
Abstract
Monitoring change in the population size of mouflon (Ovis gmelini musimon × Ovis sp.) in rugged areas is an important yet difficult task for wildlife ecologists. To assess population change of mouflon inhabiting the Caroux-Espinouse massif, France, we compared a pedestrian and helicopter survey, using counts of animals as indices of abundance. Environmental factors such as date of survey and temperature affected the detection of mouflons from the ground and the air. Both indices were sensitive to observed changes in population size. A decrease in the pedestrian index was recorded in 1994, the year following an epizootic of keratoconjunctivitis, which markedly reduced the survival rate of mouflon. Variations in pedestrian index accounted for variations in harvests when excluding epizootic events. Both surveys detected a decrease in population size, which accounted for the recent increase of harvest. Helicopter and pedestrian surveys are reliable tools to monitor annually mouflons in mountainous areas. Simulations indicated that helicopter surveys should be preferred by managers because they provide the best trade-off between cost and precision.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Assessing population abundance or density is essential for studying population dynamics and for efficient wildlife management (Wilson et al. 1996; Schwarz and Seber 1999; Williams et al. 2002). However, monitoring changes in the size of wild populations remains difficult for wildlife ecologists (Link and Sauer 1997; Pollock et al. 2002). Despite a long history of refinements in design and development of census methods (Caughley 1977; Seber 1982; Eberhardt and Simmons 1987; Lancia et al. 1994; Buckland et al. 2000; Pollock et al. 2002), few attempts have provided satisfactory results except for capture–mark–recapture methods (Schwarz and Seber 1999) and distance sampling (Buckland et al. 2004). However, for most species, mark–recapture or mark–resight techniques are costly and time-consuming (Link and Sauer 1997). Moreover, distance sampling is not well suited to 3D areas such as mountains. Approaches using count statistics have thus been developed as alternatives to census methods.
Count statistics are well adapted when cost and effort to estimate total population size are prohibitive and only the relative differences in abundance are required (Eberhardt and Simmons 1987; Pollock et al. 2002; Williams et al. 2002). Count statistics include numerous methods, such as number of birds seen and heard at a point-count location, number of ungulates seen while walking a transect, or number of small mammals caught on a trapping grid (Nichols et al. 2000). The relationship between a count statistic and the population abundance can be written as C I =N I P I , where C I denotes the count, N I the true abundance, and P I the detection probability, all associated with time and location I (Lancia et al. 1994). If a standardized method is used to obtain the count statistic, and the detection probabilities are similar across time and locations sampled (i.e. that P I =P for all I in the comparison), then the count statistics provide reliable index of abundance (Nichols et al. 2000; Williams et al. 2002).
The wild population of mouflon (Ovis gmelini musimon × Ovis sp.) of the Caroux-Espinouse massif (south of France) has been monitored since 1989 (Santosa 1990) through a pedestrian survey derived from bird surveys based on point estimates (Blondel et al. 1970). Additionally, helicopter surveys have been conducted yearly since 1994. We aim to test the relevance of such census techniques to track trends in population sizes. During our study, population size varied according to an epizootic of keratoconjunctivitis in autumn 1993 (Cugnasse 1997), which reduced the survival probability of all age and sex classes (Cransac et al. 1997). Moreover, hunting occurs annually and quotas have recently increased in response to increase in damage caused by mouflon (Fig. 1). To assess the accuracy of the methods, we tested whether the pedestrian survey was sensitive to the decrease of population size after the keratoconjunctivitis die-off. We also expected that both surveys would detect the same trend in population size and track variation in yearly harvest. Further, we used simulations to optimize the monitoring protocol used to calculate indices and we compared the cost of each type of survey to identify the method providing the best trade-off between cost and precision.

Total number of male and female mouflons removed annually through hunting from 1988 to 2003. Because hunting occurred from September to February, year reported corresponds to the end of hunting season (i.e. 1989 corresponds to 1988/1989 hunting season)
Materials and methods
Study area
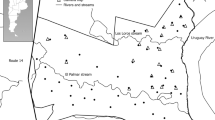
The study area was situated on the south-western border of the Massif Central, in southern France (Fig. 2). Elevations ranged from 300 m to 1,124 m. Mouflons occupy Caroux-Espinouse massif (43°40′N, 3°0′E), which covers approximately 17,000 ha. This population grew from 19 individuals introduced between 1956 and 1960 in the wildlife reserve of Caroux-Espinouse (1,708 ha), situated in the central part of the massif (Cugnasse and Houssin 1993). No other introduction occurred in nearby mountain massifs. Hunting (Fig. 1), by stalking and beating, was based on quotas and occurred from September to February. Quotas are completely harvested each year. A mean number of 107 (±34) males and 110 (±45) females were removed annually between 1973 and 2003. The population is monitored by the Office National de la Chasse et de la Faune Sauvage since 1974 (Cugnasse and Houssin 1993).

Location of the study area in southern France. Transects (and observation points) sampled during pedestrian surveys (for the sake of clarity only the sunset transects were reported) and route sampled during helicopter surveys were reported
Climatic conditions consisted of dry summers (Garel et al. 2004) and dominant south–southeast winds, wet autumns and fairly cold winters associated with dominant north–northwest winds (Thiebaut 1971). The vegetation includes an irregular mosaic of beech (Fagus silvatica), chestnut (Castanea sativa) and evergreen oak (Quercus ilex) forest in a north–south gradient. The high plateaus have been replanted with coniferous woodland (Pinus sylvestris, P. nigra, Picea abies). The vegetation in open areas is dominated by moorlands of heather (Calluna vulgaris, Erica cinerea) and broom moorlands (Cytisus purgans, C. scorparius) frequently mixed with grasses (e.g. Festuca paniculata, F. ovina, Agrostis capillaris) and whortleberry (Vaccinium myrtillus). Between 1955 and 1992, the decrease in pastoral activity and fire suppression has allowed encroachment of mouflon range by dense broomland and forests (V. Bousquel, Syndicat Interchambre Montagne Elevage, unpublished data). During this period, the area of open moorland decreased by 49% (from 4,830 ha to 2,378 ha).
Survey procedures
Pedestrian survey
We estimated population trends between 1989 and 2003 from punctual abundance index (PAI) estimated from a pedestrian survey (Santosa 1990). The PAI was calculated from days of intensive large-scale systematic census during the spring, following lambing period when large groups occur in open areas (Bon et al. 1990). Twelve transects were defined across the entire massif in open areas and were surveyed during the period of maximal activity of mouflon (Santosa 1990; Bon et al. 1991). Six were simultaneously surveyed within 2 h after sunrise, while the other six were simultaneously surveyed during the 2 h before sunset (e.g. Fig. 2). About three to four observation points were distributed along each transect. Each transect was surveyed by one observer, in the same way to prevent observation problems related to sunlight. From the 40 observation points (n=21 for sunrise transects and n=19 for sunset transect and), each observer searched and located mouflon for 15 min with binoculars (10×42 mm). Each sample area was independent from each other. The procedure was repeated over consecutive days between three and nine times (median=6) depending on weather conditions encountered each spring. In 1992 and 1999 no data were collected due to adverse weather conditions that obscured observation (wind, rain and fog). Since 1998, the surveys have been based on sunset transects alone to reduce the effort spent in the field. For each day, PAI was calculated per transect as the sum of the number of individuals observed on all points.
Helicopter survey
We conducted helicopter surveys of the open areas of the entire massif between 1994 and 2003 (Fig. 2), ensuring visual independence between observations. Each mouflon was recorded during the flight (between 23 min and 60 min, median=32 min) to estimate an aerial abundance index (AAI). The 5.0 km route was surveyed 2 h before sunset when mouflons were most active. The speed (30–50 km/h) and altitude (20–30 m) were maintained constant in all flights, allowing us to assume high and equal detectability of age and sex classes (LeResche and Rauch 1974) and to prevent double counts (mouflons join vegetal cover during the passage of the helicopter and remain below). To limit disturbance, no stops were performed. Helicopter doors were removed to improve observation. The helicopter survey took place each spring immediately after pedestrian surveys and was repeated over consecutive days between three and nine times (median=5.5). The same observer performed all aerial counts. For each census day, the AAI was calculated as the sum of the number of individuals observed.
Statistical analysis
Model adjustment
Analyses of changes in population size can be biased when factors related to the acquisition of data are not adequately controlled (Link and Sauer 1997). We therefore fitted models with the following factors of variation in the detection of mouflons: date of survey (PAI and AAI), temperature (PAI and AAI), transects (PAI) and duration of the flight (AAI). Temperature (daily mean) and the date of surveys ranged from 6.3°C to 20.3°C and from 13 May to 14 June, respectively, for the pedestrian surveys, and from 11.0°C to 23.0°C and from 5 June to 12 July, respectively, for the helicopter surveys. The date of survey and the temperature are expected to influence counts because these variables influence the use of open areas by mouflons and the intensity of spatial and sexual segregation (Bon et al. 1990; Santosa et al. 1990; Cransac and Hewison 1997; Cransac et al. 1998). We also accounted for differences in PAI and in temperature according to transects due to differences in space use of mouflons and physical characteristics of the sample areas (e.g. orientation, slope, vegetal cover). We discarded censuses (n=14) done during conditions that obstructed observation (e.g. wind, rain and fog, Santosa et al. 1990).
Analysis procedure
We first tested for an epizootic effect on PAI for which data were available both before and after the epizootic event. We then performed analyses by excluding the epizootic year to test whether PAI accounted for variation in yearly harvest and whether both surveys detected the same trend in population size. To compare both surveys, we performed the analysis on a common period (1995–2003) during which the harvest increased (Fig. 1). We believe that hunting quotas settled throughout the study period most likely have a significant effect on the population dynamics. Indeed, a simple Leslie matrix model (Caswell 2000), with optimistic demographic values, no stochasticity and including harvests, predicted a maximum population size of 2,450 mouflons in 1989 when the pedestrian survey started (M. Garel, unpublished data).
Pedestrian survey analysis was based on 751 PAI sampled on 12 transects between 1989 and 2003. During the 1989–1998 survey period, there was no difference in PAI between sunrise transects and sunset transects for the same day of sampling (F1, 454=0.04, P=0.83). Therefore, the two data sets were pooled. Data were log-transformed (after adding one individual to each count because some counts were equal to zero) to ensure a homogeneous variance across treatments (Fligner–Killeen test (Conover et al. 1981) for the homogeneity of variance throughout years: Fk11=20.93, P=0.06; and across transects: Fk11=12.44, P=0.33). We then examined the annual variations of PAI by using linear models. The observer effect was ignored because the number of observers in PAI was large (n=60). The bias associated with differences among observers is indeed likely to be offset by gains in precision obtained when ignoring observer effect (James et al. 1996).
Comparison between pedestrian and helicopter surveys was based on 415 PAI and 43 AAI sampled between 1995 and 2003 (excluding 1999 because poor weather conditions occurred for PAI). We applied the same procedures as used for the analysis of the pedestrian surveys and compared the model selected in both cases.
Model selection
Model selection was based on Akaike Information Criterion (AIC) with second order adjustment (AICc) to correct for small-sample bias (Burnham and Anderson 1998). This criterion is based on the principle of parsimony and is well adapted when performing multiple comparisons between non-nested models. The most parsimonious model (i.e. lowest AICc) was selected as the best model. We followed Burnham and Anderson (1998) to conclude that the models were different when the difference in AICc was >2. When the difference was <2, we kept the simplest model. We also computed Akaike weights (AICc weight) to compare the relative performance of the models rather than only their absolute AICc value. Weights can be interpreted as the probability that a model is the best model given the data and the set of candidate models (Burnham and Anderson 2001). Thus, the strength of evidence in favour of one model over the other is obtained by dividing their Akaike weights.
Simulation procedure to optimize the protocol surveys
We explored how the PAI and AAI estimates depended on the number of repetitions. We chose years with a large number of repetitions (1994, 1996, 1997 for pedestrian surveys and 1997, 1998, 1999 for helicopter surveys). We performed Monte-Carlo simulations, to estimate values of PAI and AAI that would have been obtained for different numbers of repetitions of the surveys. For each year with a total of R repetitions of the surveys, we randomly chose r values (1<r≤R) among the R possible values of total number of mouflons seen per survey. For each value of r, we performed 1,000 samplings of r values among the R possible values. For each sampling, we then estimated the mean number of mouflons seen over r repetitions of the survey. For each r value, we therefore obtained 1,000 values of PAI and AAI. Based on these values, we estimated the coefficient of variation (CV) of the PAI and AAI. The CV of PAI and AAI was used as a measure of the repeatability of the result and was expected to decrease with increasing r.
The cost of each survey was calculated as a function including a constant cost (staff training and organization) and a cost generated by each additional repetition (cost of realization of the survey). For pedestrian survey, we used only sunset transects in the simulation procedure and cost estimate to account for reduction in the time spent in the field since 1998. All analyses and resampling procedures were performed using R 1.8.0 (Ihaka and Gentleman 1996).
Results
Pedestrian surveys (1989–2003 period)
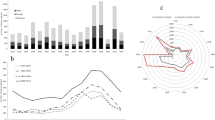
The model with the lowest AICc included an effect of the interaction between temperature and transects, and additive effects of date and year (Table 1). This model was 13 times more supported than the second best model without year effect (AIC weights ratio: 0.928/0.072=12.889). The PAI decreased with the date of observation (slope =−0.018±0.004 (SE), P<0.001). Between 13 May and 14 June, PAI decreased from 24.68±1.77 (SE) to 13.61±1.13. The PAI decreased with temperature (slope=−0.050±0.011, P<0.001) but not in the same way for all transects sampled (Table 1). As expected, the decrease of PAI following the epizootic (autumn 1993) was the most important ever recorded: PAI was significantly less in 1994 than in any other year (Fig. 3).

Temporal variation in the punctual abundance index (PAI) predicted from the selected model (see Table 1, model including epizootic effect) in the mouflon population of the Caroux-Espinouse massif, France. Coefficients of the year effect were reported to show the year to year variations in PAI (1989 corresponds to reference year, i.e. null value). In 1992 and 1999 no data were collected due to adverse climatic conditions (wind, rain and fog)
The variation in harvest best accounted for the variation from year to year in PAI (slope=−0.00093±0.00044, P=0.03) after excluding 1994 (Table 1). This model was over three times more supported than the second best model without a harvest effect (AIC weights ratio: 0.781/0.214=3.650). When the harvest increased from 149 to 427 animals, the PAI predicted from the selected model decreased from an average of 24.2±3.4 to 18.5±2.5 per transect for a survey that would occur at the median date of survey (27 May) and median temperature (12.5°C).
Comparison between pedestrian and helicopter surveys (1995–2003 period)
For pedestrian surveys, the model with the lowest AICc included an effect of the interaction between temperature and transects, and additive effects of date and harvest (Table 2). This model was three times more supported than the second best model without a harvest effect (AIC weights ratio: 0.746/0.249=3.000). The PAI tended to decrease when the harvest increased (slope=−0.00095±0.00053, P=0.07). When the harvest increased from 185 to 427 animals, the PAI predicted from the selected model decreased from an average of 24.0±3.6 to 19.4±2.9 per transect for a survey that would occur at the median date (24 May) and median temperature (13.3°C).
For helicopter surveys, the model with the lowest AICc included additive effects of date, temperature, duration and harvest (Table 2). This model was over 13 times more supported than the second best model without a harvest effect (AIC weights ratio: 0.909/0.068=13.368). The AAI decreased with the date of survey (slope=−0.03±0.009, P=0.001), temperature (slope=−0.16±0.03, P<0.001) and tended to increase with the duration of survey (slope=0.032±0.01, P=0.08). As for the pedestrian surveys, the AAI decreased when the harvest increased (slope=−0.0032±0.0011, P=0.008). When the harvest increased from 185 to 427 animals, the AAI predicted from the selected model decreased from an average of 116.2±1.1 to 54.5±1.2 for a survey that would occur at the median date (16 June), median temperature (16.0°C) and median duration (31 min).
Simulation procedure to optimize the surveys
The curves linking the CV to the number of repetitions r were similar for all years (Fig. 4a, b). The CV tended to be less variable for AAI than PAI. The simulations for PAI and AAI indicated that increasing the number of surveys decreased the variation in the mean number of animals seen from one random sample to another. Assuming that the same number of repetitions is needed to obtain the same precision for AAI and PAI estimates (Fig. 4a, b), AAI was more cost effective than PAI (difference of 34.1% between PAI and AAI costs, Table 3). Pedestrian surveys were more complex than helicopter surveys and involved at least six observers. Therefore, difference in cost between the two methods came from the additional cost of organization, training and implementation of pedestrian surveys (Table 3).

Relationship between the number of repetitions of the survey for 3 years (1994, 1996 and 1997 for PAI, and 1997, 1998, and 1999 for AAI), the cost of repetition and the CV of the PAI (a) or the CV of the AAI (b).The CV was estimated from 1,000 values of a random sample of r values of the index (PAI or AAI) among the possible observed values of the index for a given year (see text for details). For the cost of the survey, see Table 3
Discussion
Validation of the surveys and changes in mouflon abundance
To use pedestrian and helicopter surveys as reliable indices of population abundance, we assumed that such indices are directly proportional to the population size. Therefore, it is necessary to assume that the probability of detection was constant across locations and years which rarely occur in practice (Lancia et al. 1994; Nichols et al. 2000; Pollock et al. 2002; Williams et al. 2002). The increasing closure and degree of fragmentation of habitat likely modified the distribution of resources by reducing food availability for mouflons, which in turn could changed social and spatial organization of the animals. Alternatively, the detection probability of mouflons might have been reduced with increasing habitat closure. There are basically multiple approaches to deal with failure of the detectability assumptions (Nichols et al. 2000; Farnsworth et al. 2001; Pollock et al. 2002). We chose to adjust our models with date of sampling, temperature and transects to make detectability constant across locations and years. For example, animals reduce their activity and stay in dense cover when temperature increases (Santosa et al. 1990) causing the detection probability to decrease (as supported by the negative relationship we reported between temperature and the two indices). In addition, other factors may have been involved, such as human disturbance, or repartition of resources (Santosa et al. 1990; Rubin et al. 1998), and not all covariates can be measured, modelled or even perceived. In this context, some authors have suggested to develop a monitoring design, which includes an estimation of detection probability (e.g. Nichols et al. 2000; Farnsworth et al. 2001; Pollock et al. 2002). This was however not feasible in our study.
The precision of indices is also often difficult to check due to a lack of reference estimates on “true” population sizes, for example those obtained by capture–mark–recapture methods (e.g. Vincent et al. 1991). Nevertheless, one can use independent information about variations in population size to check whether the indices used track known variations. Both abundance indices were sensitive to observed changes in population size resulting from an epizootic (Cugnasse 1997) and from changes in harvest. Hunting affect population abundance directly by animal removal and possibly indirectly by influencing reproduction and survival (Festa-Bianchet 2003). Indeed, most (>90%) harvested mouflons were trophy males and females >3 years of age (M. Garel et al., unpublished data); increasing hunting pressure reduced the probability of adult survival and could thus have affected population growth (Gaillard et al. 2000). Moreover, even if there is a lack of information on the impact of trophy hunting on population dynamics (Harris et al. 2002; Festa-Bianchet 2003), some evidence suggests that it affects ungulate reproduction (e.g., Solberg et al. 2002; Sæther et al. 2003) and thus growth rate of the population (Festa-Bianchet 2003).
Management implications
To date, comparative tests of helicopter and ground surveys are rare for ungulates and mainly concern gender and age composition data (Tsukamoto 1977; Bender et al. 2003). We suggest that helicopter surveys provide a better alternative to census mouflon populations than pedestrian surveys because:
-
1.
The AAI detected similar trends in population abundance as PAI and strongly accounted for yearly changes in harvest.
-
2.
Only one observer is involved in AAI, so the observer bias is limited compared to the PAI.
-
3.
The AAI provided the best trade-off between cost and precision. It required fewer man-days of sampling, and therefore, were much more cost-effective (Table 3) for assessing mouflon abundance over a large area with an equal number of repetitions for AAI and PAI (Fig. 4a, b). Moreover, training is limited to the first year for AAI, whereas for PAI, it is difficult to work annually with the same observers (60 different observers were used during 1989–2003 vs. 2 for helicopter surveys during 1994–2003). Therefore, the difference in cost between surveys in favour of AAI tends to increase after the first year.
-
4.
The AAI allows one to reduce technical/security problems related to the access to observation points (arrival before the sunrise and back after the sunset) in large, mountainous areas.
Finally, previous comparative studies of data obtained from helicopter and ground surveys reported that helicopter counts were both more representative than ground counts and corroborated by demographic studies (Bender et al. 2003 on elk (Cervus elaphus) and mule deer (Odocoileus hemionus)).
References
Bender LC, Myers WL, Gould WR (2003) Comparison of helicopter and ground surveys for North American elk Cervus elaphus and mule deer Odocoileus hemionus population composition. Wildl Biol 9:199–205
Blondel J, Ferry C, Frochot B (1970) La méthode des indices ponctuels d’abondance (IPA) ou des relevés d’avifaune par “stations écoute”. Alauda 38:55–71
Bon R, Gonzales G, Im S, Badia J (1990) Seasonal grouping in female mouflon in relation to food availability. Ethology 86:224–236
Bon R, Cugnasse JM, Dubray D, Gibert P, Houard T, Rigaud P (1991) Le mouflon de Corse. Terre et Vie (Rev Ecol) S6:67–110
Buckland ST, Goudie IBJ, Brochers DL (2000) Wildlife population assessment: past developments and future directions. Biometrics 56:1–12
Buckland ST, Anderson DR, Burnham KP, Laake JL, Borchers DL, Thomas L (2004) Advanced distance sampling. Oxford University Press, Oxford
Burnham KP, Anderson DR (1998) Model selection and inference: a practical information-theoretic approach. Springer, Berlin Heidelberg New York
Burnham KP, Anderson DR (2001) Kullback-Leibler information as a basis for strong inference in ecological studies. Wildl Res 28:111–119
Caswell H (2000) Matrix population models: construction, analysis and interpretation. 2nd edn. Sinauer Associates, Sunderland
Caughley G (1977) Analysis of vertebrate populations. Wiley, London
Conover WJ, Johnson ME, Johnson MM (1981) A comparative study of tests for homogeneity of variances, with applications to the outer continental shelf bidding data. Technometrics 23:351–361
Cransac N, Hewison AJM (1997) Seasonal use and selection of habitat by mouflon (Ovis gmelini): comparison of the sexes. Behav Proc 41:57–67
Cransac N, Hewison AJM, Gaillard JM, Cugnasse JM, Maublanc ML (1997) Patterns of mouflon (Ovis gmelini) survival under moderate environmental conditions: effects of sex, age, and epizootics. Can J Zool 75:1867–1875
Cransac N, Gerard JF, Maublanc ML, Pepin D (1998) An example of segregation between age and sex classes only weakly related to habitat use in moufon sheep (Ovis gmelini). J Zool 244:371–378
Cugnasse JM (1997) L’enzootie de kérato-conjonctivite chez le mouflon méditerranéen (Ovis gmelini musimon × Ovis sp.) dans le massif du Caroux-Espinouse (Hérault) à l’automne 1993. Gibier Faune Sauvage 14:569–584
Cugnasse JM, Houssin H (1993) Acclimatation du mouflon en France: la contribution des réserves de l’Office National de la Chasse. Bull Mens Off Nat Chass 183:26–37
Eberhardt LL, Simmons MA (1987) Calibrating population indices by double sampling. J Wildl Manage 51:665–675
Farnsworth GL, Pollock KH, Nichols JD, Simons TR, Hines JE, Sauer JR (2002) A removal model for estimating detection probabilities from point-count surveys. Auk 119:414–425
Festa-Bianchet M (2003) Exploitative wildlife management as a selective pressure for the life-history evolution of large mammals. In: Festa-Bianchet M, Apollonio M (eds) Animal behavior and wildlife conservation. Island Press, Washington, pp 191–207
Gaillard JM, Festa-Bianchet M, Yoccoz NG, Loison A, Toigo C (2000) Temporal variation in fitness components and population dynamics of large herbivores. Annu Rev Ecol Syst 31:367–393
Garel M, Loison A, Gaillard JM, Cugnasse JM, Maillard D (2004) The effects of a severe drought on mouflon lamb survival. Proc R Soc Lond B 271(suppl):S471–S473
Harris RB, Wall WA, Allendorf FW (2002) Genetic consequences of hunting: what do we know and what should we do? Wildl Soc Bull 30:634–643
Ihaka R, Gentleman R (1996) R: A language for data analysis and graphics. J Comput Graph Stat 5:299–314
James FC, McCulloch CE, Wiedenfeld DA (1996) New approaches to the analysis of population trends in land birds. Ecology 77:13–27
Lancia RA, Nichols JD, Pollock KH (1994) Estimating the number of animals in wildlife populations. In: Bookhout TA (ed) Research and management techniques for wildlife and habitats, 5th edn. The Wildlife Society, Bethesda, pp 215–253
LeResche RE, Rausch RA (1974) Accuracy and precision of aerial moose censusing. J Wildl Manage 38:175–182
Link WA, Sauer JR (1997) Estimation of population trajectories from count data. Biometrics 53:488–497
Nichols JD, Hines JE, Sauer JR, Fallon FW, Fallon JE, Heglund PJ (2000) A double-observer approach for estimating detection probability and abundance from point counts. Auk 117:393–408
Pollock KH, Nichols JD, Simons TR, Farnsworth GL, Bailey LL, Sauer JR (2002) Large scale wildlife monitoring studies: statistical methods for design and analysis. Environmetrics 13:105–119
Rubin ES, Boyce WM, Jorgensen MC, Torres SG, Hayes CL, O’brien CS, Jessup DA (1998) Distribution and abundance of bighorn sheep in the Peninsular Ranges, California. Wildl Soc Bull 26:539–551
Sæther BE, Solberg EJ, Heim M (2003) Effects of altering sex ratio structure on the demography of an isolated moose population. J Wildl Manage 67:455–466
Santosa Y (1990) Utilisation de paramètres eco-ethologiques pour la mise au point d’une méthode d’étude quantitative des populations de grands mammifères: exemple du mouflon (Ovis ammon musimon) du Caroux-Espinouse. Doctoral Thesis, Université de Université Paul Sabatier (Toulouse III), France
Santosa Y, Maublanc ML, Cugnasse JM, Eychenne D (1990) Influence de facteurs climatiques sur le rendement d’échantillonnages de mouflons (Ovis ammon musimon). Gibier Faune Sauvage 7:365–375
Schwarz CJ, Seber GAF (1999) Estimating animal abundance: review III. Stat Sci 14:427–456
Seber GAF (1982) The estimation of animal abundance and related parameters, 2nd edn. Griffin, London
Solberg EJ, Loison A, Ringsby TH, Sæther BE, Heim M (2002) Biased adult sex ratio can affect fecundity in primiparous moose Alces alces. Wildl Biol 8:117–128
Thiebaut B (1971) La transition climatique dans le massif de l’Agoût. Vie Milieu 22:167–206
Tsukamoto G (1977) Evaluation of methods used in determining deer population trends. Pittman-Robertson Job Performance Report W-48-7, Nevada Department of Fish and Game, Reno, Nevada
Vincent JP, Gaillard JM, Bideau E (1991) Kilometric index as biological indicator for monitoring forest roe deer populations. Acta Theriol 36:315–328
Williams BK, Nichols JD, Conroy MJ (2002) Analysis and management of animal populations: modelling, estimation, and decision making. Academic Press, San Diego, California
Wilson DE, Cole FR, Nichols JD, Rudran R, Foster MS (1996) Measuring and monitoring biological diversity. Standard methods for mammals. Smithsonian Institution Press, Washington, DC
Acknowledgments
We thank the Office National de la Chasse et de la Faune Sauvage (DER, Service départemental, BMI), the Groupement d’Intérêt Environnemental et Cynégétique du Caroux-Espinouse, the Office National des Forêts, the Fédération départementale des chasseurs de l’Hérault, the Institut de Recherche sur les Grands Mammifères, and the team of Fagairolles for their participation in the field. We also thank B. Milhau (Office National de la Chasse et de la Faune Sauvage) for his participation in the helicopter survey. Thanks to L. Ellison who checked the English language of the manuscript. We acknowledge E. J. Solberg for his comments on previous drafts of this work. M. Garel was supported by the Office National de la Chasse et de la Faune Sauvage and the Fédération départementale des chasseurs de l’Hérault.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Garel, M., Cugnasse, J.M., Loison, A. et al. Monitoring the abundance of mouflon in South France. Eur J Wildl Res 51, 69–76 (2005). https://doi.org/10.1007/s10344-004-0075-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10344-004-0075-7