
Abstract
Litterfall is one of the most important fractions of forest net primary production (NPP) and important carbon flux into soil, a large carbon pool in forests worldwide, and believed to account for about one-third of forest NPP. The aim of the study was to determine litterfall amount and carbon input into the soil via litterfall as well as to understand the effects of thinning (removing 20–25% of the initial basal area) and seed cutting (i.e. regeneration cutting with removing 60% of the initial basal area) on litterfall in Scots pine (Pinus sylvestris L.) stands. Litterfall samples were collected three times in a year from 32 sample plots for 5 years in Turkey. Carbon concentrations of the litterfall components were determined by dry combustion. Data were evaluated by one-way ANOVA and multiple regression analysis. As a result, litterfall varied from 1389 kg ha−1 year−1 in young stands to 4488 kg ha−1 year−1 in mature stands. Carbon inputs into the soil changed between 714 kg C ha−1 year−1 and 2289 kg C ha−1 year−1 depending on the development stage and silvicultural treatment applied to the stands. Thinnings reduced litterfall amount at a ratio of 22% in mature stands while had no significant effect in overmature stands with a decrease ratio of 8%. Seed cutting reduced considerably the litterfall amount both in mature and overmature stands. A combination of basal area, site index, and stand age accounted for 75% of the variation in needle litterfall. In conclusion, seed cutting was recommended to do preferably in mature stands instead of overmature ones and thinning to be applied lightly in mature stands in order not to reduce carbon input into the soil via litterfall dramatically.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Litterfall is the most important source of nutrient and carbon flux to the forest floor and has long been known as a good indicator of forest ecosystem productivity as well as forest health (Tobin et al. 2006). Litterfall is one of the most important fractions of forest NPP and important carbon flux into the soil, which represents a large carbon pool in forests worldwide. About one-third of annual carbon uptake is transferred to the soil by litterfall (Neumann et al. 2018). Litterfall production is essential for understanding the carbon (Muukkonen 2005) and nutrient cycling in forest ecosystems. Litterfall is the major supply of carbon input to the soil together with root decomposition which is of great importance to sustain the soil fertility as well as nutrient cycling (Diaz-Pines et al. 2011). Carbon is stored in the soil, forest floor, biomass, and dead wood pools in a forest ecosystem (Ravindranath and Ostwald 2008). Soil carbon stock is more difficult to estimate than the other carbon pools due to its heterogeneous structure (Dincă et al. 2015). Many soil carbon models estimating the soil carbon stock and stock change by time need to litterfall input data (Palosuo et al. 2012; Ťupek et al. 2015). A great majority of the studies focused on determining the amount of litterfall and effects of driving factors on litterfall production (Irmak and Çepel 1968; Yang et al. 2004; Roig et al. 2005; Lu and Liu 2012; Bellingham et al. 2013). Although a large number of litterfall studies on Scots pine (Pinus sylvestris L.) in central or northern Europe have been published (e.g. Paavilainen 1987; Albrektson 1988; Cousens 1988; Lonsdale 1988; Kouki and Hokkanen 1992; Finér 1996; Pausas 1997; Berg et al. 1999; Liu et al. 2004; Roig et al. 2005; Starr et al. 2005; Ukonmaanaho et al. 2008; Bārdule et al. 2012; Portillo-Estrada et al. 2013; Novák et al. 2015), data regarding litterfall of Scots pine in the Mediterranean (Alvera 1980; Blanco et al. 2006; Martínez-Alonso et al. 2007; Diaz-Pines et al. 2011) are scarce, especially on the southern limit of Scots pine. It is important to understand the responses of Scots pine, one of the most widely distributed tree species in the world (McGeever and Mitchell 2016), to environmental and human factors in different regions, especially in southern, to develop adaptation strategies for climate change in forestry.
In Turkey, Scots pine is the third most important pine species, covering an area of 1,518,000 ha, with its 13% share of the total pine distribution area after Pinus brutia Ten. and Pinus nigra Arnold (OGM 2015), in terms of distribution area and importance of timber production. Litterfall of the two pine species was studied by Erkan et al. (2017) and Koray (2017), respectively. However, there is a lack of knowledge on litterfall of whole lifetime of Scots pine stands in Turkey.
On the other hand, determining forest management effects on litterfall is of also great importance to understand how human activities will influence nutrient cycling and carbon sequestration in a changing climate. However, little is known about the effects of management practices on litterfall and carbon input into the soil as well as nutrient cycling (Lado-Monserrat et al. 2016). Although some studies have reported litterfall amount in young pine forest ecosystems treated with different thinning intensities (Roig et al. 2005; Blanco et al. 2006; Navarro et al. 2013), there is lack of knowledge concerning the effects of other silvicultural intervention such as seed cutting, which is a widely used silvicultural technique in natural regeneration of mature and overmature Scots pine stands in Turkey, on litterfall. In recent years, forest management planners have tended to give priority to seed cutting in overmature stands compared to mature stands in Turkey due to the concept of even-aged forest management planning. However, the area of overmature stands is still increasing, indicating that different concepts are needed to achieve sustainable forest management.
There are few studies explaining the variation in litterfall by different independent variables. Litterfall models based on climatic parameters such as precipitation and temperature can be useful at regional scales but not at local scales due to lack of local climatic data. Some regression equations were developed based on site index, stand age and latitude, with the highest R2 of 0.65 for Scots pine by Albrektson (1988). In Fennoscandia, Berg et al. (1999) determined some linear relationships between litterfall and site index, basal area and latitude as predictor variables, but R2 of the regressions remained below 0.35. They obtained the equation by combining some pine species, including Austrian pine (Pinus nigra Arn.), lodgepole pine (Pinus contorta Dougl.), Monterey pine (Pinus radiata D. Don), maritime pine (Pinus pinaster Ait.), red pine (Pinus resinosa Ait.) and stone pine (Pinus pinea L.), that all of them had different branches and needles characteristics. Most of the equations estimating the litterfall amount rely on variables that are difficult to obtain. There is a need for functions based on less variable and easy to measure parameters to estimate litterfall amount. Although litterfall amount has not routinely determined, the most common parameters measured in forest inventory in Turkey are basal area, site index, and stand age. Thus, directly measurable parameters seem to be a promising tool for estimating the litterfall in forest ecosystems more accurately.
Objectives of the study were as follows: (1) to provide data on the annual amount of litterfall in natural Scots pine stands in Turkey, one of the distribution area of the species at southern sites; (2) to evaluate the effect of different cutting regimes on litterfall and carbon input into the soil; (3) to analyse the allometric relationship between the annual amount of litterfall and stand characteristics which are easy to measure in forest inventory. We hypothesised that litterfall production: (1) differs along the lifetime of a stand; (2) decreases with cutting intensity; (3) relates to some stand attributes. In this study, we compared litterfall from young, pole stage, mature and overmature stands which were unthinned to understand the litterfall change over time. Thus, almost entire lifetime of the managed forest stands was covered. In addition, litterfall in mature and overmature stands in which thinning and seed cutting were carried out previously was compared with litterfall of unthinned ones to determine the effect of silvicultural treatment on litterfall dynamics.
Materials and methods
Study site
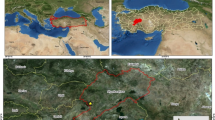
The study was carried out in pure and evenly aged Scots pine stands on Sundiken Mountain in Turkey, located at latitudes between 39º57ʹ04ʺ and 39º59ʹ14ʺN and longitude between 31°03ʹ43ʺ and 31°09ʹ31ʺE (Fig. 1). Sample plots were chosen from southern and northern aspects, with elevations ranging from 1220 m to 1679 m.

Study site [reorganised after EUFORGEN (2009)]
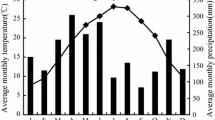
Precipitation and temperature values were obtained from Turkish State Meteorological Service. The nearest meteorological station locates at an elevation of 1388 m with long-term observation in Mihalıççık town, about 35 km far from the study area (Fig. 1). According to the data of this station, mean annual precipitation (P) is 486 mm, with a mean temperature (T) of 9.5 °C. Although this station does not represent the climate of the study site, extrapolation of the mean temperature data was in line with the data of station at study site. Thus, data of the meteorological station established within the study site in 2010 were used to identify the climatic characteristics of the study area. According to the observations during the study period, P ranged from 383 to 565 mm, and T from 7.0 to 8.7 °C (Table 1). The study area is characterised by a continental climate with low rainfall. The soil in the study site is a well-drained luvisol according to IUSS Working Group WRB (2006) originating from mica schist, with a sandy loam texture and pH of 5.6–6.0.
The stands derived from natural regeneration with ages ranged from 12 to 158 year old in 2011. Sample plots were chosen in young, pole stage, mature and overmature stands, in which local forestry directorate performed all silvicultural interventions 5–8 years before the study. Young stands were not exposed to any treatment, while no or light precommercial thinning was performed in pole stage stands. Moderate low thinning with a removal rate of 20–25% of the initial stand basal area was carried out in 2005, while seed cutting was employed in 2008, leaving 100–250 seed trees, corresponding to a reduction of 60% of the initial stand basal area, in mature and overmature stands. Some properties of the stands are shown in Table 2.
Sampling procedure
Sample plots were chosen from young, pole, mature and overmature stands with four replications, 16 plots in totally (4 development stages × 4 replications). To investigate the effects of silvicultural treatments, an extra 16 plots were taken from thinned and seed cut mature and overmature stands (2 treatments × 2 development stages × 4 replications). Thus, 32 sample plots were taken for the study in totally, taking into consideration site quality from poor to good to represent a wide range of site condition. Sample plots differing in size according to the development stage of the stands were chosen to cover the variations in age and diameter to reduce the time of working. For example, an area of 5 × 5 m was preferred for sampling young stands while 10 × 10 m for pole stage stands and 20 × 20 or 30 × 30 m for mature and overmature stands. Different plot sizes may affect the results due to variation in stand structure and environmental factors. Therefore, sample plot size was adjusted so as to have at least 20 individuals to relieve plot size effect on the results.
Litterfall was collected from May 2010 to May 2016 with five 0.5 m × 0.5 m traps placed randomly at each sample plot. These traps were nylon with 1 × 1 mm openings and set 60 cm above the forest floor to hinder decomposition. Litter traps were constructed pyramid-shaped with a minimum 50 cm in depth to prevent litter loss due to turbulent air movement. Litterfall samples were collected in May, August, and November because reaching sample plots was impossible at least 5 months of a year due to heavy snowfalls. Diameter at breast height (dbh) and height of all trees in samples plots were measured in 2010. Basal area of the stands was calculated from dbh of the trees in sample plots and then converted into m2 per hectare. Needle biomass of each tree in sample plot was estimated by following equation developed by Çömez (2010) for Scots pine in the study area, previously:
where y is needle biomass (kg tree−1) and dbh is diameter at breast height of the tree (cm). Needle biomass of sample plot was calculated by individual biomass and expressed in kg ha−1.
After digging a soil pit in each sample plot, soil samples were taken from the Ah and Ale horizons by a steel cylinder with a volume of 100 or 1000 cm3 according to horizon thickness removing the forest floor layer to identify the site properties of the study area.
Site indexes of the stands were calculated through the mean age and height of the 3–5 tallest trees in the sample plot and by taking the corresponding tree top height at the age of 100 years to the measured ones from yield tables of Scots pine developed by Alemdağ (1967).
Laboratory procedures
Litterfall samples were oven-dried until constant weight at a temperature of 70 °C and then separated into needles, bark, branches, cones, and other parts called as miscellaneous, and each portion was weighted. The samples were ground in a mill to pass through a 0.5-mm sieve. Composite litterfall samples were provided by taking into account the contribution of each year litterfall to the total weight of 5 years for carbon analysis. Oven-dried litterfall samples were analysed for carbon by LECO CNH analyser (Leco Corporation, St. Joseph, Michigan). Soil samples were first air-dried and weighted and then a sieved through a 2-mm mesh. Approximately, 10 g of subsample was taken and dried at 105 °C in a drying oven until constant weight to determine moisture content for a correction factor to use in bulk density, carbon and nitrogen analyses. Carbon and nitrogen content of the soil samples was analysed by LECO CNH analyser. Soil pH was measured by pH metre in a 1:5 soil to water solution (volume:volume). Soil texture was determined by hydrometer method (Kroetsch and Wang 2008).
Data analysis
Normality of the data was checked by Shapiro–Wilk test, while the homogeneity of variance by Levene’s test. In case of normal distribution of the data, variations in litterfall and carbon input through litterfall evaluated by one-way ANOVA. When data showed non-normal distribution, logarithmic transformation was employed. If the distribution of data set was not normal despite the transformation, Kruskal–Wallis H test was used to evaluate the significant differences. When achieved homogeneity of variance, Scheffe test was used to determine the homogenous groups, otherwise, Games-Howell post hoc test. Annual litterfall production was estimated by multiple regression analysis, using some stand parameters as independent variables. Equations with the highest R2 and the lowest standard error were given in this study. Graphical analyses were used to compare the performance of the equations obtained with other studies. To evaluate the change in litterfall by time, data derived from young, pole stage, unthinned mature and overmature stands, which were fully stock, were used. To assess the effect of thinning on litterfall, data of seed cut, thinned and unthinned mature and overmature stands were used.
Results
Litterfall quantity
There were significant variations in the amount of litterfall by components among the stands (Table 3). The lowest total litterfall was in young stands, while the highest in mature stands unthinned. The litterfall amount of overmature stands was lower than that of mature stands, but the difference was not significant. There was a rising trend from young to mature stage, while the litterfall remained almost stable since the stage of maturity (Fig. 2).

Changes in litterfall by fractions of a needles, b bark, c branches, d cones, and e miscellaneous. Different letters indicate significant differences (P < 0.05) with Scheffe test. Bars indicate standard deviation
The lowest needle litterfall occurred in young stands, while the highest in pole stands. The amounts of needle litterfall in mature and overmature stands were close to that of pole stands. An increase in needle litter was observed from young to older stage of the stands with a nearly constant amount since the pole stage. Needles accounted for 88% (± 2.9%, standard error (SE)) of the total litterfall in young stands, while 55 (± 1.7, SE) and 50% (± 3.6, SE) in mature and overmature stands, respectively. The contribution of needles to total litterfall decreased as the stand aged. Bark litterfall was lowest in young stands. The highest bark litterfall was observed in mature stands. Bark constituted 2% (± 0.6, SE) of the total litterfall in young stands while 10% (± 0.9%, SE) in older stands and remained stable from the pole stage.
The mean branch litterfall varied from young stands to overmature stands. In young stands, 2% of the total litterfall was comprised of branches while as high as 14% in overmature stands. The contribution of the branches to total litterfall increased from young to overmature stands.
Cones were the second largest component of the total litterfall, varying from young stands to mature stands. The contribution of cones to the total litterfall was found to be the lowest (3%) in young stands and the highest (15%) in mature stands. Cone litterfall in overmature stands was lower than in mature stands.
The miscellaneous fraction significantly differed among the stands, with the lowest value in young stands and the highest in overmature stands. Miscellaneous litterfall was observed to increase from young to mature stage and then remained stable as in the other litterfall fractions. The lowest contribution (5%) of miscellaneous to total litterfall was found in young stands while the highest (11%) in overmature stands.
Effect of silvicultural interventions on litterfall
Seed cutting significantly reduced the amount of litterfall, except for cones in mature stands, and for both cones and branches in overmature stands. In case of seed cutting, total litterfall of mature stands was 38% lower than that of unthinned ones (Table 3). In overmature stands, this decrease was more pronounced with 52% of the unthinned ones. Needle litterfall was found to be lower in thinned stands than in unthinned ones for both mature and overmature stands. However, these differences were not statistically significant. Bark litterfall was lower in thinned mature stands than unthinned ones, while there was no significant difference between unthinned and thinned overmature stands. Thinning led to a significant decrease in branch litterfall in mature stands but not in overmature stands. Similarly, thinning reduced significantly total and miscellaneous litterfall in mature stands, while had no significant effect in overmature stands. Mean total litterfall of thinned mature stands was 22% lower compared to unthinned ones (Fig. 3). But in overmature stands, total litterfall of thinned stands was 9% lower than that of unthinned ones.

Effects of silvicultural interventions on litterfall by fractions of a needles, b bark, c branches, d cones, and e miscellaneous. Different letters indicate significant differences (P < 0.05) with Scheffe test. Bars indicate standard deviation
Carbon input via litterfall
Carbon concentrations changed from 487 mg g−1 in miscellaneous to 523 mg g−1 in branches, but the differences were insignificant among the stands (Table 4). Annual carbon input was the least in young stands and the highest in unthinned mature stands (Table 5).
Thinning and seed cutting had a significant effect on carbon input into the soil through litterfall, except for fraction of cone. Differences in carbon input through branch litterfall were significant in mature stands while not in overmature stands. Thinning operation reduced carbon input into the soil via needle, branch, miscellaneous, and total litterfall significantly in mature stands. However, in overmature stands, carbon amount of litterfall was not affected by thinning, despite a slight decrease. Seed cutting operation led to decrease in carbon input via litterfall components both in mature and overmature stands, except for via branches and cones (Fig. 4).

Effects of silvicultural interventions on carbon input into the soil via litterfall by fractions of a needles, b bark, c branches, d cones, and e miscellaneous. Different letters indicate significant differences (P < 0.05) with Scheffe test. Bars indicate standard deviation
Models predicting litterfall production
All models predicting litterfall compartments were significant at a level of P < 0.001. Models explaining the variations in litterfall compartments are presented in Table 6. Model 1.2 accounted for 75.4% of the variance of needle litterfall by using basal area, site index, and age of the stand as independent variables. The same variables explained 92.7% of the variance of the bark litterfall. Increase in basal area, site index and stand age led to a rise in bark and branch litterfall. Stand age explained 37.9% of the variance in cone litterfall alone. Basal area and stand age were linearly correlated with miscellaneous and total litterfall, accounting for 86.9 and 66.6% of the variances, respectively. Site index was another significant variable in predicting litterfall amount, but using a combination of basal area and site index yielded a better result.
Discussion
Litterfall
The amount of all litterfall fractions changed in a wide range because we sampled various stands from young to overmature. Another wide range from 3220 to 4650 kg ha−1 year−1 was observed for total litterfall of northern and southern Scots pine stands, respectively (Neumann et al. 2018), which were similar to our results for pole-staged, mature and overmature stands, indicating an increase from north to south. Mature stands had twofold more total litterfall than young stands, indicating that litterfall amount increases with stand age. However, changes in total litterfall of pole, mature and overmature stands were insignificant. Thus, litterfall seems to increase rapidly in young stage and reach a steady state from the age of 50 s. There are few studies on litterfall of young Scots pine stands (Finér 1996; Novák et al. 2015). Our total litterfall result of young stands was higher than that reported by Finér (1996) in Finland, northern latitudes, 921 kg ha−1 year−1, but lower than that reported by Novák et al. (2015), around 3500 kg ha−1 year−1 for 15 years of age, in the Czech Republic. Considering that litterfall is positively related to precipitation (Lonsdale 1988; Liu et al. 2004; Roig et al. 2005), the lower total litterfall of our study may be due to the climatic condition in our study area where the rainfall and temperature (419 mm and 7.9 °C) are lower than in Týniště (632 mm and 8.3 °C). Furthermore, the study site of Novák et al. (2015) may have supported a higher basal area, amounting to 25 m2 ha−1 in young stands, than that of the present study with a basal area of 6 m2 ha−1.
For pole stands, total litterfall was reported to vary from 1000 to 2860 kg ha−1 year−1 (Finér 1996; Paavilainen 1987; Cousens 1988; Starr et al. 2005; Ukonmaanaho et al. 2008; Portillo-Estrada et al. 2013) for northern Europe, which was less than our result for the similar stands most likely due to the effect of latitude (Albrektson 1988; Berg et al. 1999; Starr et al. 2005; Ukonmaanaho et al. 2008). However, a considerable high total litterfall amount (8800 kg ha−1 year−1) was reported by Santa Regina and Gallardo (1995) for a Humid Mediterranean site in Spain. Blanco et al. (2006) reported a total litterfall of 3986 kg ha−1 year−1 for pole stage Scots pine stands with a continental climate in Spain which is close to our result while a higher value (5533 kg ha−1 year−1) for Mediterranean climate, indicating that climate and stand structure affect on litterfall. The effect of climatic factors, notably temperature and rainfall, on the amount of litterfall was revealed by some researchers (Pausas 1997; Liu et al. 2004; Lehtonen et al. 2008; Portillo-Estrada et al. 2013).
Around 4100 kg ha−1 year−1of total litterfall reported by Pausas (1997) and Martínez-Alonso et al. (2007) in mature Scots pine stands in Spain where located at similar latitudes to our stands was close to our results of mature and overmature stands. Lower values (1750–2400 kg ha−1 year−1) were reported for Finland mature Scots pine stands situated at latitudes of 61–62°N (Paavilainen 1987; Finér 1996; Starr et al. 2005). Krynytska et al. (2017) reported a higher litterfall (5800–6600 kg ha−1 year−1) for mature Scots pine at 50°N latitude in Ukraine than our result, probably due to more fertility and the inclusion of Quercus robur L. and Carpinus betulus L. litterfall.
As for litterfall fractions, the needle litterfall of pole stands in our study was higher than that in Finland, about 1600 kg ha−1 year−1 (Starr et al. 2005; Portillo-Estrada et al. 2013), while lower than in Spain, about 3000 kg ha−1 year−1 (Blanco et al. 2006). In mature stands, however, lower needle litterfall amounts (555–1400 kg ha−1 year−1) were reported for Finland (Albrektson 1988; Kouki and Hokkanen 1992; Finér 1996; Starr et al. 2005; Ukonmaanaho et al. 2008). Needles were the most important contributor to the total litterfall, with 88% in young stands to 50% in overmature stands. That the contribution of needles to total litterfall decreased from young towards mature stage may result from the fact that needle biomass increases rapidly in young stage until the canopy is closed and do not develop further after a certain density is reached (Berg and Laskowsky 2005; Starr et al. 2005).
Bark litterfall determined in our study was lower than that in Bergmann et al. (1998), 704–876 kg ha−1 year−1, in Germany, but comparable to that in Pausas (1997) and Blanco et al. (2006)—between 360 and 600 kg ha−1 year−1, in Spain, for mature Scots pine stands. The lower bark litterfall can be explained by tree growth. Although there is no clear seasonal pattern in bark litterfall, bark fraction increases as the tree stem and branches grow and fall caused by windstorms (Blanco et al. 2006).
Branch litterfall of pole stands detected in our study is lower than the results reported by some researchers for Spain and Germany, about 250–500 kg ha−1 year−1 (Pausas 1997; Bergman et al. 1998), and for southern Finland, 580 kg ha−1 year−1 (Portillo-Estrada et al. 2013). This difference may result from climatic conditions, indicating that climate may considerably influence litterfall. Branch litterfall does not show a regular periodic pattern. It is related, rather, to specific events such as heavy winds, heavy rainfall, or snowfall (Berg and Laskowsky 2005).
Cone litterfall was reported to vary from 40 kg ha−1 year−1 for overmature stands (Lehtonen et al. 2008) to 200 kg ha−1 year−1 for pole stands (Portillo-Estrada et al. 2013) in Finland. Higher amounts (up to 900 kg ha−1 year−1) were observed by some researchers (Pausas 1997; Bergman et al. 1998; Santa Regina and Tarazona 2000; Bārdule et al. 2012) for southern sites, Latvia, Germany, and Spain, which were higher than our results. In contrast to our results, Ayari et al. (2012) reported that the number of cones was negatively related to age and stand density for Pinus halepensis Mill. in Tunisia. Cone litterfall is related to the amount of cone production, as well as climatic factors. In our study, large trees were also the older trees with larger crowns. Some researchers showed that tree size affected cone production for pine trees (Ordóñez et al. 2005; Turner et al. 2007), as in line with our results.
Silvicultural intervention
Seed cutting reduced total litterfall as well as needles, bark, and miscellaneous fractions significantly in mature and overmature stands due to a reduction in aboveground biomass. Similar results were reported by earlier studies in Europe (Blanco et al. 2006) and in the Czech Republic (Slodičák and Novák 2006) for thinning of young Scots pine stand. Navarro et al. (2013) also reported a reduction in litterfall after thinning for young Pinus halepensis stands in the southeastern Iberian Peninsula. In contrast, we found that total litterfalls were similar in thinned and unthinned overmature stands, indicating that mature and overmature stands might not be affected by thinning in the same way. Although total litterfall of mature stands was reduced considerably by thinning, the effect of thinning remained limited in overmature stands. This difference may result from differences in the application of thinning. Mature stands have generally higher stand density than the older ones. Therefore, foresters tend to cut less tree in overmature stand during the thinning. Remaining trees after the thinning may increase photosynthesis and enhance needle production in overmature stands, compensating the litterfall decrease resulting from biomass reduction. As a result, extracting less trees due to less tree number might have resulted in a limited influence on litterfall of overmature stands compared to mature stands. Jiménez and Navarro (2016) suggested that thinning did not significantly change the litterfall of Pinus halepensis over time except for the first year at tree level. Therefore, it seems that thinning reduces litterfall production by decreasing the biomass rather than changing the abscission process. On the other hand, expandig of remaining tree crown after thinning (Smith et al. 1997) might have caused more miscellaneous fraction and thicker branches, thus mitigated the decrease in litterfall in overmature stands.
Many earlier studies suggested that foliage biomass decreases as a result of thinning treatment due to the reduction of litterfall production (Roig et al. 2005; Blanco et al. 2006; Kim et al. 2009). However, a decrease in litterfall caused by thinning is compensated by recovery of the stand canopy after a couple of years (Roig et al. 2005). In contrast to the studies above mentioned, the decrease in needle litterfall was not pronounced in our study, indicating that litterfall pattern might change in mature and overmature stages compared to youth stage in Scots pine. Novák et al. (2013) reported that thinning did not change litterfall amount, but did influence on litterfall composition, with a higher share of needles (89%) in thinned plots, compared to the unthinned ones. In our study, the contribution of needles to total litterfall was found to decrease to 40% with seed cutting. In a study by Blanco et al. (2006), the share of needles in total litterfall showed a slight increase with an increase in thinning intensity, as in our results. But our results also clearly showed that drastic interventions such as seed cutting could considerably alter the composition of litterfall. In young as well as dense stands, cones did not constitute important part of the total litterfall; however, it became a more dominant litterfall component in seed cuts. Thinning results in an increase in photosynthetically active radiation, which triggers cone production. Increased cone litterfall in seed cuts can be attributed to increased light due to the gaps formed by cutting (Krannitz and Duralia 2004).
Carbon input
Carbon concentrations were not significantly different among the stands. However, Neumann et al. (2018) reported a higher value (530 mg kg−1) for southern European conifers than our results probably due to various conifer species. Chemical composition, e.g. lignin content, can vary considerably among the tree species, which influences on carbon concentration of litterfall component. Carbon input into the soil was reported as 1800–2400 kg C ha−1 year−1 (Diaz-Pines et al. 2011) and 2150 kg ha−1 year−1 (Bārdule et al. 2012) for Scots pine mature stands in Spain and Latvia, respectively, which were very close to our results for thinned and unthinned mature stands. The result (1910 kg C ha−1 year−1) reported by Neumann et al. (2018) for European conifers was also within our range. However, Krynytska et al. (2017) found almost twofold (6000 kg C ha−1 year−1) carbon input in Ukrainian Scots pine stands which had similar properties with our stands, probably due to mixed and more fertile stands as mentioned above. In a young Pinus halepensis stand in Spain (Segura et al. 2017), increasing in thinning intensity was reported to reduce carbon input via needle litterfall from 760 kg C ha−1 year−1 in control to 324 kg C ha−1 year−1 in heavy thinned stands. This decrease corresponds to 57%, which was very close to our results. In our study, although seed cutting resulted in a decrease of 55% in carbon input through needle litterfall in mature stands, decrease in total litterfall remained at 37%, due to a noticeable increase in cone litterfall. It seems that a reduction in some litterfall fractions due to the intervention applied to stand may be compensated by the other fractions. By thinning, carbon input was reduced at a ratio of 22% in mature stands while only 8% in the case of overmature stands, indicating that response of the stands to different silvicultural interventions might change according to stand age. Furthermore, litterfall carbon input of mature stands might be more responsive to thinning than that of overmature stands. However, applying a more severe treatment than thinning such as seed cutting led to further reduction in overmature stands than that in mature stands, likely due to less number of trees left in the area after seed cutting.
Litterfall models
As a stand characteristic, basal area was the strongest predictor for all litterfall components, as shown by many earlier researchers (Berg et al. 1999; Starr et al. 2005; Kunhamu et al. 2009; Diaz-Pines et al. 2011; Erkan et al. 2017). Needle litterfall is the dominant component of total litterfall and was estimated by a regression model with an R2 of 0.754, using stand basal area, stand age and site index, easy to measure variables in national forest inventory. Lehtonen et al. (2008) suggested a litterfall model with an R2 of 0.74 that needs very detailed data including an index of pollen cone litter, daily rainfall, and temperature. The model seems impractical in forest carbon inventory due to the lack of information on data required by the model in most of the countries. Neumann et al. (2018) developed a robust litterfall model in which leaf area index (LAI), mean annual temperature and precipitation were used as independent variables for conifers in the northern, central, and the southern regions of Europe. However, this model was not able to be compared with our model due to the lack of LAI information. Jagodziński and Kałucka (2008) modelled LAI of young Scots pine stand on the basis of mean stand diameter and height. But their model biased after 20 years of stand age. Liu et al. (2004) developed a regression model explaining the needle litterfall variation for conifers in Eurasia region that mean annual temperature and annual precipitation were used as predictors, with a rather low R2 (0.231) compared to that of the present study. A more robust model was suggested by Lonsdale (1988) with an R2 of 0.63, predicting leaf litterfall for world forest using latitude, altitude, and precipitation as independent variables. In their study, maximum leaf litterfall was assumed to be 0.89 t ha−1 in the equation. However, all of the needle litterfall values were higher than 0.89 t ha−1 in our study. Another model with an R2 of 0.735 and including latitude, site index and stand basal area as the predictor was developed for pine in Fennoscandia region by Berg et al. (1999). Ågren and Knecht (2001) and Wutzler and Mund (2007) calculated annual litterfall production by multiplying needle biomass with a turnover rate of 0.4 and 0.25 for pine stands, respectively. As seen in Fig. 5, Berg et al. (1999) model and method used by Wutzler and Mund (2007) overestimated needle litterfall of the present study, while the model of Starr et al. (2005) underestimated. The model developed by Albrektson (1988) relying on site index with R2 of 0.65 tended to underestimate needle litterfall at larger basal area than 40 m2 ha−1, with better agreement at an intermediate basal area, likely because site index concealed the effect of basal area on litterfall. Ågren and Knecth (2001) method is also in line with our model except for young stands and stands with a basal area of less than 10 m2 ha−1. All these shifts may result from the fact that silvicultural treatments such as thinning and seed cut alter the forming of new needle and litterfall.

Comparisons with the models predicting a needles and b total litterfall found in the literature
Basal area, site index, and stand age were found to be the best predictors for needle, bark and branch litterfall, while age was the strongest for cones. Needle and total litterfall amount have been shown to be strongly related to the basal area for Scots pine by many researchers (Berg et al. 1999; Liu et al. 2004; Starr et al. 2005; Matala et al. 2008), as in our study. Lehtonen et al. (2004) reported that branch litterfall depended on tree size and stocking density. They found higher branch litterfall in small trees than in large trees in contrast to our results. Higher branch litterfall may be due to larger branch biomass and heavier self-pruning owing to a light deficiency in mature and overmature stands consisting of large trees in our study.
Site index was another good predictor for litterfall production. Good sites produce more biomass than the poor ones with more litter as a consequence (Albrektson 1988). Site index is a result of the combination of temperature, precipitation and soil fertility, which of those affect on tree growth and litter production. In our study, sample plots in good sites were mainly located on the north-facing slope where the water deficit could be less than the southern slopes due to possibly more precipitation and less evapotranspiration as well as better soil condition, i.e. depth and less stony.
Most of the litterfall models have generally developed for regional scales estimations. Therefore, they include variables, which are different from those of presented study, namely latitude, longitude, and some climatic parameters (Lonsdale 1988; Starr et al. 2005; Matala et al. 2008). Because our models were developed for stands level, geographic location and climatic parameters were not included in the equations. In our study, the effect of factors other than stand features on litterfall could not be determined probably due to complex interactions between the stand, environment and treatment effects. Although most of the variation in litterfall was explained by some stand parameters, the effect of environmental characteristics remained unclear.
Conclusions
Litterfall amount and carbon input were found to increase as the stand growth until the maturity was reached and then remained almost constant in this study. Thinnings reduced litterfall, correspondingly carbon input through litterfall, in mature stands in our study as reported by earlier studies for young stands, while had no significant effect in overmature stands. Seed cutting was found to considerably decrease litterfall amount both in mature and overmature stands, indicating that carbon and nutrient cycling can be altered by silvicultural treatment whereby influencing the litterfall production. Furthermore, seed cut operation caused more decline in carbon input into the soil via litterfall in overmature stands than in mature stands, implying that litterfall of overmature stands was more vulnerable to sever silvicultural treatment than that of mature stands. Therefore, seed cutting should be considered to be planned in preferably mature stands in forest management in order not to reduce carbon input into the soil. Besides, to increase carbon input into the soil, thinning should be applied lighter in mature stands while a bit heavier in overmature stands. Stand basal area, site index, and stand age can be used in estimating the litterfall production of Scots pine stands. In addition, accurate LAI estimation through allometric equations encompassing whole stages of managed Scots pine stand is needed to compare litterfall estimation from various regions.
References
Ågren GI, Knecht MF (2001) Simulation of soil carbon and nutrient development under Pinus sylvestris and Pinus contorta. For Ecol Manag 141:117–129
Albrektson A (1988) Needle litterfall in stands of Pinus sylvestris L. in Sweden, in relation to site quality, stand age and latitude. Scand J For Res 3:333–342
Alemdağ Ş (1967) Structure and yield potential of Scotsch pine (Pinus Sylvestris L.) forests in Turkey and the principles to be followed in managing these forests. Forest Research Institute, Ankara
Alvera B (1980) Retorno el suelo de materia organica y bioelementos minerales en dos Bosques altoaragonoeses. Publ Cent PIR Biol Exp 12:55–62
Ayari A, Zubizarreta-Gerendiain A, Tome M, Tome J, Garchi S, Henchi B (2012) Stand, tree and crown variables affecting cone crop and seed yield of Aleppo pine forests in different bioclimatic regions of Tunisia. For Syst 21(1):128–140
Bārdule A, Lazdiņš A, Bārdulis A, Lazdiņa D, Stola J (2012) Fractions and chemical content of forest litterfall in Scots Pine (Pinus sylvestris L.) stand in level II forest monitoring. Proc LATV Univ Agric 27(322):16–21
Bellingham PJ, Morse CW, Buxton RP, Bonner KI, Mason NWH, Wardle DA (2013) Litterfall, nutrient concentrations and decomposability of litter in a New Zealand temperate montane rain forest. N Z J Ecol 37(2):162–171
Berg B, Laskowsky R (2005) Litter decomposition—a guide to carbon and nutrient turnover. In: Advances in ecological research, vol 38, Elsevier Academic Press, California
Berg B, Albrektson A, Berg MP, Cortina J, Johansson MB, Gallardo A, Madeira M, Pausas J, Kiratz W, Vallejo R, McClaugherty C (1999) Amounts of litterfall in some pine forests in a European transect, in particular Scots pine. Ann For Sci 56:625–639
Bergmann C, Fischer T, Hüttl RF (1998) Seasonal variability of organic matter and N input with litterfall in Scots pine stands. In: Hüttl RF, Bellmann K (eds) Changes of atmospheric chemistry and effects on forest ecosystems. Nutrients in ecosystems, vol 3. Springer, Dordrecht, pp 177–186
Blanco JA, Imbert JB, Castillo FJ (2006) Influence of site characteristics and thinning intensity on litterfall production in two Pinus sylvestris L. forests in the western Pyrenees. Forest Ecol Manag 237:342–352
Çömez A (2010) Determination of carbon sequestration in Scots pine (Pinus sylvestris L.) stands on Sündiken Mountain. Dissertation, Istanbul University
Cousens JE (1988) Report of a twelve-year study of litterfall and productivity in a stand of mature Scots pine. Forestry 61(3):255–266
Diaz-Pines E, Rubio A, Montes F (2011) Aboveground soil C inputs in the ecotone between Scots pine and Pyrenean oak in Sierra de Guadarrama. For Syst 20(3):485–495
Dincă LC, Dincă M, Vasile D, Spârchez C, Holonec L (2015) Calculating organic carbon stock from forest soils. Not Bot Horti Agrobot 43(2):568–575
Erkan N, Comez A, Aydin AC, Denli O, Erkan S (2017) Litterfall in relation to stand parameters and climatic factors in Pinus brutia forests in Turkey. Scand J For Res. https://doi.org/10.1080/02827581.2017.1406135. Accessed 17 Feb 2018
EUFORGEN (2009) Distribution map of Scots pine (Pinus sylvestris). http://www.euforgen.org/species/pinus-sylvestris/. Accessed 23 Aug 2018
Finér L (1996) Variation in the amount and quality of litterfall in a Pinus sylvestris L. stand growing on a bog. For Ecol Manag 80:1–11
Irmak A, Çepel N (1968) Determining litterfall amount and nutrient return to the soil through litterfall in a beach, oak, and pine stands in Belgrad Forest. J Fac For Istanb Univ 17(2A):53–76 (in Turkish)
IUSS Working Group WRB (2006) World reference base for soil resources 2006. World soil resources reports no. 103. FAO, Rome
Jagodziński AM, Kałucka I (2008) Age-related changes in leaf area index of young Scots pine stands. Dendrobiology 59:57–65
Jiménez MN, Navarro FB (2016) Thinning effects on litterfall remaining after 8 years and improved stand resilience in Aleppo pine afforestation (SE Spain). J Environ Manag 169:174–183
Kim C, Son Y, Lee WK, Jeong J, Noh NJ (2009) Influences of forest tending works on carbon distribution and cycling in a Pinus densiflora S. et Z. stand in Korea. For Ecol Manag 257:1420–1426
Koray EŞ (2017) Nutrient inputs by litterfall into ecosystems in Anatolian black pine stands at Turkmendagi. Master of Science Thesis, Istanbul University
Kouki J, Hokkanen T (1992) Long-term needle litterfall of a Scots pine Pinus sylvestris stand: relation to temperature factors. Oecologia 89:176–181
Krannitz PG, Duralia TE (2004) Cone and seed production in Pinus ponderosa: a review. West N Am Natur 62(4):208–218
Kroetsch D, Wang C (2008) Particle size distribution. In: Angers DA, Larney FJ, Carter, Gregorich EG (eds) Section VI soil physical analysis. Soil sampling and methods of analysis. CRC Press, Boca Raton, pp 713–725
Krynytska O, Bondarenko T, Capuliak J, Trenčiansky M (2017) Quantitative analysis of litterfall in hornbeam-oak-pine stands in the Lviv Roztoche region. Cent Eur For J 63:35–42
Kunhamu TK, Kumar BM, Viswanath S (2009) Does thinning affect litterfall, litter decomposition, and associated nutrient release in Acacia mangium stands of Kerala in peninsular India? Can J For Res 39:792–801
Lado-Monserrat L, Lidón A, Bautista I (2016) Erratum to: litterfall, litter decomposition and associated nutrient fluxes in Pinus halepensis: influence of tree removal intensity in a Mediterranean forest. Eur J For Res 135:203–214
Lehtonen A, Sievänen R, Mäkelä A, Mäkipää R, Korhonen KT, Hokkanen T (2004) Potential litterfall of Scots pine branches in southern Finland. Ecol Model 180:305–315
Lehtonen A, Lindholm M, Hokkanen T, Salminen H, Jalkanen R (2008) Testing dependence between growth and needle litterfall in Scots pine—a case study in northern Finland. Tree Physiol 28:1741–1749
Liu C, Westman CJ, Berg B, Kutsch W, Wang GZ, Man R, Ilvesniemi H (2004) Variation in litterfall-climate relationships between coniferous and broadleaf forests in Eurasia. Glob Ecol Biogeogr 13:105–114
Lonsdale WM (1988) Predicting the amount of litterfall in forests of the world. Ann Bot 61:319–324
Lu SW, Liu CP (2012) Patterns of litterfall and nutrient return at different altitudes in evergreen hardwood forests of Central Taiwan. Ann For Sci 69:877–886
Martínez-Alonso C, Valladares F, Camarero JJ, López Arias M, Serrano M, Rodríguez JA (2007) The uncoupling of secondary growth, cone and litter production by intradecadal climatic variability in a Mediterranean Scots pine forest. For Ecol Manag 253:19–29
Matala J, Kellomäki S, Nuutinen T (2008) Litterfall in relation to volume growth of trees: analysis based on literature. Scand J For Res 23(3):194–202
McGeever AH, Mitchell FJG (2016) Re-defining the natural range of Scots Pine (Pinus sylvestris L.): a newly discovered microrefugium in western Ireland. J Biogeogr. https://doi.org/10.1111/jbi.12761. Accessed 24 July 2018
Muukkonen P (2005) Needle biomass turnover rates of Scots pine (Pinus sylvestris L.) derived from the needle-shed dynamics. Trees 19:273–279
Navarro FB, Romero-Freire A, Del Castillo T, Foronda A, Jiménez MN, Ripoll MA, Sánchez-Miranda A, Huntsinger L, Fernández-Ondoño E (2013) Effects of thinning on litterfall were found after years in a Pinus halepensis afforestation area at tree and stand levels. For Ecol Manag 289(1):354–362
Neumann M, Ukonmaanaho L, Johnson J, Benham S, Vesterdal L, Novotný R, Verstraeten A, Lundin L, Thimonier A, Michopoulos P, Hasenauer H (2018) Quantifying carbon and nutrient input from litterfall in european forests using field observations and modeling. Biogeochem Cycles 32:784–798
Novák J, Slodičák M, Dušek D, Kacálek D (2013) Norway spruce litterfall and forest floor in the IUFRO thinning experiment CZ 13—Vítkov. J For Sci 59(3):107–116
Novák J, Dušek D, Slodičák M, Kacálek D (2015) Litterfall in young Scots pine stands. Zprávy Lesnického Výzkumu 60(2):115–121
OGM (2015) Forest resources in Turkey (in Turkish). Published by Republic of Turkey General Directorate of Forestry
Ordóñez JL, Retana J, Espelta JM (2005) Effects of tree size, crown damage, and tree location on post-fire survival and cone production of Pinus nigra trees. For Ecol Manag 206:109–117
Paavilainen E (1987) Effect of fertilization on the litterfall of Pinus sylvestris and Betula pubescens on drained peatlands. Scand J For Res 2:59–75
Palosuo T, Foereid B, Svensson M, Shurpali N, Lehtonen A, Herbst M, Linkosalo T, Ortiz C, Todorovic GR, Marcinkonis S, Li C, Jandl R (2012) A multi-model comparison of soil carbon assessment of a coniferous forest stand. Environ Model Softw 35:38–49
Pausas JG (1997) Litterfall and litter decomposition in Pinus sylvestris forests of the eastern Pyrenees. J Veg Sci 8:643–650
Portillo-Estrada M, Korhonen JFJ, Pihlatie M, Pumpanen J, Frumau AKF, Morillas L, Tosens T, Niinemets Ü (2013) Inter- and intra-annual variations in canopy fine litterfall and carbon and nitrogen inputs to the forest floor in two European coniferous forests. Ann For Sci. https://doi.org/10.1007/s13595-013-0273-0. Accessed 17 Feb 2018
Ravindranath NH, Ostwald M (2008) Carbon inventory methods: handbook for greenhouse gas inventory, carbon mitigation, and roundwood production projects. In: Advances in global change research, vol 29, Springer, Berlin
Roig S, del Río M, Cañellas I, Montero G (2005) Litterfall in Mediterranean Pinus pinaster Ait. stands under different thinning regimes. For Ecol Manag 206:179–190
Santa Regina I, Gallardo JF (1995) Biogeochemical cycles in forests of the “Sierra de Béjar” mountains (province of Salamanca, Spain): decomposition index of the leaf litter. Ann Sci For 52:393–399
Santa Regina I, Tarazona T (2000) Nutrient return to the soil through litterfall and throughfall under beech and pine stands of Sierra dela Demanda, Spain. Arid Soil Res Rehabil 14:239–252
Segura C, Fernández-Ondoño E, Jiménez MN, Navarro FB (2017) Thinning affects the needlefall nutrient return to soil in a semiarid Aleppo pine afforestation while the nutrient dynamics remain unchanged. For Ecol Manag 405:257–270
Slodičák M, Novák J (2006) Litter-fall as a source of nutrients in Scots pine Stands with different thinning regime. Conference on stabilization of the forest functions in biotopes disturbed by anthropogenic activity under changing ecological conditions. Opočno 5. - 6. 9. Antonín Jurásek, Jiří Novák, Marian Slodičák. Jíloviště-Strnady (eds) Výzkumný ústav lesního hospodářství a myslivosti—Výzkumná stanice
Smith DM, Larson BC, Kelty MJ, Ashton PMS (1997) The practise of silviculture—applied forest ecology. Wiley, New York
Starr M, Saarsalmi A, Hokkanen T, Merilä P, Helmisaari HS (2005) Models of litterfall production for Scots pine (Pinus sylvestris L.) in Finland using stand, site and climate factors. For Ecol Manag 205:215–225
Tobin B, Black K, Osborne B, Reidy B, Bolger T, Nieuwenhuis M (2006) Assessment of allometric algorithms for estimating leaf biomass, leaf area index and litterfall in different-aged Sitka spruce forests. Forestry 79(4):453–465
Ťupek B, Mäkipää R, Heikkinen J, Peltoniemi M, Ukonmaanaho L, Hokkanen T, Nöjd P, Nevalainen S, Lindgren M, Lehtonen A (2015) Foliar turnover rates in Finland—comparing estimates from needle-cohort and litterfall-biomass methods. Boreal Environ Res 20:283–304
Turner MG, Turner DM, Romme WH, Tinker DB (2007) Cone production in young post-fire Pinus contorta stands in Greater Yellowstone (USA). For Ecol Manag 242:119–126
Ukonmaanaho L, Merilä P, Nöjd P, Nieminen TM (2008) Litterfall production nutrient return to the forest floor in Scots pine and Norway spruce stands in Finland. Boreal Environ Res 13(suppl. B):67–91
Wutzler T, Mund M (2007) Modelling mean above and below ground litter production based on yield tables. Silva Fenn 41(3):559–574
Yang Y, Guo J, Chen G, Xie J, Cai L, Lin P (2004) Litterfall, nutrient return, and leaf- litter decomposition in four plantations compared with a natural forest in subtropical China. Ann For Sci 61(5):465–476
Acknowledgements
This study was supported financially by the Turkish General Directorate of Forestry (project number ESK-09. (6303)/2010-2016). We thank Dr Osman Yalçın Yılmaz (Istanbul University-Cerrahpasa, Faculty of Forestry) for preparing the map of the study area.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Lluís Coll.
Rights and permissions
About this article

Cite this article
Çömez, A., Tolunay, D. & Güner, Ş.T. Litterfall and the effects of thinning and seed cutting on carbon input into the soil in Scots pine stands in Turkey. Eur J Forest Res 138, 1–14 (2019). https://doi.org/10.1007/s10342-018-1148-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-018-1148-6