core Team 2011).
Results
In the study period, a total of nine censuses were conducted; six censuses were conducted in the nonhunting season (July 28 to September 3, September 3 to October 14, October 14 to November 18, 2010; April 16 to May 19, May 19 to June 19, June 19 to July 28, 2011) and three censuses overlapped with the hunting season (November 18 to December 26, 2010; December 30, 2010 to February 3, 2011; February 3 to March 21, 2011). RAI of wild boars showed no significant difference between Shingo and Himuro in both nonhunting and hunting seasons (Mann–Whitney’s U test, P > 0.05, Table 1). RAI of humans was greater in Shingo than in Himuro in the nonhunting season (Mann–Whitney’s U test, P < 0.01, Table 1), and there was no significant difference in the hunting season (Mann–Whitney’s U test, P > 0.05, Table 1). Images of humans included hikers, farmers or local residents as well as hunters.
The results of mixed models revealed that the RAI of wild boars increased at night (time: P < 0.001, Table 2), and decreased in the hunting season (hunting: P < 0.001, Table 2). Moreover, RAI of wild boars tended to be higher at night during in the hunting season, as indicated by the significant interaction term between season and time (hunting × time: P < 0.001, Table 2). The effect of distance from settlement was significant during the night and tended to increase in the cameras closer to the settlement (distance × time: P < 0.001, Table 2). In addition, the effect of distance from settlement was significant in the hunting season and tended to decrease in the cameras closer to the settlement (distance × hunting: P < 0.05, Table 2). In Himuro, RAI of wild boars significantly declined in the hunting season (region × hunting: P < 0.001, Table 2). The effect of distance from the settlement at night was more significant in Shingo compared with in Himuro (distance × region × time: P < 0.001, Table 2).
In the nonhunting season, observed number of cameras with RAIDay > RAINight was lower than 95 % CI of null distribution obtained from 100,000 randomization trials in the cameras at 0–200 m from the settlement (Fig. 2). On the other hand, observed number of cameras with RAIDay > RAINight was within the range of 95 % CI of null distribution in the other groups of cameras. In the hunting season, observed number of cameras with RAIDay > RAINight was within the range of 95 % CI of null distribution in all groups of cameras (Fig. 2).

Difference in proportion of diurnal and nocturnal activity of wild boar according to hunting activity and distance from settlement. Vertical line with closed triangle shows observed number of cameras with RAIDay > RAINight. Histogram shows number of pairs with RAIDay > RAINight obtained from 100,000 randomization trials. Gray zone indicates 95 % confidence interval obtained from randomization procedure. a Nonhunting season. b Hunting season. Left 0–200 m from the settlement, middle 200–400 m from the settlement, right >400 m from the settlement
In the cameras within 200 m from the settlement, wild boars showed clear nocturnal activity in both the nonhunting and hunting season in two regions (Fig. 3). In the cameras further than 200 m from the settlement in Shingo, wild boars showed higher activity during the day in the nonhunting season. On the other hand, no clear pattern was observed in the hunting season in Shingo and in both seasons in Himuro (Fig. 3).

The activity pattern of wild boars (filled circles) and humans (empty squares) in cameras located <200 m from the settlement (left) and >200 m from the settlement (right) in Shingo (a) and Himuro (b). Upper activity pattern in nonhunting season; lower activity pattern in hunting season
Activity of humans was mainly concentrated between 0900–1500 hours throughout the year, in all camera locations. Active times of wild boars and humans had significant negative correlation in the cameras within 200 m from the settlement. However, no significant correlation was found in the cameras further than 200 m from the settlement (Fig. 3).
Discussion
Effect of hunting on the activity pattern of wild boars
The wild boar is a species that shows remarkable flexibility in its ecological and behavioural traits. When undisturbed, wild boars tend to be more or less diurnal (Kurz and Marchinton 1972), and this is assumed to be their natural behaviour. However, wild boars are also known to become nocturnal under hunting pressure (Boitani et al. 1994; Russo et al. 1997), the impact of which also affects the activity patterns of a number of other wildlife species (Grignolio et al. 2011). For the wild boar, hunting activity causes changes in nocturnal activity (Sodeikat and Pohlmeyer 2007), instability of spatial usage and increase in resting range (Maillard and Fournier 1995; Tolon et al. 2009; Scillitani et al. 2010). Heavy hunting pressure can sometimes cause wild boars to abandon parts of their native range (Scillitani et al. 2010). However, Keuling et al. (2008a, b) pointed out the importance of factors other than hunting, such as seasonal change in availability of food and shelter, in impacting wild boar activity patters. The study of Keuling (2008b) demonstrated lower nocturnal activity during the hunting season in study areas with nocturnal hunting. However, their results did not exclude the effect of weather condition, whereby wild boars decreased their activity to save energy during cold winter nights when frozen soil prevented rooting (Massei et al. 1997; Keuling 2008b). In this study, results of generalized mixture models indicated that, in both study sites, the RAI of wild boars in the hunting season showed a significant decrease in activity overall and a higher proportion of nocturnal behaviour, compared with the nonhunting season (Table 2). Because trapping was conducted all year round in our study area, the difference in human disturbance regimes between the nonhunting and hunting seasons resulted from the presence of gun hunting. In contrast to the result of Keuling (2008b), our results revealed a significant decrease in diurnal activity in the study area only with daytime hunting. Although several studies have documented the seasonal change of activity patterns of wild boars resulting from thermoregulation (Kurz and Marchinton 1972; Singer et al. 1981), food supply and weather condition (Keuling et al. 2008a), these findings are not applicable to our results, because significantly increasing nocturnal activity in the cooler season would require increased energy and in turn significantly reduce stored fat deposits. Therefore, our results suggest that gun hunting was a major factor determining the activity pattern of wild boars, as the animals were changing their activity pattern to avoid hunters and their hounds.
The number of wild boars captured by gun shooting was ca. 10 times smaller than the number captured by trapping in both study sites (Tochigi prefectural government, unpublished). In addition to the animals captured, the direct disturbance of hunting behaviour, such as being chased by hunters or hounds, might have a large impact on the activity pattern of wild boars. Previous studies have shown that hunting can significantly reduce the population density of wild boars (Acevedo et al. 2006). However, the effect of actual harvest number on wild boar population dynamics might be limited, when compared with factors such as climatic condition and food supply (Okarma et al. 1995). Geisser and Reyer (2004) have shown the importance of hunting in reducing the frequency of damage inflicted by wild boars, rather than the effect of actual harvest. Sakata et al. (2008) also reported that sighting per unit effort of wild boars decreased according to the gun hunting effort. The results of our study suggested that disturbance from hunting activity may play an important role on the activity pattern of wild boars. However, unregulated hunting may cause undesired effects, including wild boars seeking refuge in valuable areas, such as densely populated areas or golf courses, in which gun hunting is restricted.
Because we divided season by presence of hunting activity, the biological interpretation for seasonal differences of wild boar activity was restricted in our analysis. For intelligent management of wild boar populations, more study is required in areas such as analysis of individual spatial usage, changes in home range size or degree of escaping behaviour, as well as changes in temporal activity patterns related to behaviour, such as rutting season or denning period.
Effect of distance from settlement on the activity pattern of wild boars
In both study sites, RAI of wild boars at night increased with increasing proximity to settlements. However, distance from settlements did not influence RAI during the day (Table 2). Because camera traps are triggered by the movement of animals, higher RAI intrinsically indicate a higher activity level of the target species. Thus, our results indicated that high activity areas of wild boars were concentrated near settlements. Wild boars are known to use the main part of their active time for foraging (Cahill et al. 2003), and we suggest that the wild boar population in the study area might be depending on human-related food resources, such as bamboo shoots, fruit and possibly crops, that are distributed around the settlement. The level of dependency of wild boars on crops is likely to differ between individuals. Honda et al. (2008) confirmed that individuals using areas outside a forest, and that were assumed to have high dependency on crops, also intensively used the area within 200 m from the forest edge. We suspect that the wild boar population in our study area is mainly composed of individuals with high dependency on the environment around the settlement. Thus, for wild boar management in zones similar to our study area, we recommend, in addition to animal trapping or shooting, that efforts be made to change the habitat quality around the settlement by removing food resources, such as abandoned fruits or bamboo shoots.
In our study, wild boars displayed nocturnal activity in the area 0–200 m from the settlement and showed more random patterns in the cameras far from the settlement in the nonhunting season (Fig. 2). Although activity pattern was not significantly biased from random chance in the hunting season (Fig. 2), wild boars significantly avoided activity during the times when humans were active when they were within 200 m from the settlement (Fig. 3), but not in the cameras further than 200 m from the settlement (Fig. 3) in both nonhunting and hunting seasons. The home range size of wild boars has an approximate radius of 500–600 m when converted into a circle (Kodera et al. 2010), thus a distance of 200 m is not sufficiently great to prevent the movement of wild boars. Despite the small distance, activity time of wild boars did differ according to the distance from the settlement at the threshold of 200 m (Fig. 2). Keuling (2008b) pointed out that the activity of wild boars during the day increased at 150 m away from artificial structures such as a road or building. Ueda and Jiang (2004) compared the active time of wild boars in functioning and abandoned orchards, and revealed that wild boars appear earlier at abandoned orchards with few humans. Wild boars in this study seemed to change their activity patterns at small scales and avoid habitats close to the settlements at times at which the risk of encountering humans is high.
In our study, the difference in active time in relation to distance from the settlement was smaller in Himuro compared with in Shingo (Table 2). This could be because Himuro is located further from the central part of the city. In Japan, depopulation and aging of residents are increasing more rapidly in remote areas with lower economic activity and less access to basic services, education and medical assistance. Therefore, pressure from human activity associated with transportation and daily life, which indirectly affects wild boar activity, might be lower in Himuro compared with in Shingo, as indicated by the lower RAI of humans in the nonhunting season (Table 1). Our results might reflect the reduced human activity in the Himuro settlement that in turn is causing wild boars to use the habitat around the settlement more during the day and to display similar activity patterns as they do in the forest. Further, the intensity of management pressure on agricultural land has been inevitably weakened in Himuro as a result of depopulation and aging. Expansion of abandoned fields can create suitable habitat for wild boars very near settlements, sometimes even within a few metres from the houses of local residents. This proximity of suitable wild boar habitat to human residential areas may enhance wild boar habituation to human activity. In Himuro, we found wild boars rooting all over the settlement and even found a wild boar resting bed in the garden of a vacant house within a human residential area (Ohashi et al., personal observation). However, our analysis could not separate the concurrent effects of human activity and environmental condition. Further analysis combined with environmental factors, in addition to direct and indirect human disturbance, is required to investigate the impact of human activity and environmental condition created by human management regimes on the activity pattern of wild boars.
Conclusion
In this study, we conducted camera trap surveys in the Tochigi prefecture, central Japan, to clarify the effect of human activity on activity patterns of wild boars. Activity patterns of wild boars were different between the nonhunting and hunting seasons, with wild boars showing more nocturnal activity in the hunting season. Additionally, wild boars avoided high activity during the time of day when humans were active when they were within 200 m from settlements, but not 200 m away from settlements. Our study revealed the high behavioural flexibility of wild boars to the many types of human activities present in satoyama areas, which represent a landscape in which the territories of humans and wildlife meet. Decline of human activity in the satoyama area might result in the expansion of more favourable conditions for wild boar populations, especially when combined with changes in habitat condition around the settlement. Further study is required to understand the effect of human activity on the activity pattern of wild boars, and to advance more intelligent methods for damage control and population management.
References
Acevedo P, Escudero MA, Muńoz R, Gortázar C (2006) Factors affecting wild boar abundance across an environmental gradient in Spain. Acta Theriol 51:327–336
Allendorf FW, England PR, Luikart G, Ritchie PA, Ryman N (2008) Genetic effects of harvest on wild animal populations. Trends Ecol Evol 23:327–337
Boitani L, Mattei L, Nonis D, Corst F (1994) Spatial and activity patterns of wild boars in Tuscany, Italy. J Mammal 75:600–612
Bridges AS, Vaughan MR, Klenzendorf S (2004) Seasonal variation in American black bear Ursus americanus activity patterns: quantification via remote photography. Wildl Biol 10:277–284
Cahill S, Llimona F, Gràcia J (2003) Spacing and nocturnal activity of wild boar Sus scrofa in a Mediterranean metropolitan park. Wildl Biol 9(Suppl):3–13
Cai J, Jiang ZG, Zeng Y, Li CW, Bravery BD (2008) Factors affecting crop damage by wild boar and methods of mitigation in a giant panda reserve. Eur J Wildl Res 54:723–728
Carbone C, Christie S, Conforti K, Coulson T, Franklin N, Ginsberg JR, Griffiths M, Holden J, Kawanishi K, Kinnaird M, Laidlaw R, Lynam A, Macdonald DW, Martyr D, McDougal C, Nath L, O’Brien T, Seidensticker J, Smith DJL, Sunquist M, Tilson R, Wan Shahruddin WN (2001) The use of photographic rates to estimate densities of tigers and other cryptic mammals. Anim Conserv 4:75–79
de Boer HY, van Breukelen L, Hootsmans MJM, van Wieren SE (2004) Flight distance in roe deer Capreolus capreolus and fallow deer Dama dama as related to hunting and other factors. Wildl Biol 10:35–41
Di Bitetti MS, Paviolo A, De Angelo C (2006) Density, habitat use and activity patterns of ocelots (Leopardus pardalis) in the Atlantic Forest of Misiones, Argentina. J Zool 270:153–163
Dinata Y, Nugroho A, Haidir IA, Linkie M (2008) Camera trapping rare and threatened avifauna in west-central Sumatra. Bird Conserv Int 18:30–37
French SS, González-Suárez M, Young JK, Durham S, Gerber LR (2011) Human disturbance influences reproductive success and growth rate in California sea lions (Zalophus californianus). PLoS One 6:1–8
Geisser H, Reyer H (2004) Efficacy of hunting, feeding, and fencing to reduce crop damage by wild boars. J Wildl Manag 68:939–946
Grignolio S, Merli E, Bongi P, Ciuti S, Apollonio M (2011) Effects of hunting with hounds on a non-target species living on the edge of a protected area. Biol Conserv 144:641–649
Honda T (2005) Evaluation of simple fences in preventing wild boar (Sus scrofa) damage to agricultural crops: verification from factor “installation and maintenance”. Wildl Conserv Jpn 9:93–102, in Japanese with English abstract
Honda T, Hayashi Y, Sato Y (2008) Habitat selection of wild boar (Sus scrofa) captured near the forest edge. Mamm Sci 48:11–16, in Japanese with English abstracts
Karanth K (1995) Estimating tiger Panthera tiglis populations from camera-trap data using capture-recapture models. Biol Conserv 71:333–338
Katoh K, Sakai S, Takahashi T (2009) Factors maintaining species diversity in satoyama, a traditional agricultural landscape of Japan. Biol Conserv 142:1930–1936
Keuling O, Stier N, Roth M (2008a) Annual and seasonal space use of different age classes of female wild boar Sus scrofa L. Eur J Wildl Res 54:403–412
Keuling O, Stier N, Roth M (2008b) How does hunting influence activity and spatial usage in wild boar Sus scrofa L.? Eur J Wildl Res 54:729–737
Kinnaird MF, Sanderson EW, O’Brien TG, Wibisono HT, Woolmer G (2003) Deforestation trends in a tropical landscape and implication for endangered large mammals. Conserv Biol 17:245–257
Kodera Y, Kanzaki N, Kaneko Y, Tokida K (2001) Habitat selection of Japanese wild boar in Iwami district, Shimane Prefecture, western Japan. Wildl Conserv Jpn 6:119–129, in Japanese with English abstract
Kodera Y, Nagatsuma T, Sawada S, Fujihara S, Kanamori H (2010) How does spreading maize on fields influence behavior of wild boars (Sus scrofa)? Mamm Sci 50:137–144, in Japanese with English abstract
Kurz JC, Marchinton RL (1972) Radiotelemetry studies of feral hogs in South Carolina. J Wildl Manag 36:1240–1248
Kuwabara T, Ohashi H, Saito M, Hiroshige Y, Koike S, Toda H, Kaji K (2010) Conditions and course of solution on rural agricultural damages caused by wild boar. J Rural Econ Special Issue 2010:305–312, in Japanese
Li C, Monclús R, Maul TL, Jiang Z, Blumstein DT (2011) Quantifying human disturbance on antipredator behavior and flush initiation distance in yellow-bellied marmots. Appl Anim Behav Sci 129:146–152
Lima SL, Bednekoff PA (1999) Temporal variation in danger drives antipredator behavior: the predation risk allocation hypothesis. Am Nat 153:649–659
Maillard D, Fournier P (1995) Effects of shooting with hounds on size of resting range of wild boar (Sus scrofa L.) groups in Mediterranean habitat. IBEX J M E 3:102–107
Markovchick-Nicholls L, Regan HM, Deutschman DH, Widyanata A, Martin B, Noreke L, Hunt TA (2008) Relationships between human disturbance and wildlife land use in urban habitat fragments. Conserv Biol 22:99–109
Massei G, Genov PV, Staines BW, Gorman ML (1997) Factors influencing home range and activity of wild boar (Sus scrofa) in a Mediterranean coastal area. J Zool 242:411–423
Ministry of Agriculture, Forestry and Fisheries (2010) Report of Tochigi Prefecture: census of agriculture and forestry 2010 (in Japanese). http://www.maff.go.jp/j/tokei/census/afc/2010/dai1kan.html. Accessed 11 Feb 2012
Ministry of Internal Affairs and Communications (2010) Population census of the Japan in 2010. http://www.stat.go.jp/English/index.htm. Accessed 11 Feb 2012
Nakagoshi N, Hong S-K (2001) Vegetation and landscape ecology of East Asian ‘Satoyama’. Glob Environ Res 5:171–181
National Astronomical Observatory of Japan (2011) Koyomi Station. http://eco.mtk.nao.ac.jp/koyomi/index.html.en. Accessed 1 Oct 2011
Nomoto K, Takahashi T, Koganezawa M, Fukumura K (2010) Relationships between crop damage by wild boar and surrounding environmental characteristics in paddy fields and non-paddy fields in Motegi, Japan. Mamm Sci 50:129–135, in Japanese with English abstract
Nores C, Llaneza L, Álvarez MÁ (2008) Wild boar Sus scrofa mortality by hunting and wolf Canis lupus predation: an example in northern Spain. Wildl Biol 14:44–51
O’ Brien TG, Kinnaird MF, Wibisono HT (2003) Crouching tigers, hidden prey: Sumatran tiger and prey populations in a tropical forest landscape. Anim Conserv 6:131–139
Ohtsuka-Ito E, Kanzaki N (1998) Population trends of the Japanese wild boar during the Showa era. Wildl Conserv Jpn 3:95–105, in Japanese with English abstract
Okarma H, Jędrzejewska B, Jędrzejewski W, Kransiński, Miłkowski L (1995) The roles of predation, snow cover, acorn crop, and man-related factors on ungulate mortality in Białowieża Primeval Forest, Poland. Acta Theriol 40:197–217
Park C-R, Lee W-S (2003) Development of a GIS-based habitat suitability model for wild boar Sus scrofa in the Mt. Baekwoonsan region, Korea. Mamm Stud 28:17–21
Phillips GE, Alldredge AW (2000) Reproductive success of elk following disturbance by humans during calving season. J Wildl Manag 64:521–530
R Development Core Team (2011) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Riley SP, Sauvajot MS, Fuller TK, York EC, Kamradt DA, Bromley C, Wayne RK (2003) Effects of urbanization and habitat fragmentation on bobcats and coyotes in southern California. Conserv Biol 17:566–576
Rippa D, Maselli V, Soppelsa O, Fulgione D (2011) The impact of agro-pastoral abandonment on the rock partridge Alectoris graeca in the Apennines. Ibis 153:721–734
Russo L, Massei G, Genov PV (1997) Daily home range and activity of wild boar in a Mediterranean area free from hunting. Ethol Ecol Evol 9:287–294
Sad’d S, Tolon V, Brandt S, Baubet E (2012) Sex effect on habitat selection in response to hunting disturbance: the study of wild boar. Eur J Wildl Res 58:107–115
Saito M, Momose H, Mihira T (2011) Both environmental factors and countermeasures affect wild boars damage to rice paddies in Boso Peninsula, Japan. Crop Prot 30:1048–1054
Sakata H, Samejima H, Yokoyama M (2008) Present status of a wild boar (Sus scrofa) population estimated by sighting per unit effort (SPUE), and relationships to snowfall, vegetation, hunting effort, agricultural damages and the SPUE of Sika deer (Cervus nippon). Mamm Sci 48:245–253, in Japanese with English abstracts
Scillitani L, Monaco A, Toso S (2010) Do intensive drive hunts affect wild boar (Sus scrofa) spatial behaviour in Italy? Some evidences and management implications. Eur J Wildl Res 56:307–318
Sibbald AM, Hooper RJ, McLeod JE, Gordon IJ (2011) Responses of red deer (Cervus elaphus) to regular disturbance by hill walkers. Eur J Wildl Res 57:817–825
Singer FJ, Otto DK, Tipton AR, Hable CP (1981) Home ranges, movements, and habitat use of European wild boar in Tennessee. J Wildl Manag 45:343–353
Sirami C, Brotons L, Burfield I, Fonderflick J, Martin J-L (2008) Is land abandonment having an impact on biodiversity? A meta-analytical approach to bird distribution changes in the north-western Mediterranean. Biol Conserv 141:450–459
Sodeikat G, Pohlmeyer K (2007) Impact of drive hunts on daytime resting site areas of wild boar family groups (Sus scrofa L.). Wildl Biol Pract 3:28–38
Takeuchi M, Eguchi Y (2007) The wild boar blocker fence. Res J Food Agric 30:15–18, in Japanese with English abstract
Theuerkauf PJ, Rouys S (2008) Habitat selection by ungulates in relation to predation risk by wolves and humans in the Białowieza Forest. For Ecol Manag 256:1325–1332
Tod’go C, Servanty S, Gaillard J-M, Brandt S, Baubet E (2008) Disentangling natural from hunting mortality in an intensively hunted wild boar population. J Wildl Manag 72:1532–1539
Tolon V, Dray S, Loison A, Zeileis A, Fischer C, Baubet E (2009) Responding to spatial and temporal variations in predation risk: space use of a game species in a changing landscape of fear. Can J Zool 87:1129–1137
Tolon V, Martin J, Dray S, Loison A, Fischer C, Baubet E (2012) Predator–prey spatial game as a tool to understand the effects of protected areas on harvester-wildlife interactions. Ecol Appl 22:648–657
Ueda H, Jiang Z (2004) The use of orchards and abandoned orchard by wild boars in Yamanashi. Mamm Sci 44:25–33
Vine SJ, Crowther MS, Lapidge SJ, Dickman CR, Mooney N, Piggott MP (2009) Comparison of methods to detect rare and cryptic species: a case study using the red fox (Vulpes vulpes). Wildl Res 36:436–446
Acknowledgments
This study was supported by the “Development of wildlife management system with DPSIR scheme” project, the special budgets for research and education in the fiscal year of 2010–2011 from the Ministry of Education, Culture, Sports, Science and Technology of Japan. We would like to thank to the prefectural office of Tochigi, and the city offices of Sano and Ashikaga for facilitating our survey. We would like to thank the local residents in the cities of Sano and Ashikaga for allowing the establishment of the camera trap stations around the settlements. We would like to thank Dr. Munemitsu Akasaka, Dr. Masato Yoshikawa, the students of the Laboratory of Vegetation Management and the students of the Laboratory of Wildlife Conservation in Tokyo University of Agriculture and Technology for the assistance in field surveys and helpful comments on an early draft of the manuscript. We would like to thank the editor and two anonymous referees for useful comments in revising the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by C. Gortázar
An erratum to this article is available at http://dx.doi.org/10.1007/s10344-014-0819-y.
Rights and permissions
About this article
Cite this article
Ohashi, H., Saito, M., Horie, R. et al. Differences in the activity pattern of the wild boar Sus scrofa related to human disturbance. Eur J Wildl Res 59, 167–177 (2013). https://doi.org/10.1007/s10344-012-0661-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10344-012-0661-z