
Abstract
The need to preserve and enhance biodiversity in agricultural landscapes is widely accepted. In the case of maize, there is a chance to combine production and biodiversity in the same field by microsegregation: wildflowers are sown between the future maize rows after harvest of the last main crop. These wildflower strips provide flowers and vegetation structure within fields and favour biodiversity without losing production area. The system is based on reliable cropping techniques such as strip-till, underground fertilization and band spraying, allowing the poor habitat quality of conventional maize caused by late soil tillage and lack of vegetation structure to be overcome. Field trials at two sites in Germany were conducted, testing the agricultural feasibility, ecological efficiency and yield impacts. The results show the successful establishment of the wildflower strips between the maize rows. Flowering diversity was up to eight times higher than in conventional maize crop stands. Positive implications for pollinators and ground beetles could be proved. The habitat quality for the skylark could be improved by a factor of 2–3, to nearly normal reproduction of the population. A yield reduction of at least 30% was observed. Further investigations will address this yield gap. Furthermore, the management of spontaneous weeds needs further improvement.
Zusammenfassung
Biodiversität in Agrarlandschaften zu schützen und zu fördern, ist ein weithin akzeptiertes Ziel. Hier wird eine Möglichkeit vorgestellt, im Maisanbau hohe Produktionsleistung mit hoher Biodiversität auf derselben Fläche zu kombinieren. Dazu werden zwischen die Maisreihen Wildpflanzen gesät, bereits nach der Ernte der letzten Hauptfrucht. Diese Blühstreifen stellen Blüten und Vegetationsstruktur innerhalb des Maisfelds bereit und fördern so viele Bereiche der Biodiversität ohne Flächenverlust für die Produktion. Das Anbausystem gründet sich auf gut eingeführte Anbautechniken wie Strip-Till, Unterflurdüngung und Bandspritzung. Diese Techniken werden so kombiniert, dass die schlechte Habitatqualität von konventionell angebautem Mais, bedingt durch die späte Bodenbearbeitung und das Fehlen von Vegetationsstruktur, deutlich aufgewertet wird. Es wurden an 2 Versuchsstandorten in Deutschland Feldversuche durchgeführt, um die landwirtschaftliche Machbarkeit, ökologische Effizienz und Auswirkungen auf die Erträge zu prüfen. Die Ergebnisse der Feldversuche zeigen, dass die Blühstreifen zwischen den Maisreihen erfolgreich etabliert werden konnten. Das Blütenangebot war mit dem neuen System bis zu 8‑mal höher als im konventionellen Maisanbau und stand die ganze Anbausaison zur Verfügung. Positive Wirkungen wurden für Bestäuber und Laufkäfer festgestellt. Die Habitatqualität für die Feldlerche erhöhte sich um den Faktor 2–3 und führte damit zu einer beinahe normalen Reproduktion der Population. Es wurde eine Ertragslücke von mindestens 30 % gemessen, die in weiteren Versuchen verringert werden soll. Auch das Management von Unkräutern in den Blühstreifen muss weiter verbessert werden.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The decline in biological diversity in European agricultural landscapes has been well documented in regional and Europe-wide studies (Defra 2013; Meyer et al. 2013). Projections of future development predict further declines (Pereira et al. 2010) to be strongest in areas cropped with annual crops (de Baan et al. 2013). Recent agricultural developments, like the promotion of bioenergy crops, are regarded as drivers or even accelerators of further biodiversity loss (Flade 2012; Leadley et al. 2010; Robertson et al. 2012).
The continuously decreasing biodiversity has triggered intense discussions concerning appropriate and efficient countermeasures. The search for options to achieve both conservation of biodiversity and highly productive agriculture is the objective of the “segregation or integration” discussion for biodiversity-promoting measures. While one side promotes ways for cash crop production sharing the area with biodiversity (e.g. Andreasen and Andresen 2011), the other side prefers to set aside arable areas for biodiversity separate from areas designated for production (e.g. Berger et al. 2003; Meyer et al. 2013).
Recent results have shown that the “either–or” debate is a simplification of the problem, disregarding regional biodiversity, land use settings and interactions (Dauber and Miyake 2016). In many cases, nature conservation targets are incompatible with modern land use (Haber 2016). Furthermore, the uptake of greening measures—an instrument to include nature conservation in farming practices—by farmers is much higher when they can be integrated into existing agricultural production (Pe’er et al. 2017). The question remains, how to combine promotion of biodiversity within a highly productive agricultural system without losing land for production. Here, a new approach to combine high biodiversity and intensive agricultural production of maize using “microsegregation” is presented.
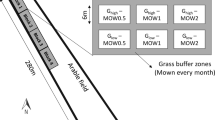
The field area is divided (segregated) into fine-scale “production” and “wildflower” strips (Fig. 1). The production strips are stocked with maize rows, while the area between them, the wildflower strip, is used to grow rows of wildflowers. These wildflowers provide vegetation structure and other floristic resources and form a key factor of biodiversity (Benton et al. 2003; Wilson et al. 2005; Reeleder et al. 2006; Patterson and Best 1996, Wagner and Schmidt 2016) within the maize field. Maize was chosen because this crop combines agronomic feasibility, a large cropping area and therefore impact in Germany (StaBu 2018), low biodiversity value (e.g. Glemnitz et al. 2015; Saska et al. 2014; Tissier et al. 2016) and ecological potential (see below).

Principle of wildflower strips and production strips on a maize field by microsegregation for enhanced biodiversity in maize fields. Here, stem elongation has just started
From the wildlife point of view, the wide row spacing in maize offers open access to the soil surface in a part of the year when other crops have a much too high and too dense vegetation layer (Hoffmann et al. 2018). In Germany, maize is typically sown at the end of April. During the time between the harvest of the pre-crop and the sowing of maize, the fields stay unused or are sown with catch crops, which die during winter. This long period can be used to establish wildflowers beneficial for the wildlife and with only low competition with maize, so there is cover during winter and flowering can start early in the next year (Fig. 2). Because maize is predominantly grown in rows of 0.75 m width, the wildflowers can remain in the field by using strip-till for maize seeding and band spraying of herbicides to spare the wildflower strips. Both herbicide application and late tillage cause reduction of biodiversity in conventional maize cropping (McLaughlin and Mineau 1995). Using microsegregation the wildflower strips stay undisturbed for over one year. Additionally, the very low intensity of insecticide applications (0.1–0.2 applications per year, Roßberg 2016) enables insects to utilize the available food resources. Microsegregation might turn maize crop into a source of biodiversity.

Timeline for vegetation development of the different plants (cover crop, sown wildflowers, maize)
In early stages maize is very sensitive to competition by weeds (Cerrudo et al. 2012; Page et al. 2012). The challenge for the success of the microsegregation approach is to establish and fine-tune the wildflower strips in order to limit the competition from the wild plants sown in the wildflower strips, especially during the “critical period” in the early maize growth stage. Besides, the basic idea of the microsegregation approach is that a certain non-crop biomass is tolerated by maize and does not reduce the yield more than acceptable at any given point in time, even in the critical period.
The selection of species for the wildflower strips is one tool to influence the competition between the sown wildflower mixture and the maize crop. An appropriate selection of wildflowers that build up a flowering chain in the early vegetation stages and provide small-scale habitat structures such as nectar, pollen and host plants is very important for the expected effects on biodiversity. Thus, they should be ecologically effective but must not become problematic weeds in any regard. Moreover, the selected wildflower plants should be able to suppress competitive weeds to a low density. In addition, the species selected have to meet technological demands such as feasibility with agricultural sowing techniques. Because many wildflowers show a slow early development, non-hardy cover crops are sown simultaneously with the wildflowers to support them through suppressing weed infestation and volunteer crops. Combined with strip-till, underground fertilization is provided below the maize only. Accordingly, fertilization will not enhance biomass production of the wildflower strips, where especially well-adapted weeds would profit. The combination of the selected technological measures ensures a soil tillage break and only very limited impacts of fertilization and chemical weed control on the wildflower strips.
The impact of maize cropping on overall biodiversity has essentially increased with the growing acreage cropped with maize (Gevers et al. 2011). There are different approaches to how to stabilize the early maize growing and to reduce environmental risks like erosion (Laloy and Bielders 2010) or nitrate leaching (Justes et al. 2012). However, there are hardly any approaches working on improvement of the effects of maize cropping on biodiversity or wildlife. One exception is Norris et al. (2016, 2018), who tested different cultivation systems including strip-till regarding their effects on arthropods and pollinators. The current approach of “microsegregation” aims to improve maize cropping for wildlife without losing area for production.
Material and Methods
Location, Soils and Climate of the Sites
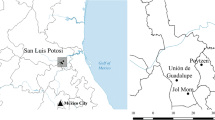
The microsegregation cropping system was tested at the two experimental sites Dedelow and Müncheberg in northeast Germany (Fig. 3) in plot trials from late summer 2016 to autumn in 2017.

Location of the experimental sites Dedelow and Müncheberg in northeast Germany
The geomorphology and soil structure of both experimental sites contains deposits of the last ice age (Weichselian glacial) and of post-glacial processes but with different characteristics. The soil types pallid soil and pseudogley are dominant, which have been formed from strong loamy sands and loam of the ground moraine. The climatic conditions of the experimental sites were characterised by a transition between oceanic and continental climate. Both sites are typical for the precipitation-limited climate conditions in Middle Europe. The two sites vary mainly regarding soil conditions and soil fertility. An outline of the long-term averages of temperature, precipitation and basic soil conditions is given in Table 1.
The investigational season 2016/2017 was characterized by only small deviations from the long-term average weather conditions. The end of 2016 was warmer and more humid than the average. In winter there were two short periods of frost in December and January. Maize sowing was followed by a period of low temperature and no rainfall.
Experimental Design
At both experimental sites, field trials were performed in an equal block plot design. The single plots were fully randomised and every variant was repeated four times. The tested variants consisted of combinations of the following four test factors:
-
Composition of the wildflower mixture (2 mixtures)
-
Kind of cover crop over winter (3 crops: field bean, cress, buckwheat)
-
Sowing density of cover crops over winter (2 levels: one row per habitat strip, full area)
-
Width of wildflower strips (2 levels: 1‑row/25 cm, 2‑row/35 cm), width of production strip (50 cm; 40 cm)
The experiments did not realise full combinations of all the test factors; see summary of applied variants and their combinations of the single test factors in Table 2.
Wildflower Selection
The selection of species for the wildflower strips was oriented towards plant traits gained from available plant trait databases (e.g. BfN 2018; Jäger 2011). First of all, the plant species were selected according to their life cycle (only biannual or perennial) and regional natural occurrence. Priority was given to high feed values for insects as a nectar and pollen source and a long flowering period over the spring and summer. Moreover, the wildflower species should be of low final biomass and low plant height, thus with low competition with maize. Finally, the species lists were screened for the traits sowing depth, seed size and germination time in late summer/autumn, to meet the technological demands of the current approach. Species known as a phytosanitary risk to any neighbouring or subsequently grown crop (noxious weeds, host plants for common diseases) were excluded from the list. At the last step, the availability of seeds with regional origin was applied as a selection criterion. An additional desired function of the selected wildflowers was the suppression of spontaneous weeds growing in the wildflower strips. As result of the selection process, 28 different plant species were identified and composed into two seed mixtures consisting of the identical species but varying in the quantity of the specific traits (Table 3). In mixture 1 (M1), 80% of the total sowing quantity was seeds of tall-growing species, while in mixture 2 (M2), seeds of low-growing species accounted for 80% of the sowing quantity.
All management measures in the experiments were carried out with technical solutions which are available in agricultural practice. Sowing of the wildflowers was realized with a seed drill for fine seeds, the cover crop was sown with conventional seed drills. Productions strips were cleared up in spring with band spraying machinery applying glyphosate. Soil in the production strips was tilled with strip-till machinery. Fertilization of the production strips was performed simultaneously with the maize sowing as underground fertilization, using the same machinery. After sowing, herbicides were applied for a second time in the production strips only. No other pesticides were applied. For details of the particular technological measures applied in the experiments, please see at Appendix A and B (Tab. 7 and 8).
Surveyed Parameters
Data elevation was based on well-established methods and followed a unified protocol. Table 4 presents the selection of the observed parameters as addressed in this paper.
Wildflowers and Weeds
Weeds—and sown wildflowers—form the basis of biodiversity on arable fields. Species diversity was measured. For the quantification of the flower supply, every species was categorized: 1: just flower buds (early flowering); 2: some individuals of a plant species flower; 3: full-flowering species (nearly all individuals of a species are in flower). For further methodological details see Table 4.
Wild Bees and Hoverflies
According to their different ecological traits and functions, wild bees and hoverflies were selected to be investigated among pollinators. Besides pollination, hoverflies are efficient predators of aphids. Both wild bees and hoverflies show a high mobility in space, some species move 150 to 600 m randomly around their reproduction habitat (Gathmann and Tscharntke 2002), which is normally a natural habitat. Due to this high mobility, pollinator traps (e.g. yellow traps) in small-sized plots may actively attract flying insects and thereby distort the observation results. To avoid these effects, a direct method to measure effects on these insects was chosen, i.e. counting the insects sitting on flowers during a 60-min time span at four different dates with a two-week separation. This method is limited in the number of replications. Therefore, the pollinator activity density of any variant of the wildflower strips was compared 1) with a maize field without wildflower strips, 2) with a pure wildflower mixture and 3) with the habitats in the surroundings of the field trials.
Carabid Beetles
The diversity and activity density of carabid beetles were investigated during the early growing stages of maize crop stands from June to July. Investigations were done in Variants (Var.) 1, 3 and 10 at Müncheberg. The period coincides with the second part of the breeding season for most of the regional farmland birds like skylark.
Farmland Birds—Example of Skylark (Alauda arvensis)
The abundance and reproduction of the skylark are closely related to crop type and the vegetation structure (Wilson et al. 2005), as well as the extent and timing of soil tillage and pesticide use. Therefore, habitat suitability of four variants of the trials was modelled for the skylark. The potential abundance of skylarks was estimated by observing the vegetation (Hoffmann et al. 2016). The vegetation height and density was measured weekly during the breeding season beginning in the middle of March and ending at the end of July. Daily vegetation values where modelled using the method of Hoffmann et al. (2018). Data on farm management were included additionally. The algorithms obtained were used to allocate abundances to measured heights and degrees of coverage of the variants. The assessment of the habitat effects of the variants takes into account three breeding cycles of the skylark per season, each lasting for 40 days, as well as the date and the extent of soil tillage and herbicide application.
On the basis of the field data, an evaluation matrix was applied to quantify the habitat effect and the habitat quality level of the single variants. The habitat quality level, derived from the calculated habitat effect (%), describes the habitat effects for the skylark in the variants in relation to reproduction. Five groups were derived: strong sink, sink, moderately sink, balanced, source and strong source. Sink stands for insufficient reproduction (<100%), balance for adequate reproduction (±100%) and source for expanded reproduction of the population (>100%; Hoffmann and Wittchen 2018). The assessment was carried out for Var. 1, 3, 10 and 12 (see Table 2). In addition to these four selected variants of the field trials, the variant “set aside” was included as an ecological reference. This was carried out completely without herbicides, fertilisation and tillage.
Maize Yield
The maize was harvested by hand at 1.33 m long row sections (1 m2) to measure the yield. Harvest was performed at the end of August.
Data Analysis
All surveyed data except the pollinator counts and the model results for the farmland birds were statistically tested. The yield data were analysed by generalized linear models (GLM) with normally distributed error variance and the Tuckey HSD multiple-mean test. For the ecological parameters GLM were performed, which are robust for a non-normal data distribution as typical for ecological data. For data with count values statistical models based on Poisson distribution and Wald chi2 tests were applied. For percentage values, the inverse normal value distribution was used. In the last case, mean values were tested with the maximum likelihood quotient. The presented results are based on estimated means from GLM analyses. In all GLM analyses, the experimental site and the test variants were used as fixed factors.
For the interpretation, only those results were selected which achieved the highest variance explanation, as measured with the corrected R2-value. All statistical analyses were performed with the software packet SPSS Statistics 22.0 (IBM, Armonk, NY, USA; 2016).
Results
Sown Wildflowers
All 28 species included in the seed mixtures 1 and 2 germinated in the wildflower strips. The frequency of the single species varied. Nine species were very frequent und 11 species occurred regularly. Only the manifestation of eight species was restricted: three species occurred infrequently and five rarely.
First wildflowers started flowering already shortly before maize sowing. In maize at BBCH stage 0–39 (germination, leave setting and longitudinal growth) at both sites, 12–22 species from the sown wildflowers were flowering in one plot. The number of flowering species and intensity of flowering dropped with ongoing ripening of the crop (Fig. 4).

Number of flowering sown wildflower species in relation to BBCH stages of maize. a Site Müncheberg, b site Dedelow. Flowering category see chapter “Wildflower selection”. BBCH phenological development stage of crop plants, cat. category
The variants showed no significant differences in the number of flowering wildflowers. The flowering chain lasted from day 124 to day 207 in the year at Müncheberg, from day 105 till day 244 at Dedelow. At both sites a well-balanced mixture of four or five different flower colours provided by the sown wildflowers was found.
Weeds
The species number of spontaneous weeds was on average two times higher in variants with sown wildflower strips between the maize rows compared to maize without wildflower strips over the whole growing season. In Var. 1, up to 20 weed species were found, in the variants with wildflower strips there were 34–40 species at site Dedelow and 37–50 species at site Müncheberg. In contrast to the complete period, the differences in weed species number were much higher at the specific development stages of the maize stands (Fig. 5). The statistical analyses revealed significant differences of the weed species number between the two sites, the two sowing mixtures, the width of the wildflower strips and the kind of cover crop over winter. Weed species number was slightly but significant higher at Dedelow. Especially in variants with the tall-growing M1 sowing mixture and in variants with the broader wildflower strips, the species number increased. At Dedelow, field bean (whole area) as cover crop resulted in higher species diversity for weeds in the subsequent growing season, at the site Müncheberg the same was found for buckwheat and cress cover crop.

Number of weed species in the variants at different development stages of the maize crop. Accumulated value of four replications per variant. a Site Müncheberg, b site Dedelow. BBCH phenological development stage of crop plants
Wild Bees and Hoverflies
The investigation of wild bees and hoverflies showed a clear ecological surplus provided by the sown wildflower strips (Fig. 6). While in conventional maize hardly any pollinators on a weed flower were found, the sown wildflower strips provide a habitat quality comparable in effect size with pure wildflower plots. At Dedelow, the flower strips in the maize field attracted just 20% less pollinator individuals than flowers in the semi-natural habitats of the surroundings. At Müncheberg the effect was lower. There, just 40% of the wild bees’ activity could be found in maize with wildflower variants.

Cumulative number of individuals sitting on flowers in surrounding habitats, pure wildflowers, maize and wildflowers and pure maize. a Site Müncheberg, b site Dedelow
For hoverflies, wildflower strips in the maize rows provided a similar habitat quality as a pure wildflower mixture or the surrounding habitats.
The wildflower strips attracted mainly unspecialized but also some highly specialized (oligolectic) wild bee species. All species can be very important for the pollination of some field crops and fruits. While at Müncheberg 72 bee species were found in the semi-natural habitats of the surroundings, 30 species were found in the flower strips in the maize field and 31 species in the pure wildflower variant. At Dedelow, 51 wild bee species were discovered in the surroundings, 39 species in the flower strips in the maize field and 43 species in the pure wildflower mixture.
Besides bees, some unspecialized species of hoverflies were observed at both sites. At Müncheberg, 15 species were found in the semi-natural habitats of the surroundings, 14 species in the flower strips in the maize field and 9 species in the pure wildflower variant. At Dedelow, 17 hoverfly species were found in the surroundings, 11 species in the flower strips in the maize field and 18 species in the pure wildflower mixture.
Carabid Beetles
The differences in the carabid beetle species number between the variants were not significant. Var. 3 showed significant more species then Var. 1 when the single measurements were compared (Table 5), but during the observation, 34 species could be found in every variant. The activity density showed a significant effect between the variants. Even in Var. 1, both parameters, i.e. the carabid beetle species number and activity density, showed relatively high values. During the investigation period, 1.5 times more individuals were found in the 25 cm (Var. 10) wide and more than twice as many individuals in the 35 cm (Var. 3) wide wildflower rows compared to conventional maize stands.
Habitat Effects on Skylark
Full-surface soil cultivation leads to a complete disturbance of the first breeding cycle in Var. 1. Depending on the width of the wildflower strip, an area of 33 or 47% remains undisturbed by tillage and herbicide application and the vegetation structure therefore varied. The variants with wildflower strips led to significantly higher habitat quality compared to the conventional variant (Table 6). These variants showed differences in the range of the improvement of the habitat effect. The lowest effect was found in Var. 1, the highest in Var. 10. Var. 1 acts as strong sink for the reproduction of the skylark population. The ecologically improved variants were characterised as moderately sink and balanced. The habitat function of the ecologically improved variants increased up to three times in relation to Var. 1. The added variant set aside proved to be a strong source for expanded reproduction of the skylark population.
Yield of Maize
At both experimental sites the yield of Var. 1 was highest. The variants with microsegregation showed nearly no significant differences in maize yield. Only the cover crop field beans sown in rows showed significant yield reduction in most of the variants (Fig. 7). The variants 2, 4 and 5 showed the highest yield and reached 63–70% of the yield of Var. 1. This group is characterized by the cover crop field beans on full area and either narrow wildflower strip rows (Var. 2, 4) or the low-growing wildflower mixture (Var. 4, 5). Var. 3 and 7 with cover crop field beans, tall-growing wildflowers and a wide wildflower strip—only the sowing density of the field beans differ—had the lowest maize yield. Theses variants are regarded the ones with the highest competition to the maize crop.

Average yield of the variants (four replicates per variant, result of Tuckey HSD test). a Müncheberg, b Dedelow. a, b, c, d, e significant contrasts of group means
Discussion
The raison d’être of the microsegregation approach is a positive ecological effect. In this regard, the sown wildflower strips were very successful. The flowering started very early in the season and lasted until maize harvest. Even after harvest the wildflowers were flowering again. Most of the species flowered consistently, but with differences between the experimental sites most likely caused by the differing soils and precipitation patterns. The width of the strips did not affect the complex flowering pattern; thus, a small strip provides nearly the same pattern as a broad one. Therefore, future trials can concentrate on the small strip. Weeds were found in conventional maize as well as in microsegregation maize. The difference in weed species numbers was relatively small for the complete growing season (~double of the species number in microsegregation). The view on the single observation dates indicates that the weed species in the reference variant (Var. 1) were not only found in lower numbers (up to 8 times lower), the species number rather tended to decrease than to increase over time. In Var. 1, the species number was at a low level during the vegetation period, reduced by tillage. The other variants had even increasing numbers of weed species. These weeds even increase the difference of flowering shown in Fig. 4. Confirming results from Norris et al. (2016) in England, sown wildflowers and weeds together formed a highly diverse and consistent habitat, which is the basis for other organism groups as the presented results of the current investigation prove.
Wildflowers in addition to weeds in the wildflower strips attracted a big part of the pollinator community occurring in the surroundings of the field. While conventional maize was an absolutely hostile area for wild bees, the microsegregation maize attracted at least 40% of the wild bee species that occur in the natural habitats in the surroundings of the experimental fields; for hoverflies the effect was even stronger, even though the species diversity in hoverflies is not as high as in bees in open (agricultural) landscapes. Similarly, Norris et al. (2018) showed more pollinator visits in a wildflower intercropped maize compared to a conventional maize field. The diversity of species of wild bees proved the high value of microsegregation for this species group.
Similar to hoverflies and in contrast to the wild bees, carabid beetles occur regularly in conventional maize crops and all variants showed the same species number within the observation. Nevertheless, the wildflower strips had a clear positive effect on their activity density. The number of caught individuals increased in the plots with wildflower strips, especially with the broader wildflower strip. These results are in accordance with those of Norris et al. (2016), who found arthropod richness and density increased by applying strip-till and sowing of additional plants into the maize rows. This effect was not caused by the reduction of soil tillage but through the sown companion plants.
Skylarks essentially benefit from the introduced vegetation structure. Conventional maize only exceptionally allows successful breeds and thus serves as strong sink for skylarks. This is because the skylarks start breeding in maize but soil tillage prior to maize sowing destroys the nests. Avoiding full area soil tillage allows the skylarks to finish breeding and shifts the habitat value close to the reproduction of the population. The highest habitat values for the skylark were detected for variants with low wildflower row width, which also have a low weed density. The simulation results suggest that the skylarks don’t need the broad wildflower strip. They profit from the narrow strip, which fits well with the findings for the flowering aspect. The positive effect on carabids and vegetation diversity adds confidence in the simulation results. In many cases, a very important reason for unsuccessful breeding is predation (Wilson et al. 1997). Since the effect of vegetation structures on predation is a complex issue (Wilson et al. 2005), proving the effect of microsegregation on predation remains a future challenge. The yield gap between conventional maize and microsegregation is a weak point at the current development stage of the new cropping system. A yield reduction of at least 30% was observed, whereas there are relatively large yield differences between the microsegregation variants. The variants with field bean as cover crop (whole area) had the highest yield, while the lowest yield was reported for Var. 7 with field bean (1-row), which was established in Dedelow only. At both sites, Var. 3 and 11 had relatively low yields, reaching approximately 50% of the control. The general influence of the non-hardy cover crop stresses the importance of the time before winter for the maize yield in maize with microsegregation. Norris et al. (2016) reported a yield gap of 30–45% for an experiment with strip-till variants using ryegrass or diverse crop seed mixtures as companion crops. Similar yield effects have been communicated from cropping systems with traditional undersowings or from mixed cropping, as reviewed by Verret et al. (2017). In Germany, Nurk et al. (2017) reported relevant yield gaps in maize–beans mixed cropping. These consistent results of reduced yield raise the question of whether the yield equivalence is a realistic objective for this kind of cropping system. Nevertheless, reducing the yield gap is a challenge for further development of the cropping system.
A major reason for the maize yield gap is most likely the competition by weeds and wildflowers. The biomass of both reduced the yield. While the wildflowers are selected for a low competition value, the weeds are highly competitive. Therefore, a high diversity of weed species combined with low abundances, especially of problematic weeds, was a desired effect of the wildflower selection. Weed control through living mulches or intercropping is possible (Hartwig and Ammon 2002; Verret et al. 2017), but the selected seed mixtures failed to accomplish this goal. Additionally, the weeds germinating in autumn in the wildflower strips might not be typical maize weeds. Therefore, problems could rise in controlling them chemically due to missing approved herbicides. At both experimental sites, some plots were dominated by specific weeds, though microsegregation needs further improvements in controlling the weeds and the biomass in the wildflower strips. To reduce the competition, the selection of wildflowers is a promising tool (von Cossel and Lewandowski 2016). Norris et al. (2016) additionally suggest precise tillage and adjusting of the intercropping row widths.
Besides weed control (Cerrudo et al. 2012), water and nitrogen supply are most sensitive processes in maize early development, which need explicit attention. Negative effects on these parameters of mixed cropping with maize or undersowings are not regularly the case but can’t be prevented totally (Conrad and Fohrer 2016; Justes et al. 2012). The long growing time of the wildflower strips over winter might be able to reduce nitrogen leaching effectively (Komainda et al. 2018).
An important criterion for the microsegregation in maize fields was agricultural feasibility, to ensure its uptake from farmers (Pe’er et al. 2017). The single agrotechnical measures should be feasible with existing machinery. Considering this, the use of fine seed machinery for sowing the wildflower mixtures, band spraying techniques for preparing and managing the weed control within the production strips and strip-till machinery for soil tillage, fertilization and sowing maize only at the production strips was successful. Measures additional to conventional maize cropping could be applied between workload peaks without time pressure or competition for machines. This proved the feasibility of the microsegregation approach for farmers. The challenge to working exactly on the different strips can easily be mastered through GPS tracking techniques, which are already available at many farms. Still, microsegregation is much more complex than what is known about undersowing or intercropping cultivation systems (Hartwig and Ammon 2002; Norris et al. 2016). The main differences for microsegregation are the following: sowing mixtures have to be compiled according to special ecological and agrotechnical traits, wildflowers have to be sown in the autumn of the previous year, the wildflowers need a cover crop for weed control during autumn, fertilization and chemical weed control should be spatially restricted to the maize production strips.
Microsegregation is a relatively elaborate production system. Several measurements are necessary while the crop yield is most likely reduced compared to conventional cropping systems. On the other hand, the additional ecological return is huge. It is known that cropping systems with intercropping or similar approaches can be very expensive and possibly not profitable for farmers in Germany (Nurk et al. 2017), even though resources are saved through the strips (Tauchnitz et al. 2018). Therefore, the economic frame of microsegregation should be defined in future work. This may include the agro-environmental policy of the EU (CAP), for which the current approach provides options for “greening”, since it combines yield and quality of maize with the promotion of biodiversity. Therefore, microsegregation fulfils some requirements for sustainable farming (Andreasen and Andresen 2011; Tscharntke et al. 2012). Since farmers may use it without losing area for production, even the uptake of this cropping system as a greening measure has a high prospect of success (Pe’er et al. 2017).
Conclusion
The complex system presented here cannot be run in a static way. It needs adaption to different conditions at different sites: the soil and weather conditions differ; different problematic weeds need attention; the settings of the farms are different. To further examine the possibilities and obstacles of microsegregation trials in contrasting regions, specific plant mixtures are the appropriate next step. To enhance the practical aspect, the trials should cover fields rather than small plots. Important objectives to investigate are the yield, the suppression of volunteer weeds, the control of the sown wildflowers, as well as the economic and ecological effects of microsegregation. This huge effort is reasonable because microsegregation promises to change the cropping system altogether, which is a difference to most of the “greening” measures. Even though a yield gap was found, it was shown that this approach successfully combines high-yield production with strongly enhanced biodiversity.
References
Andreasen C, Andresen L (2011) Managing farmland flora to promote biodiversity in Europe. Cab Rev 6:1–11. https://doi.org/10.1079/pavsnnr20116047
de Baan L, Alkemade R, Koellner T (2013) Land use impacts on biodiversity in LCA: A global approach. Int J Life Cycle Assess 18(6):1216–1230. https://doi.org/10.1007/s11367-012-0412-0
Benton TG, Vickery JA, Wilson JD (2003) Farmland biodiversity: Is habitat heterogeneity the key? Trends Ecol Evol (Amst) 18:182–188
Berger G, Pfeffer H, Kaechele H, Andreas S, Hoffmann J (2003) Nature protection in agricultural landscapes by setting aside unproductive areas and ecotones within arable fields (“Infield Nature Protection Spots”). J Nat Conserv 11:221–233
Bundesamt für Naturschutz (BfN) (2018) FloraWeb – Daten und Informationen zu Wildpflanzen und zur Vegetation Deutschlands. http://www.floraweb.de. Accessed 11 Oct 2018
Cerrudo D, Page ER, Tollenaar M, Stewart G, Swanton CJ (2012) Mechanisms of yield loss in maize caused by weed competition. Weed Sci 60:225–232
Conrad Y, Fohrer N (2016) Simulating impacts of silage maize (Zea mays) in monoculture and undersown with annual grass (Lolium perenne L.) on the soil water balance in a sandy-humic soil in Northwest Germany. Agric Water Manag 178:52–65
von Cossel M, Lewandowski I (2016) Perennial wild plant mixtures for biomass production: Impact of species composition dynamics on yield performance over a five-year cultivation period in southwest Germany. Eur J Agron 79:74–89
Dauber J, Miyake S (2016) To integrate or to segregate food crop and energy crop cultivation at the landscape scale? Perspectives on biodiversity conservation in agriculture in Europe. Energy Sustain Soc 6(1):25
Defra (2013) Defra national statistics release: Wild bird populations in the UK, 1970 to 2012. https://www.gov.uk/government/organisations/department-for-environment-food-ruralaffairs/series/biodiversity-and-wildlife-statistics (Created 17 Oct 2013). Accessed 11 Oct 2018
Flade M (2012) From ‘Renewable Energies’ to the biodiversity disaster—comments on the current situation of bird conservation in Germany. Vogelwelt 133:149–158
Gathmann A, Tscharntke T (2002) Foraging ranges of solitary bees. J Animal Ecol 71(5):757–764
Gevers J, Hoye TT, Topping CJ, Glemnitz M, Schroeder B (2011) Biodiversity and the mitigation of climate change through bioenergy: Impacts of increased maize cultivation on farmland wildlife. Glob Change Biol Bioenergy 3:472–482
Glemnitz M, Zander P, Stachow U (2015) Regionalizing land use impacts on farmland birds. Environ Monit Assess 187(6):336. https://doi.org/10.1007/s10661-015-4448-z
Haber W (2016) Flächenansprüche – Wie erfüllt man wachsende Ansprüche an begrenzte Landflächen? Schriftenr Dtsch Landeskulturges 14:23–40
Hartwig NL, Ammon HU (2002) Cover crops and living mulches. Weed Sci 50(6):688–699
Hoffmann J, Wittchen U (2018) Abschätzung der Habitatwirkung veränderter Produktionsverfahren auf Indikatorvogelarten der Ackerbaugebiete im Forschungsvorhaben “Maisanbau für hohen Ertrag und biologische Vielfalt” am Beispiel der Feldlerche (Alauda arvensis). Berichte aus dem Julius Kühn-Institut, vol 195. Julius Kühn-Institut, Kleinmachnow
Hoffmann J, Wittchen U, Stachow U, Berger G (2016) Moving window abundance—a method to characterize the abundances dynamics of farmland birds: The example of the skylarks (Alauda arvensis). Ecol Indic 60:317–328
Hoffmann J, Wittchen U, Stachow U, Berger G (2018) Moving window growth—a method to characterize the dynamic growth of crops in the context of bird abundance dynamics with the example of the skylark (Alauda arvensis). Ecol Evol 8(17):8880–8893. https://doi.org/10.1002/ece3.4398
IBM (2016) SPSS statistics for windows, version 22.0. IBM Corp, Armonk
Jäger EJ (ed) (2011) Rothmaler – Exkursionsflora von Deutschland, 20th edn. Spektrum Akademischer Verlag, Heidelberg
Justes E, Beaudoin N, Bertuzzi P, Charles R, Constantin J, Dürr C, Hermon C, Joannon A, Le Bas C, Mary B, Mignolet C, Montfort F, Ruiz L, Sarthou JP, Souchère V, Tournebize J, Savini I, Réchauchère O (2012) The use of cover crops in the reduction of nitrate leaching: Impact on the water and nitrogen balance and other ecosystem services. Summary of the study report. INRA, Paris
Komainda M, Taube F, Kluß C, Herrmann A (2018) Effects of catch crops on silage maize (Zea mays L.): Yield, nitrogen uptake efficiency and losses. Nutr Cycl Agroecosyst 110(1):51–69
Laloy E, Bielders CL (2010) Effect of intercropping period management on runoff and erosion in a maize cropping system. J Environ Qual 39(3):1001–1008
Leadley PW, Pereira HM, Alkemade R, Fernandez-Manjarrés JF, Proença V, Scharlemann JPW, Walpole MJ (2010) Biodiversity scenarios: Projections of 21st century change in Biodiversity, and associated ecosystem services: A technical report for the global Biodiversity outlook 3. Technical series, vol 50. Secretariat of the Convention on Biological Diversity, Montreal
McLaughlin A, Mineau P (1995) The impact of agricultural practices on biodiversity. Agric Ecosyst Environ 55(3):201–212
Meyer S, Wesche K, Krause B, Leuschner C (2013) Dramatic losses of specialist arable plants in Central Germany since the 1950s/60s—a cross-regional analysis. Divers Distrib 19:1175–1187
Norris SL, Blackshaw RP, Dunn RM, Critchley NR, Smith KE, Williams JR et al (2016) Improving above and below-ground arthropod biodiversity in maize cultivation systems. Agric, Ecosyst Environ, Appl Soil Ecol 108:25–46. https://doi.org/10.1016/j.apsoil.2016.07.015
Norris SL, Blackshaw RP, Critchley CNR, Dunn RM, Smith KE, Williams J, Randall NP, Murray PJ (2018) Intercropping flowering plants in maize systems increases pollinator diversity. Agric For Entomol 20(2):246–254
Nurk L, Graß R, Pekrun C, Wachendorf M (2017) Effect of sowing method and weed control on the performance of maize (Zea mays L.) intercropped with climbing beans (Phaseolus vulgaris L.). Agric (switzerland) 7:51. https://doi.org/10.3390/agriculture7070051
Page ER, Cerrudo D, Westra P, Loux M, Smith K, Foresman C, Wright H, Swanton CJ (2012) Why early season weed control is important in maize. Weed Sci 60(3):423–430
Patterson MP, Best LB (1996) Bird abundance and nesting success in Iowa CRP fields: The importance of vegetation structure and composition. Am Midl Nat 135(1):153–167
Pe’er G, Zinngrebe Y, Hauck J, Schindler S, Dittrich A, Zingg S et al (2017) Adding some green to the greening: Improving the EU’s ecological focus areas for biodiversity and farmers. Conserv Lett 10(5):517–530
Pereira HM, Leadley PW, Proença V, Alkemade R, Scharlemann JPW, Fernandez-Manjarrés JF, Araújo MB, Balvanera P, Biggs R, Cheung WWL, Chini L, Cooper HD, Gilman EL, Guénette S, Hurtt GC, Huntington HP, Mace GM, Oberdorff T, Revenga C, Rodrigues P, Scholes RJ, Sumaila UR, Walpole M (2010) Scenarios for global Biodiversity in the 21st century. Science 330(6010):1496–1501. https://doi.org/10.1126/science.1196624
Reeleder RD, Miller JJ, Coelho BB, Roy RC (2006) Impacts of tillage, cover crop, and nitrogen on populations of earthworms, microarthropods, and soil fungi in a cultivated fragile soil. Agric, Ecosyst Environ, Appl Soil Ecol 33(3):243–257
Robertson BA, Porter C, Landis DA, Schemske DW (2012) Agroenergy crops influence the diversity, biomass, and guild structure of terrestrial arthropod communities. Bioenergy Res 5(1):179–188
Roßberg D (2016) Erhebungen zur Anwendung von Pflanzenschutzmitteln im Ackerbau (Survey on application of chemical pesticides in agriculture). J Kultpfl 68(2):25–37
Saska P, Němeček J, Koprdová S, Skuhrovec J, Káš M (2014) Weeds determine the composition of carabid assemblage in maize at a fine scale. Sci Agric Bohemica 2:85–92
Statistisches Bundesamt (Destatis) (2018) Field crops and grassland. https://www.destatis.de/EN/Homepage.html. Accessed 24 Sept 2018
Tauchnitz N, Bischoff J, Schrödter M, Ebert S, Meissner R (2018) Nitrogen efficiency of strip-till combined with slurry band injection below the maize seeds. Soil Tillage Res 181:11–18
Tissier ML, Handrich Y, Robin J‑P, Weitten M, Pevet P, Kourkgy C, Habold C (2016) How maize monoculture and increasing winter rainfall have brought the hibernating European hamster to the verge of extinction. Sci Rep 6:25531. https://doi.org/10.1038/srep25531
Tscharntke T, Clough Y, Wanger TC, Jackson L, Motzke I, Perfecto I, Vandermeer J, Whitbread A (2012) Global food security, biodiversity conservation and the future of agricultural intensification. Biol Conserv 151(1):53–59
Verret V, Gardarin A, Pelzer E, Médiène S, Makowski D, Valantin-Morison M (2017) Can legume companion plants control weeds without decreasing crop yield? A meta-analysis. Field Crops Res 204:158–168
Wagner C, Schmidt C (2016) Blühflächen erhöhen die Tierartenvielfalt in der Feldflur. In: Bayerische Landesanstalt für Landwirtschaft (LfL) (ed) Wildtiere in der Agrarlandschaft 14. Kulturlandschaftstag, Freising, 05.10.2016 Bayerische Landesanstalt für Landwirtschaft (LfL), Freising-Weihenstephan, pp 53–62
Wilson JD, Evans J, Brown SJ, King JR (1997) Territory distribution and breeding success of skylarks Alauda arvensis on organic and intensive farmland in southern England. J Appl Ecol 34(6):1462–1478
Wilson JD, Whittingham MJ, Bradbury RB (2005) The management of crop structure: A general approach to reversing the impacts of agricultural intensification on birds? Ibis (Lond 1859) 147:453–463
Acknowledgements
We thank Katrin Lück, Sigrid Ehlert, Cornelia Fischer, Edelgunde Jerusel, Petra Rischewski, Udo Wittchen and Cora Ksinzyk for their great help with the field work and data processing. This research was partly funded by the Federal Ministry of Food and Agriculture and Consumer Protection in the project “Energiemaisanbau für hohen Ertrag und Biologische Vielfalt (Vorstudie)” (grant 22011914).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
C. von Redwitz, M. Glemnitz, J. Hoffmann, R. Brose, G. Verch, D. Barkusky, C. Saure, G. Berger and S. Bellingrath-Kimura declare that they have no competing interests.
Additional information
C. von Redwitz and M. Glemnitz contributed equally.
Appendices
Appendix A
Appendix B
Rights and permissions
About this article

Cite this article
von Redwitz, C., Glemnitz, M., Hoffmann, J. et al. Microsegregation in Maize Cropping—a Chance to Improve Farmland Biodiversity. Gesunde Pflanzen 71, 87–102 (2019). https://doi.org/10.1007/s10343-019-00457-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10343-019-00457-7