
Abstract
The optimization of forest management under climate change uncertainty requires a comparison of many alternative management options under different climate scenarios and the use of stochastic and adaptive approaches. Empirical growth and yield models are highly suitable for this, provided they include sensitivity to environmental influences. Here, we present a climate-sensitive empirical growth and yield model that is based on the direct integration of environmental effects in dynamic growth and survival functions, which allows for the evaluation of changing site conditions over time. Individual-tree diameter and height growth and the probability of a tree to survive any 5-year period were modelled for even-aged beech (Fagus sylvatica) stands in Switzerland using a distance-independent approach. Changing site conditions were based on a drought index (locally adjusted water balance) and sum of degree-days. The data for fitting the model were taken from 30 permanent yield plots repeatedly measured from 1930 to 2010. Reasonable results were obtained in the model evaluation: (1) validation against independent National Forest Inventory data indicated that the incorporation of drought and sum of degree-days in the model was appropriate; (2) accurate simulations over around 50 years of past stand development were achieved (for changes in basal area over 5-year measurements in all plots, the bias was 3 % and the root mean square error 32 %); and (3) the impact of climate change may vary considerably along the range of current site conditions. We thus conclude that the model can be used in management planning under climate change uncertainty.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Forest management planning under climate change requires a comparison of many alternative management options under different climate scenarios and their estimated impacts on ecosystem goods and services. Management options are typically evaluated every 10 years along the considered simulation period, taking into account abiotic and biotic risks. The simulation results define the decision space that is required by the planning process, in which a planning model composed of a set of functions integrating the main objectives and preferences of the decision makers is solved with optimization algorithms (Pukkala 2002). Thus, a number of candidate plans at the stand or at the forest level and contingent on climate change are obtained. Uncertainty on climate change and risks (e.g. storms, fire) can be accommodated using stochastic and adaptive approaches (cf. Jacobsen and Thorsen 2003; González et al. 2005; Pukkala and Kellomäki 2012; Yousefpour et al. 2014).
Process-based models that explicitly integrate physiological processes such as photosynthesis, transpiration and respiration (cf. Gracia et al. 1999; Pietsch et al. 2005) are highly versatile tools to assess climate change effects on stand development, because of the direct link to changes in growth conditions. However, these models often require calibration on empirical data, which can be quite difficult in a climate setting because they employ many parameters for which data are not always available (Fontes et al. 2010). In addition, computational effort is high for these models, and many optimization routines used in advanced management planning under climate change uncertainty (e.g. those related to dynamic programming; cf. Jacobsen and Thorsen 2003) may not be feasible.
An alternative approach is to use simpler models based on a few variables that are suitable for optimization of forest management. Empirical growth and yield models (i.e., statistical models based on stand dynamics data from inventory plots or long-term trials; cf. Vanclay 1994; Pretzsch 2009) have a long tradition in forest science, but they are typically not sensitive to environmental conditions other than “site index” or similar concepts, which are treated as site-specific constants. However, this type of model could be further developed to explicitly consider environmental sensitivity. These models are based on few predictor variables that are often available from standard forest inventories. They further provide accurate yield predictions and are sensitive to management effects.
In this study, we present a climate-sensitive empirical growth and yield model that is based on the direct integration of environmental effects in dynamic individual-tree growth and survival functions. This allows for the direct prediction of stand development related to environmental variables, both static variables (e.g. topography, soil) and dynamic variables (e.g. climate). Two conditions need to be met for this approach: (1) permanent plot data on stand dynamics and data on environmental variables (primarily climate and soil) are combined at the same temporal and spatial resolution; (2) these data cover a wide range of climatic and soil conditions as well as management (e.g. thinning intensity and type) for the targeted species and region, such that the model can be used with confidence to extrapolate tree growth–site relationships under scenarios of climatic change.
Other approaches for integrating climate sensitivity in empirical growth and yield models have been developed in recent years. Among these are the use of signal-transfer functions (Baldwin et al. 2001; Matala et al. 2005; Kellömaki et al. 2015), the derivation of summary models on the basis of environmental variables and simplified process model structures (Härkönen et al. 2010), the prediction of site index based on environmental variables available in National Forest Inventory data (Seynave et al. 2005, 2008) and the direct use of environmental variables in dynamic equations describing stand-level growth (González-García et al. 2015; Sharma et al. 2015). Compared with the three first approaches, the main advantage of the model presented in this study relies on the direct integration of environmental effects as explanatory variables. This prevents the use of additional intermediate models, be it a single equation or a whole set, for predicting an indicator of site productivity (e.g. site index), and thus avoids incorporating the error components of those models in simulations. The use of signal-transfer functions (Baldwin et al. 2001; Matala et al. 2005; Kellömaki et al. 2015) may be taken as an example for illustrating this. In this approach, a process-based model and an empirical growth and yield model are combined. The development process works as follows: (1) a variable of site productivity available in both models is selected, (2) the effects of the selected environmental changes (e.g. temperature, precipitation, CO2 or soil conditions) on the selected indicator are simulated for a broad array of conditions using the process-based model, and (3) simulation results are used to calibrate the signal-transfer functions. In this case, error propagation occurs at least at two levels: (i) the error inherent to the process-based model used to calculate the required data for fitting the function(s) and (ii) the error component of the fitted functions for predicting the used site index in the empirical growth and yield model. Regarding the direct use of environmental variables in dynamic stand-level equations, the individual-tree-level approach presented in this study is analogous in preventing error propagation, but it incorporates additional desirable features for forest management planning such as flexible simulation of many types of cuttings and more detailed illustration of the stand.
The aim of this study was to develop a climate-sensitive empirical growth and yield model for even-aged stands that (1) allows the direct integration of explicit and biologically consistent environmental effects, (2) can be used with variables normally available in practical management planning and (3) is sensitive to management effects. The model was developed at the individual-tree distance-independent level (cf. Palahí et al. 2003; Trasobares et al. 2004) and considers individual-tree diameter and height growth, and the probability of a tree to survive any 5-year period. Even-aged beech (Fagus sylvatica L.) stands in Switzerland, which are one of the main forest types in the country occupying 18.3 % of the forest area (Brändli 2010), were used as a case study, taking advantage of the high-quality data available. The model should be computationally efficient to allow the combined use with optimization techniques in advanced forest management planning.
Materials and methods
Data
Permanent plot data
The data for model development were taken from permanent yield plots repeatedly measured from 1930 to 2010 by the Swiss Federal Institute for Forest, Snow and Landscape Research WSL (Birmensdorf). A total of 30 permanent yield plots distributed all over Switzerland were used (Fig. 1). They were selected to represent even-aged beech (Fagus sylvatica L.) stands featuring >70 % of basal area by this species. The sites cover a wide range of environmental conditions (climate, water holding capacity, slope, aspect; these being implicitly represented in site index, calculated using Eq. A2 in the Appendix), development stage and stand density (Table 1). These data represent stands which had been naturally regenerated by shelterwood systems and were exposed to a broad range of management interventions in terms of thinning type and intensity (natural thinning; moderate, heavy and very heavy thinning from below; thinning from above; for details cf. Schütz and Zingg 2010).

Geographical distribution of sample plots used for model development and evaluation representing even-aged stands dominated by beech (at least 70 % in stand basal area) in Switzerland; © swisstopo (JD100042). Note that because some of the modelling plots are so close to each other not all modelling plots (a total of 30) can be differentiated on the map
The measurements were taken every 4–7 years, immediately after a thinning intervention. Plot size generally is about 0.25 ha, and all trees are identified by a number. At each measurement, dbh (diameter at breast height) of all trees >4 cm was measured crosswise with a calliper to the mm, and in some cases for young stands dbh > 2 cm was recorded. Measurement height and locations are permanently marked on the tree. The heights of a sample of 20–40 trees per site were measured with an accuracy of about 0.8 m (Schütz and Zingg 2010). Each tree observed as living in the previous measurement was identified in each following measurement as standing, dead or thinned. Based on all standing beech trees in all plots, measured along several consecutive periods, a total of 25,611 diameter growth observations and 3900 height growth observations were available. From the standing dead trees, which were used for survival modelling, 16,883 observations were available (Table 2). Because it was not known whether trees removed in a thinning were alive or dead at the time of thinning, the thinned trees were not used as observations in survival modelling. For each measurement, the characteristics of the growing stock were computed from the individual-tree measurements of the plots.
The data for validating the diameter growth model were taken from the permanent plots of the Swiss National Forest Inventory (SNFI) (WSL 2010), which consists of a systematic sample of plots distributed on a square grid of 1.4 km mesh width, with a 10-year re-measurement interval. From the inventory plots over the whole of Switzerland, all plots classified as even-aged were selected in which soil measurements were available (i.e. ICP Forests Level I and Swiss Sanasilva plots, Webster et al. 1996) and the proportion of beech basal area was at least 70 %. This resulted in 50 plots that due to the systematic sampling design provided an appropriate representation of the range of environmental conditions, development stage and stand density (Fig. 1; Table 1). The sample plots were measured in 1982–1986, 1993–1995 and 2004–2006. A circular plot of 0.02 ha was used for all trees with dbh ≥ 12 cm, while a 0.05 ha circular plot was used for trees with dbh ≥ 36 cm. Dbh was measured one time (towards the plot centre) from all sampled trees to cm resolution; measurement height and direction were permanently marked on the tree. In total, this sample provided 360 diameter increment observations (Table 2). In addition, the diameter growth observations from one plot (Neunkirch, canton of Schaffhausen) of the ICP Forests Level II network (Swiss Long-term Forest Ecosystem network maintained by WSL, Dobbertin 2005), growing under severe drought conditions (Table 1) were compared graphically to the fitted diameter growth model.
Soil water holding capacity
Water holding capacity (WHC) is defined as the plant available water storage capacity of the soil between field capacity (pF = 1.8) and permanent wilting point (pF = 4.2). In soils without limits for the rooting system, WHC was calculated for 0–100 cm depth, including the organic layer where present. Where rooting depth was limited to <100 cm by parent rock or permanent anaerobic conditions, WHC was calculated only to rooting depth. WHC was estimated for each soil horizon using pedotransfer functions (PTF). In mineral soil horizons, WHC was estimated according to Teepe et al. (2003); input parameters for the PTF are fine earth density (five classes), texture (ten classes) and humus content. In organic horizons, the method proposed by Zuber (2007) was used; in F and H horizons, a WHC of 27.4 and 35.7 v % was always used, respectively. Finally, the WHC of each horizon had to be reduced proportionally to take the respective stone content into account and was then summed up to 100 cm depth (Table 1).
For the WSL permanent yield plots used for modelling, only soil parameters estimated from soil pit information were available. For the Level I plots (SNFI data used for the validation of the diameter increment model) and the Level II plot in Neunkirch, data on texture and humus content were available as measured values, but not for rooting depth, soil density and stone content, which were estimated from the soil profiles. Texture was measured with the sedimentation method according to Gee and Bauder (1986) and organic carbon (Corg) content by dry combustion (Walthert et al. 2010).
Climate data
For developing the model, we obtained daily data of mean temperature and precipitation sum from the database of the Land Use Dynamics Research Group at WSL. The data are based on daily climate maps of Switzerland in a 100-m resolution for the years 1930–2006. The maps were produced using the interpolation software “DAYMET” by Thornton et al. (1997), which uses daily minimum, maximum, mean temperature and precipitation sums of all available MeteoSwiss climate stations and a 100-m digital elevation model as input. For calculating the climatic values for each growing period in a plot, we averaged the data over a 3 × 3 window of grid cells centred around the respective plot (Didion et al. 2009). Daily data for mean temperature were used to calculate monthly averages, which were then used to calculate monthly potential evapotranspiration (PET) according to Thornthwaite and Mather (1957) and the monthly sum of degree-days (using a 5 °C threshold). Monthly precipitation sum was also calculated. For each growth period registered in a permanent plot, the precipitation sum, the sum of degree-days and PET values were calculated as follows: first monthly values were calculated for the period between 1 year prior to the initial measurement and the year of the re-measurement; second, the values for that period were calculated by adding the monthly values included in the vegetation period assumed for the species in the region (April–October). The year prior to the initial year was added to account for potential lag effects due to extreme climatic conditions. The models were also fitted using 2- and 3-year lag periods, but fitting was slightly better using a 1-year period.
Climate scenario data
The climate scenario data used for assessing the behaviour of the model in applications were provided by the Research Unit Landscape Dynamics of the WSL. Daily mean temperature and precipitation sum for a set of sites selected from the modelling and the Level II validation samples to represent the measured range in terms of drought and temperature sum were available from DAYMET daily climate maps of Switzerland (100-m resolution; period 1961–2000 for current climate) and from three regional circulation model simulations of the IPCC AR4 A1B emission scenario from 2001 to 2100, hereafter referred to as “climate scenarios” (Table 3). These climate scenarios were selected to include climate change uncertainty in the simulations. The regional climate scenario data were processed and downscaled to plot level by the Research Unit Landscape Dynamics of the WSL. Absolute mean temperature and precipitation sum values of a scenario day were calculated as follows for each plot: (1) monthly averages for the period 1961–2000 were calculated; (2) each climate scenario was ran to yield daily predictions for the period 1961–2100 and monthly averages of those predictions calculated for the period 1961–2000; (3) a daily anomaly was defined as the difference between an absolute daily prediction and the monthly average (1961–2000) of the predictions; and (4) the absolute values of a scenario day were calculated by adding the anomalies to the corresponding monthly average of the reference period. The anomalies were all first bilinearly interpolated to 10" (coordinate system: WGS84), and then, the value at the requested coordinates was extracted by another bilinear interpolation. For each stand, the precipitation sum, PET and the sum of degree-days for every 5-year growth period from 2010 to 2100 were calculated exactly in the same way as the climatic variables used for fitting the model, also considering a 1-year lag period.
Model development
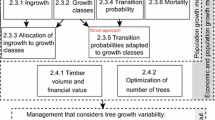
Individual-tree modelling of 5-year diameter growth, height growth and survival probability was based on tree size, competition and site effects (c.f. Vanclay 1994; Palahí et al. 2003; Trasobares et al. 2004; Pretzsch 2009). Both statistical criteria and existing knowledge on biological processes were considered to select the best-fitting candidate models, searching iteratively for an optimal combination of explanatory variables representing tree size, competition and site effects in each model. For representing local site and dynamic climate effects in the growth and survival models various climatic, edaphic and topographic variables were initially considered (see “Modelling of site-specific climate effects” section). Finally, a drought index and a degree-day index were developed (Fig. 2). Local site effects were incorporated by using soil water holding capacity, while changing environmental conditions were incorporated by the use of potential evapotranspiration, precipitation and the sum of degree-days (heat sum above 5 °C).

Schematic drawing of the model and its functioning in simulations. The model, designed for 5-year projections and based on tree size, competition and site effects, considers individual-tree diameter and height growth, and the probability of a tree to survive. The initial stand can be represented by its diameter class distribution or diameter distribution of measured individual trees
A model for dominant height growth was also developed (c.f. Appendix) for two main reasons: (1) to allow site evaluation following the classical site index approach (dominant height at a given reference age) and compare its performance with the developed drought and degree-day indices; (2) to provide a reliable dominant height growth model for evaluating the behaviour of the individual-tree height growth model in long-term simulations.
The independent individual-tree diameter growth and height growth models (i.e. height growth is not calculated as a function of diameter growth of projected tree diameter) allow sensitivity to changes in stem height/diameter ratios, which is relevant for forest management. Environmental effects on survival probability were integrated using the stand drought index in the first 5-year simulation, while more accurate predictions can be obtained by using (calculated) actual growth from the second 5-year simulation period onwards.
The model, designed for 5-year projections, allows simulating the development of any even-aged beech stand in Switzerland (Fig. 2). The initial stand can be represented by its diameter class distribution or diameter distribution of measured individual trees; a static height model is used for calculating tree heights (cf. Appendix). If the initial stand is simulated using data measured in plots of size significantly different from 0.25 ha, approximate size of the plots used for fitting the model, this may have an impact on the ability of the model to capture within-stand variation. In that case the calibration of model predictions should be considered (see e.g. Salas-González et al. 2001). The current model version does not include a regeneration model for simulating establishment. For simulating more than one rotation, a suitable regeneration model or a typical initial stand (adjusted to site conditions) can be used. A more detailed description of the simulation of one 5-year time step can be found in the Appendix.
Modelling of site-specific climate effects
Site effects were incorporated using climatic, edaphic and topographic variables. After exhaustive preliminary analyses of fitted models using various climatic (mean temperature, precipitation, potential evapotranspiration, sum of degree-days, etc.), edaphic (soil depth, water holding capacity) and topographic (mean slope, aspect) explanatory variables and multiple interactions and transformations of them, site effects were integrated by combining a drought index (Eq. 1) and an index derived from the sum of degree-days (Eq. 2):
where \(WB = PREC/PET \ if \,\frac{PREC}{PET} \le 1 \,else \ WB = 1\) and \(WHC = WHC_{m} /210 \ if \,\frac{{WHC_{m} }}{210} \le 1 \,else \ WHC = 1\). DI is a stand drought index in the range]0,1], A is a weighting scalar that maximizes the fitting of the diameter growth and height growth models, WB is the climatic water balance index in the range]0,1], PREC is the precipitation sum (April–October; mm), PET is potential evapotranspiration (April–October; mm), WHC is the plant available soil water holding capacity index in the range ]0,1] and WHC m is soil water holding capacity (for 1 m of soil depth; mm);
where GDDI is the degree-day index in the range]0,1] and GDD is the sum of degree-days (April–October).
For the finally selected climatic and edaphic variables, both indices included threshold values which were determined by comparing the model fits based on several values in the proximity of the visual threshold and selecting the best-fitting model: (1) PREC/PET values ≥1 mean that site productivity is not limited by the climatic water balance; (2) WHCm values ≥210 mm mean that site productivity is not limited by WHC; and (3) a sum of degree-days ≥1900 (for the period April–October) means that site productivity is not limited by the temperature sum. DI and GDDI values equal to one mean no limitation by water or temperature sum, respectively, while values around 0.7 (or even less) represent limitations by extreme drought or low temperature sum. Optimal or close-to-optimal site conditions for beech growth in the region are reached when DI and GDDI simultaneously adopt a value of “1.”
Modelling of diameter and height increment
The total height measurements of a sub-sample of 20–40 remeasured trees at each plot did not show systematic errors. Thus, dbh–height curves fitted in successive measurements of the same plot showed expected evolution and the height increment relationships adopted the typical unimodal shape of tree growth processes that allowed the development of height growth models. The log-transformed 5-year growth of both diameter and height was modelled at the level of the individual tree using linear regression, which resulted in multiplicative models (Flewelling and Pienaar 1981; Wykoff 1990). Five-year diameter growth and height growth were calculated as the difference between two consecutive diameter and height measurements, respectively. The resulting diameter and height growth observations (4- to 7-year growth) were linearly scaled to 5-year periods, according to the number of vegetation periods between two measurements. The predictors were chosen from tree, stand and site characteristics as well as their transformations. Different models were compared searching for an optimal combination of explanatory variables and their transformations representing tree size, competition and site effects. The significance level was set to 5 %, and the residual diagnostics were checked for all models. Due to the hierarchical structure of the data (i.e. there were several observations from the same trees and trees were grouped into plots), the generalized least-squares (GLS) technique was applied to fit mixed-effects linear models (Goldstein 1996) using the maximum likelihood procedure PROC MIXED in the software SAS/STAT (2011). The diameter growth (Eq. 3) and height growth (Eq. 4) models for beech were as follows:
where id5 is diameter growth (cm in 5 years); ih5 is height growth (m in 5 years); dbh is diameter at breast height at the beginning of the period (cm); h is tree height at the beginning of the period (m); BAL_rem is the total basal area of trees larger than the subject tree remaining after thinning (beginning of the period) (m2 ha−1); G_rem is stand basal area of trees remaining after thinning at the beginning of the period; BAL_thin is the total basal area of trees larger than the subject tree thinned at the beginning of the next 5-year period (m2 ha−1); DI is the drought index for the next 5-year period; and GDDI is the degree-day index for the next 5-year period. Subscripts l, k and t refer to plot l, tree k and measurement t, respectively. u l ,u lt , u lk and e lkt are independent and identically (normally) distributed between-plot, between-measurement, between-tree and within-tree random effects with a mean of 0 and constant variances of \(\sigma_{\text{plot}}^{2}\), \(\sigma_{\text{meas}}^{2}\),\(\sigma_{\text{tree}}^{2}\) and \(\sigma_{\text{e}}^{2}\), respectively. These variances and the parameters β i were estimated using the GLS method. At first, the three random effects u l , u lt and u lk were included in both models, but since the between-plot random effect was not significant in the diameter growth model and the between-plot and between-tree random effects were not significant in the height growth model, these random effects were therefore excluded. Other competition indices such as modified basal area of larger trees proposed by Schröder and Gadow (1999) were examined but the model using the above covariates capturing competition performed better.
To estimate the scalar value A of the drought index (Eq. 1), 10 equally spaced values in the range [0.1–1] were examined at first, i.e. both models (Eqs. 3 and 4) were fitted with each candidate value using all plots until fitting was maximized. The optimal A value was 0.8 for both models; for determining the final value, ten values around this number (0.75, 0.76, 0.77, 0.78, 0.79, 0.81, 0.82, 0.83, 0.84 and 0.85) were tried. To convert the logarithmic predictions of Eqs. 3 and 4 to the arithmetic scale, an empirical ratio estimator for bias correction in logarithmic regression was applied to Eqs. 3 and 4. Following Snowdon (1991), the proportional bias in logarithmic regression was estimated from the ratio of mean diameter growth \(\left( {\overline{\iota d5} } \right)\) (Eq. 3) and mean height growth \(\left( {\overline{\iota h5} } \right)\) (Eq. 4) to the mean of the back-transformed predicted values from the regression \(\exp \left[ {\ln \;\hat{i}d5} \right]\) and \(\exp \left[ {\ln \;\hat{i}h5} \right]\), respectively. To correct for the exclusion of negative growth observations (2 and 4 % of the observations in diameter growth and height growth, respectively) in logarithmic regression, which may lead to biased predictions, the mean diameter growth \(\left( {\overline{\iota d5} } \right)\) and mean height growth \(\left( {\overline{\iota h5} } \right)\) values corresponded to the whole of the sample (i.e. without excluding negative growth observations).
Collinearity statistics were obtained, also for the combinations of variables used in survival modelling, to detect the presence and severity of multicollinearity. The eigenvalues of the correlation matrix for the standardized explanatory variables were arranged from the largest to the smallest, and the square root of the ratio of the largest to smallest eigenvalue, the condition index, was calculated. Multicollinearity was evaluated when a component associated with a condition index greater than 30 contributed strongly to the variance of two or more variables (Belsley et al. 1980).
Modelling of tree survival
In the data analysis, two types of mortality were distinguished: background mortality (related to stand density and structure) versus disturbance-induced mortality (storms or insect attack). Because the objective was to predict background mortality, data from plots that had been affected by disturbance-induced mortality were not used in model fitting. Various authors have demonstrated that growth is an important explanatory variable for mortality and that actual growth is more relevant than predicted growth for estimating mortality (Monserud 1976; Waring 1983; Bigler and Bugmann 2004). Actual growth, however, is often not available from initial state inventory data. Therefore, two different models were fitted (Eqs. 5 and 6) using the binary logistic procedure in SPSS (2010). They can be used depending on the information available during the period of interest in simulations. Equation 5 may be used in the initial simulation step, while Eq. 6, which provides more accurate predictions by linking individual-tree survival to environmental changes and competition, may be used from the second step onwards, when past growth values are available.
where P(surv) is the probability of a tree surviving the next 5-year growth period and pid5 is past diameter increment (cm in 5 years).
Model evaluation
The models were evaluated quantitatively using the data for model development as well as the independent sample of the SNFI. The data for model fitting were used for calculating the residuals of the models and to examine its distribution, for all possible combinations of variables included in each model. Furthermore, the independent diameter increment sample of the SNFI was used for calculating the residuals of the diameter increment model and examining its distribution for all combinations of variables. The aim was to detect dependencies or patterns that indicate systematic discrepancies. To determine the accuracy of model predictions, the bias and the root mean square error (RMSE) were calculated for the models and also for the independent SNFI diameter increment sample. The relative bias (bias %) and RMSE (RMSE %) were calculated by dividing the absolute values by the mean of the model predictions.
In addition, the models were further evaluated by graphical comparisons between measured stand development, simulated stand development (under current climate), and predictions using other models.
The following simulation tests were carried out to explore model behaviour: (1) the development of five plots selected along the range of site and management was simulated using current climate and compared to measured stand development and predictions using the site index model in the Appendix and the self-thinning model by Schütz and Zingg (2010), (2) the simulated 5-year increment of basal area after thinning (calculated using Eqs. 3 and 6) of all growth intervals of all plots was compared to the measured increments.
Model application
The applicability of the model such as in applied management planning or in research was assessed by the simulation of stand development under alternative climate change scenarios and a typical management scenario. The development of a typical 20-year-old stand (real stand selected from the modelling data; cohort of 2367 trees ha−1 resulting from a 20-year period of natural regeneration in a shelterwood system and a tending intervention) at four sites selected from the WSL plots and Level II plots along the site range of beech in Switzerland was simulated under four climate scenarios (Table 3) and a typical shelterwood management schedule (two thinning and a 20-year regeneration period with two regeneration cuts). The diameter distribution of the selected 20-year stand was used for all four sites. Because this stand corresponded to a productive site, to use realistic initial stand conditions (at 20 years of age) in medium and poor productivity sites (according to current climate conditions) the same diameter distribution was used but the number of trees in each diameter class was multiplied by 0.85 at medium sites and 0.7 at poor sites. This adjustment was based on previous analyses of available data from unmanaged plots in the WSL permanent yield plots. Tree volumes were calculated using a formula provided by the Forest Resources and Management research unit at WSL, which can be found in the Appendix.
Results
Diameter growth model and height growth model
Parameter estimates of the diameter growth model (Eq. 3) and the height growth model (Eq. 4) were significant at the 0.5 % level (Table 4). The shape of the relationships dbh-to-diameter growth (Eq. 3) and height-to-height growth (Eq. 4) adopts the typical unimodal shape of tree growth processes. Increasing competition [BAL_rem and G_rem in diameter growth (Eq. 3); BAL_rem in height growth (Eq. 4)] resulted in decreasing diameter growth and height growth. Under a given competition level (i.e. a given BAL_rem value in Eq. 4), increasing dbh increased height growth. The thinned competition (BAL_thin in Eqs. 3 and 4) decreased diameter growth and increased height growth up to an early threshold value (around 1 m2 ha−1) followed by decreased height growth.
Growht increased with increasing stand drought index and increasing stand degree-day index (Eqs. 3 and 4; Fig. 3), which is also reflected in the SNFI validation data. The weight for the drought index (A in Eq. 1) that maximized the fitting of the diameter growth and height growth models was 0.78. The multiplicative ratio estimator for bias correction in logarithmic regression was 1.0969 for the diameter growth model (Eq. 3) and 1.0886 for the height growth model (Eq. 4).

Multicollinearity was not present in the diameter increment and height increment models. The largest condition index was never >10, despite the used transformations of dbh and BAL_thin to define unimodal patterns in the diameter growth and height growth models (Eqs. 3 and 4).
Survival models
The model using past growth (Eq. 6) provided more accurate estimates of the measured individual-tree survival probabilities (see χ 2 and −2 log-likelihood values in Table 5; both models were fitted using the same sample) than the model omitting this variable (Eq. 5). For both models, the Wald test showed significant parameter estimates (P < 0.005) (Table 5). By analysing Eqs. 5 and 6, it can be deduced that: (1) the greater the past 5-year diameter growth, the greater the probability of a tree surviving (Fig. 4) (Eq. 6); (2) the greater the basal area of trees larger than the subject tree, the smaller the survival probability (Eq. 6); (3) the greater the ratio of the basal area of trees larger than the subject tree to tree dbh, the smaller the survival probability (Eq. 5); and (4) the greater the stand drought index, the greater the probability of a tree surviving (Eq. 5). Figure 4 shows for a mean tree growing under average competition (BAL_rem in Eq. 6), how decreasing past 5-year diameter increment from 1.1 cm to 0.1 cm (e.g. due to abrupt drought stress) decreases the survival probability from 0.9934 to 0.8993.

Expected survival probabilities of an average beech tree as a function of past diameter increment (Eq. 6) plotted with the measured survival probabilities. The survival probabilities were calculated as the average of trees within a given past growth (cm) interval (<0.5; 0.5–0.9; 1–1.9; 2–2.9; ≥3) with a value of “0” used for dead trees and a value of “1” for surviving trees. A mean value is given to BAL_rem
Model evaluation
The residuals of the diameter growth model and the height growth model showed no trends when displayed as a function of predictors or predicted growth. Due to the estimators used for correction of bias, the absolute and relative biases of the diameter growth and height growth models were zero. The absolute and relative RMSE values were 0.68 cm/5a and 50.8 % for the diameter growth model and 0.71 m/5a and 59.6 % for the height growth model, respectively.
The residuals of the diameter growth model calculated using the independent SNFI observations showed no trends when displayed against stand drought index (DI in Eq. 3) (Fig. 5), stand degree-day index (GDDI in Eq. 3), stand water holding capacity index (WHC in Eq. 1), individual-tree dbh (dbh in Eq. 3) and thinned competition (BAL_thin in Eq. 3). Trends were neither found with stand mean slope nor potential site predictor tested in model development. The residuals showed some trend when displayed against competition (BAL_rem and G_rem in Eq. 3) and predicted diameter growth (id5 in Eq. 3). The bias, bias %, RMSE and the RMSE % calculated using the independent SNFI data were 0.12 cm/5a, 8 %, 1.2 cm/5a and 77.4 %, respectively.

Mean bias/residual (in antilog scale; calculated using the independent sample of the SNFI) of the diameter growth model (Eq. 3) as a function of drought index (DI), degree-day index (GDDI), water holding capacity index (WHC; Eq. 1), dbh, total basal area of thinned larger trees at the beginning of the growth period (BAL_thin), total basal area of larger trees remaining after thinning (BAL_rem), total basal area of trees remaining after thinning (G_rem) and predicted diameter growth (predicted id). The thin lines indicate the standard error of the mean
The comparisons of measured versus simulated stand development showed that the model allows accurate long-term simulation of stand development along the range of sites (climate, soil) and management (thinning intensity and type) of beech stands in Switzerland. Figure 6 shows actual and simulated stand development for an unmanaged stand (using Eq. A1 and Eqs. 3, 4, 5 and 6). Despite mortality related to disturbances, which is reflected in measured stand development, the model depicts well the mean trend in stand development (G and N in Fig. 6A, B) and predicts an asymptote in stand basal area. The model also provides accurate prediction of the development in dominant height, which was obtained using the individual-tree height growth model (Eq. 4) and was in line with Eq. A2 in the Appendix (Fig. 6C). Moreover, the individual-tree survival model (Eq. 6) permits accurate prediction of stand-level density-dependent mortality: (1) the predicted evolution of N as a function of Dg resembles closely the measured stand development and the self-thinning limit (Nmax) obtained by applying the self-thinning model from Schütz and Zingg (2010) (Fig. 6D); and (2) the simulated development of G as a function of Dg (Fig. 6E) is also in line with the trend defined by the available basal area measurements in unmanaged stands.

Measured and simulated stand development of an unmanaged plot in a fertile site. Under current climate, drought index is 0.99, degree-day index is 0.96, and site index (calculated using Eq. A2 in the Appendix) is 27 m. G is basal area, N is number of trees per hectare, Hdom is dominant height, and Dg is quadratic mean diameter. Hdom and N (N_max) are also plotted using the site index model and the self-thinning model by Schütz and Zingg (2010), which provides maximum N [N_max] as a function of Dg, respectively. In addition, all basal area measurements in the unthinned plots of the modelling sample are plotted versus simulated stand development to show how the model resembles maximum stand basal area as a function of Dg (E)
The model allows accurate predictions of stand development in managed stands (using Eq. A1 and Eqs. 3, 4, 5 and 6), for four plots selected to represent the range of sites, age, and thinning intensities (Fig. 7). Plotting of the measured vs. predicted stand basal area after thinning (Eqs. 3 and 6) for all plots in all the measurements showed little bias in the predictions (bias 3 %). The predicted range of variation was similar to the observed change. The obtained RMSE was 32 %.

Measured and simulated stand development (under current climate) of four representative managed sample plots along the site amplitude and thinning intensity of beech stands in Switzerland. Plots managed according to A heavy thinning from below, DI = 0.99, GDDI = 0.96 and site index (SI) = 27 m (calculated using Eq. A2 in the Appendix); B very heavy thinning from below, DI = 0.91, GDDI = 0.96 and SI = 18 m; C weak thinning from below, DI = 0.87, GDDI = 1 and SI = 16.5 m; and D weak thinning from below, DI = 1, GDDI = 0.78 and SI = 16 m. G is basal area and Hdom is dominant height. Hdom is also plotted using the site index model. Note the different scales on the x and y axes
Model application
The development of a typical initial stand after shelterwood regeneration at four representative sites along the climatic and local site amplitude of beech stands in Switzerland under a given management schedule (the same in all cases) and four alternative climate scenarios is shown in Fig. 8. Results show how the impact of climate change may vary considerably along the range of current site conditions. While small changes in production are expected at fertile sites with moderate expected changes in climate (Fig. 8A) or at poor sites already facing extreme drought conditions under current climate (Fig. 8D), more relevant changes are expected at sites with limitations by the current temperature sum (Fig. 8B) and at sites currently facing moderate drought (Fig. 8C). It is also of interest to note the clear gradient between the maximum predicted volume at the fertile site (560 m3 ha−1; Fig. 8A) and the maximum predicted volume at the site facing extreme drought (208 m3 ha−1; Fig. 8D).

Simulated development of four representative stands (along the climatic amplitude of beech stands in Switzerland) under four climate scenarios (current climate and three regional circulation model realizations of the IPCC AR4 A1b emission scenarios: SMHI, MPI, HCCPR; Table 3) and a shelterwood management schedule based on two intermediate thinnings (same basal area % removed from all diameter classes) and a 20-year regeneration period (two regeneration cuts): A fertile site under current climate that remains very similar under climate change; B poor site (due to low sum of degree-days) that becomes more productive under climate change; C mean site facing drought in which drought increase decreases production; D stand facing extreme drought (WHC = 79 mm) in which an increase in drought decreases production moderately. The left-hand charts show the evolution of drought and degree-day indices (DI and GDDI, respectively) under current and HCCPR climate scenarios. A typical initial stand at 20 years of age (already tended; 2367 trees ha−1) is used in all cases
Discussion
The basis for the empirical integration of explicit environmental effects in the model was the combination of the available data on stand dynamics and climate at the same temporal and spatial resolution. Besides, the permanent plots used for modelling and validation represented well the range of climatic and site conditions of beech stands in Switzerland, which included some sites and periods under extreme climatic conditions, and the resulting effects from the mixed-effects models facilitated the correct interpretation of spatial and temporal patterns in the data. More observations in very dry plots and hydromorphic soils would have been useful, but we believe the used sample was sufficient for representing the main effects.
The data used for model development, measured at the individual-tree level, provided an adequate representation of management interventions and effects in terms of thinning type and intensity. In simulations, the used individual-tree resolution allows flexible representation of treatments, while effects of those are depicted by the used competition variables (BAL_rem; G_rem; and BAL_thin in Eqs. 3, 4, 5 and 6). The identified thinning effect (BAL_thin in Eqs. 3 and 4) was based on the delay for the coming 5-year period shown by beech trees in using released growing space from above, which was also identified in previous studies (Nord-Larsen and Johansen 2007). Beech trees usually react quickly to moderate crown release, e.g., by growth of new branches; however, it seems reasonable that the remaining trees experience some stress when the release is substantial (Pretzsch 2009). The model also has the desirable property of being sensitive to changes in stem height/diameter ratios, which is useful, e.g., for the calculation of timber assortments.
The individual-tree model set allows accurate and biologically consistent simulation of measured stand development for the sites and management in the region and predicts an expected asymptote in stand basal area (Fig. 6A, E), according to available data and previous studies (Álvarez-González et al. 2010; Schütz and Zingg 2010). These results also showed how the used environmental variables for site evaluation provide suitable predictions of local site productivity (Bontemps and Bouriaud 2014). Statistics were in line with previous growth and yield studies using the same type of data, i.e. permanent plot data that contains rather large random measurement errors which are not predicted by the models (c.f. Palahí et al. 2003; Trasobares et al. 2004). For the diameter growth validation using SNFI data, it should also be remembered that dbh measurements in the SNFI were rounded to the cm instead of mm in the modelling data. When evaluating the obtained validation results (see “Results” section), it is important to note that even though the amount of observations used for validating the model was considerably smaller than for fitting the model, the observations based on the systematic NFI sampling design represented a greater number of plots (locations) and a broader range of environmental conditions than the sample used for fitting the model (Fig. 1). Trends of mean residuals from the diameter growth model versus stand basal area, basal area of larger trees and predicted growth (Fig. 5) can be explained by the different sampling methods used for the modelling and validation data, i.e. plot size in the SNFI is on average seven times smaller than in the long-term WSL plots and the sampling probability of trees of dbh ≥ 36 cm is 2.5 times greater. This affects the representation of spatial variability in the stand and is likely to overestimate the number of trees ha−1 represented by each SNFI tree. If the model is applied using data measured in plots smaller than 0.25 ha, the calibration of model predictions should be considered (e.g. Salas-González et al. 2001).
The data used for survival modelling presented some limitations: (1) the thinned trees, which are often suppressed and in the stage of dying, were not used as observations and (2) mortality related to natural disturbances was only approximately removed, because plot measurements showing clear natural disturbance effects were not used, but weaker effects may have remained in the sample. This created some bias in the survival model without past growth (Eq. 5). However, the use of past growth (Eq. 6) in addition to improving accuracy allowed significant independence from data limitations; the model using past growth is not sensitive to sampling bias created in Eq. 5 caused by the omission of trees killed by natural hazards and thinned trees. This allowed a close representation of density-dependent mortality (Fig. 4). In management planning applications, density-independent mortality caused by disturbances can be integrated using risk models (Schütz et al. 2006; Hanewinkel et al. 2004).
Applicability in practical management planning was a central consideration when developing the growth and yield model. All required variables for simulating stand development (diameter distribution, height from some representative trees, estimates from digital climatic maps) are usually available from forest inventories and climatic maps, though WHC might not always be explicitly available. For fitting the model, we used WHC estimates of the soil, which were obtained using soil pedotransfer functions. Such estimates contain some uncertainty inherent to the used function and estimates from soil pits, but permit the integration of local site potential in simulations, which is one of the advantages of this model. However, if this information is not available, simpler methods can be used as well such as integrating available texture diagrams and/or soil maps. The use of log-transformed 5-year growth in both the diameter and height growth models resulted in multiplicative models, which is a desirable feature in terms of biological consistency when thinking of the representation of environmental changes by drought and degree-day indices. The model provided coherent simulation of stand development under climate change along a wide range of the current site amplitude of beech (Fig. 8) and highlighted that the impact of climate change may vary considerably among sites. The model also showed sensitivity to the gradient between different site conditions (Fig. 8A, D). The increase in production due to the extension of the growth period in sites with temperature limitations under current climate (Fig. 8b) was consistent and in line with recent studies on tree phenology for the species (Vitasse et al. 2011). In dry sites (Fig. 8C, D), simulations were also coherent, but the drought index reached values lower than 0.7, i.e. growth–drought index relationships in the growth models (Eqs. 3 and 4) were used beyond the available limit in the modelling data. Some authors argue that under changing environmental conditions some of the fixed relationships in empirical models may change (Kramer et al. 2008; Fontes et al. 2010). We believe, however, that the relationships established in this model, which are based on long-term climatic series that included various extreme periods, should provide reasonably valid extrapolations, at least not far beyond the limits of the used data. For example, minimum values for drought and degree-day indices were lower in the SNFI sample than in the modelling sample (see Fig. 3) but followed the modelled relationship.
The intervals between measurements in the data used for fitting the model were quite large compared to likely response periods of trees and stands to climate effects. If the model is used for simulating 5-year periods, which are comparable to measurement periods, climate effects on growth should be well represented. Sensitivity to simulated effects of climate on growth in shorter periods (1–3 years) should be evaluated using data that consider annual effects of climate on tree growth. Another aspect to be considered is whether monthly climatic indicators (potential evapotranspiration and degree-days) sufficiently reflect daily extreme climate change impacts (e.g. a period of few days of extreme temperature in early summer). Even though the used modelling data included such extreme periods, the resulting degree-day sum of months including those extreme days may not be too unusual because weather conditions during the rest of days in those months may have been milder. Hence, future studies could concentrate on additional validations using data from beech stands under more severe drought conditions and comparisons with existing process-based models (Pietsch et al. 2005; Seidl et al. 2005; Rasche et al. 2011), especially beyond the limits of the used data.
The model set developed in this study allows tree-level distance-independent simulation of stand development for a broad range of management and climate scenarios in Switzerland, which could also be tested in neighbouring regions. The growth and yield model allows the direct integration of environmental effects in dynamic growth and survival functions without need of fitting intermediate models and functions for the prediction of an implicit site parameter. The model provides accurate predictions, is sensitive to management effects, uses normally available input data, requires low computational effort and therefore is suitable for advanced management planning.
References
Álvarez-González JG, Zingg A, Von Gadow K (2010) Estimating growth in beech forests: a study based on long term experiments in Switzerland. Ann For Sci 67:307. doi:10.1051/forest/2009113
Baldwin VC Jr, Burkhart HA, Westfall JA, Peterson KD (2001) Linking growth and yield and process models to estimate impact of environmental changes on growth of loblolly pine. For Sci 47(1):77–82
Belsley DA, Kuh E, Welsch RE (1980) Regression diagnostics: identifying influential data and sources of collinearity. Wiley, Hoboken
Bigler C, Bugmann H (2004) Predicting the time of tree death using dendrochronological data. Ecol Appl 14:902–914
Bontemps JD, Bouriaud O (2014) Predictive approaches to forest site productivity: recent trends, challenges and future perspectives. Forestry 87:109–128. doi:10.1093/forestry/cpt034
Collins M, Booth BBB, Harris GR, Murphy JM, Sexton DMH, Webb MJ (2006) Towards quantifying uncertainty in transient climate change. Clim Dyn 27:127–147
Didion M, Kupferschmid A, Zingg A, Fahse L, Bugmann H (2009) Gaining local accuracy while not losing generality—Extending the range of gap model applications. Can J For Res 39:1092–1107
Dobbertin M (2005) Tree growth as indicator of tree vitality and of tree reaction to environmental stress: a review. Eur J For Res 124:319–333
Flewelling JW, Pienaar LV (1981) Multiplicative regression with lognormal errors. For Sci 27:281–289
Fontes L, Bontemps JD, Bugmann H, Van Oijen M, Gracia C, Kramer K, Lindner M, Rötzer T, Skovsgaard JP (2010) Models for supporting forest management in a changing environment. For Syst 19:8–29
Gee GW, Bauder JW (1986) Particle-size analysis. In: Klute A (ed) Methods of soil analysis. Part 1 physical and mineralogical methods, 2nd edn. American Society of Agronomy, Madison, pp 383–411
Goldstein H (1996) Multilevel statistical models. Arnold, London
González JR, Palahí M, Pukkala T, Trasobares A (2005) Integrating fire risk considerations in forest management planning—a landscape level perspective. Lands Ecol 20(8):957–970
González-García M, Hevia A, Majada J, Calvo de Anta R, Barrio-Anta M (2015) Dynamic growth and yield model including environmental factors for Eucalyptus nitens (Deane & Maiden) Maiden short rotation woody crops in Northwest Spain. New Forest 46:387–407. doi:10.1007/s11056-015-9467-7
Gracia CA, Tello E, Sabaté S, Bellot J (1999) GOTILWA: an integrated model of water dynamics and forest growth. In: Rodà F et al (eds) Ecology of Mediterranean evergreen oak forests. Springer, Berlin, pp 163–179
Hanewinkel M, Zhou W, Schill C (2004) A neural network approach to identify forest stands susceptible to wind damage. For Ecol Manage 196:227–243
Härkönen S, Pulkkinen M, Duursma R, Mäkelä A (2010) Estimating annual GPP, NPP and stem growth in Finland. For Ecol Manage 259:524–533. doi:10.1016/j.foreco.2009.11.009
Hollweg HD, Böhm U, Fast I, Hennemuth B, Keuler K, Keup-Thiel E, Lautenschlager M, Legutke S, Radtke K, Rockel B, Schubert M, Will A, Woldt M, Wunram C (2008) Ensemble Simulations over Europe with the Regional Climate Model CLM forced with IPCC AR4 Global Scenarios. Max Planck Institute for Meteorology, Hamburg
Jacobsen JB, Thorsen BJ (2003) A Danish example of optimal thinning strategies in mixed-species forest under changing growth conditions caused by climate change. For Ecol Man 180:375–388
Kellomäki S, Peltola H, Strandman H, Väisänen H (2015) Changes in growth of Scots pine, Norway spruce and birch induced by climate change in boreal conditions as related to prevailing growing conditions. Manuscript in preparation
Kjellström E, Nikulin G, Hansson U, Strandberg G, Ullerstig A (2011) 21st century changes in the European climate: uncertainties derived from an ensemble of regional climate model simulations. Tellus A 63:24–40
Kramer K, Buiteveld J, Forstreuter M, Geburek T, Leonardi S, Menozzi P, Povillon F, Schelhaas MJ, Teissier du Cross E, Vendramin GG, Van der Werf DC (2008) Bridging the gap between ecophysiological and genetic knowledge to assess the adaptive potential of European beech. Ecol Model 216:333–353. doi:10.1016/j.ecolmodel.2008.05.004
Matala J, Ojansuu R, Peltola H, Sievanen R, Kellömaki S (2005) Introducing effects of temperature and CO2 elevation on tree growth into a statistical growth and yield model. Ecol Model 181:173–190
Monserud RA (1976) Simulation of forest tree mortality. For Sci 22:438–444
Nord-Larsen T, Johannsen VK (2007) A state space approach to stand growth modelling of European beech. Ann For Sci 64(4):365–374
Palahí M, Pukkala T, Miina J, Montero G (2003) Individual-tree growth and mortality models for Scots pine (Pinus sylvestris L.) in north-east Spain. Ann For Sci 60:1–10
Pietsch SA, Hasenauer H, Thornton PE (2005) BGC-model parameters for tree species growing in central European forests. For Ecol Man 211(3):264–295
Pretzsch H (2009) Forest dynamics, growth and yield. Springer, Berlin
Pukkala T (2002) Multi-objective forest planning. Kluwer Academic Publishers, Dordrecht
Pukkala T, Kellomäki S (2012) Anticipatory vs adaptive optimization of stand management when tree growth and timber prices are stochastic. Forestry 85(4):463–472. doi:10.1093/forestry/cps043
Rasche L, Fahse L, Zingg A, Bugmann H (2011) Getting a virtual forester fit for the challenge of climatic change. J App Ecol 48(5):1174–1186
Salas-González R, Houllier F, Lemoine B, Pignard G (2001) Forecasting wood resources on the basis of national forest inventory data: application to Pinus pinaster in southwestern France. Ann For Sci 58:785–802
SAS Institute Inc (2011) SAS/STAT® 9.3 User’s Guide. Cary, NC: SAS Institute Inc
Schröder J, Von Gadow K (1999) Testing a new competition index for Maritime pine in northwestern Spain. Can J For Res 29:280–283
Schütz JP, Zingg A (2010) Improving estimations of maximal stand density by combining Reineke’s size-density rule and the yield level, using the example of spruce (Picea abies (L.) Karst.) and European Beech (Fagus sylvatica L.). Ann For Sci 67:507. doi:10.1051/forest/2010009
Schütz JP, Götz M, Schmid W, Mandallaz D (2006) Vulnerability of spruce (Picea abies) and beech (Fagus sylvatica) forest stands to storms and consequences for silviculture. Eur J For Res 125:291–302. doi:10.1007/s10342-006-0111-0
Seidl R, Lexer MJ, Jager D, Honninger K (2005) Evaluating the accuracy and generality of a hybrid patch model. Tree Phys 25(7):939–951
Seynave I, Gegout JC, Hervé JC, Dhôte JF, Drapier J, Bruno E, Dume G (2005) Picea abies site index prediction by environmental factors and understorey vegetation, a two-scale approach based on survey databases. Can J For Res 35(7):1669–1678
Seynave I, Gégout JC, Hervé JC, Dhôte JF (2008) Is the spatial distribution of European beech (Fagus sylvatica L.) limited by its potential height growth? J Biogeogr 35:1851–1862
Sharma M, Subedi N, Ter-Mikaelian M, Parton J (2015) Modeling climatic effects on stand height/site index of plantation-grown jack pine and black spruce trees. For Sci 61(1):25–34. doi:10.5849/forsci.13-190
SPSS Inc (2010) SPSS Base system syntax reference Guide Release 19.0
Snowdon P (1991) A ratio estimator for bias correction in logarithmic regressions. Can J For Res 21:720–724
Teepe R, Dilling H, Beese F (2003) Estimating water retention curves of forest soils from soil texture and bulk density. J Plant Nut Soil Sci 166:111–119
Thornthwaite CW, Mather JR (1957) Instructions and tables for computing potential evapotranspiration and the water balance. Publ Climatol 10:183–311
Thornton PE, Running SW, White MA (1997) Generating surfaces of daily meteorological variables over large regions of complex terrain. J Hydrol 190:214–251. doi:10.1016/S0022-1694(96)03128-9
Trasobares A, Tomé M, Miina J (2004) Growth and yield model for Pinus halepensis Mill. in Catalonia, north-east Spain. For Ecol Man 203:49–62
Vanclay JK (1994) Modelling forest growth and yield: applications to mixed tropical forests. CABI Publishing, Wallingford
Vitasse Y, François Ch, Delpierre N, Dufrêne E, Kremer A, Chuine I, Delzon S (2011) Assessing the effects of climate change on the phenology of European temperate trees. Agric For Meteorol 151:969–980. doi:10.1016/j.agrformet.2011.03.003
Walthert L, Graf U, Kammer A, Luster J, Pezzotta D, Zimmermann S, Hagedorn F (2010) Determination of organic and inorganic carbon, d13C, and nitrogen in soils containing carbonates after acid fumigation with HCl. J Plant Nutr Soil Sci 173:207–216
Waring RH (1983) Estimating forest growth and efficiency in relation to canopy leaf area. Adv Ecol Res 13:327–354
Webster R, Rigling A, Walthert L (1996) An analysis of crown condition of Picea, Fagus and Abies in relation to environment in Switzerland. For 69:347–355
WSL (2010) Swiss National Forest Inventory NFI. Data of the surveys 1983–1985, 1993–1995 and 2004–2006. 280510UU. Swiss Federal Research Institute WSL, Birmensdorf
Wykoff RW (1990) A basal area increment model for individual conifers in the northern Rocky Mountains. For Sci 36:1077–1104
Yousefpour R, Jacobsen JB, Meilby H, Thorsen BJ (2014) Knowledge update in adaptive management of forest resources under climate change: a Bayesian simulation approach. Ann For Sci 71:301–312
Zuber T (2007) Untersuchungen zum Wasserhaushalt eines Fichtenwaldstandorts unter Berücksichtigung der Humusauflage. Dissertation, University of Bayreuth
Acknowledgments
We thank Matthias Dobbertin (WSL) for the provision of Level I (Sanasilva) and Level II forest inventory and soil data. It was nice working with him. We also thank U.-B. Brändli (WSL) for the provision of the SNFI data and Dirk Schmatz and Nick Zimmermann (WSL) for the provision of downscaled climate scenario data. Harald Bugmann is acknowledged for participating actively in the development of the presented approach and providing important support. We thank Jerry Vanclay, Jari Miina, Jette B. Jacobsen, Annikki Mäkelä and the two reviewers for their comments. This research was funded by the MOTIVE project within the European commission’s 7th framework program (Grant agreement No. 226544).
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Aaron R Weiskittel.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Trasobares, A., Zingg, A., Walthert, L. et al. A climate-sensitive empirical growth and yield model for forest management planning of even-aged beech stands. Eur J Forest Res 135, 263–282 (2016). https://doi.org/10.1007/s10342-015-0934-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-015-0934-7