
Abstract
The growth patterns of annually resolved tree rings are good indicators of local environmental changes, making dendrochronology a valuable tool in air pollution research. In the present study, tree-ring analysis was used to assess the effects of 16 years (1991–2007) of chronic nitrogen (N) deposition, and 10 years (1991–2001) of reduced nitrogen input, on the radial growth of Norway spruce (Picea abies (L.) Karst.) and Scots pine (Pinus sylvestris L.) growing in the experimental area of Lake Gårdsjön, southwest Sweden. In addition to the ambient input of c. 15 kg N ha−1 year−1, dissolved NH4NO3 was experimentally added to a 0.52-ha watershed at a rate of c. 40 kg ha−1 year−1. Atmospheric N depositions were reduced by means of a below-canopy plastic roof, which covered a 0.63-ha catchment adjacent to the fertilized site. The paired design of the experiment allowed tree growth in the N-treated sites to be compared with the growth at a reference plot receiving ambient N deposition. Nitrogen fertilization had a negative impact on pine growth, while no changes were observed in spruce. Similarly, the reduction in N and other acidifying compounds resulted in a tendency towards improved radial growth of pine, but it did not significantly affect the spruce growth. These results suggest that spruce is less susceptible to changes in the acidification and N status of the forest ecosystem than pine, at least in the Gårdsjön area.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Nitrogen (N) is one of the most abundant nutrients in the phytomass of terrestrial vegetation and is an important component in metabolic processes of a plant. Thus, the vitality of a tree is highly dependent on a balanced supply of N (Arnold and van Diest 1991). Although most ecosystems of Northern Hemisphere contain large stores of N bound in the soil organic matter, climate and soil conditions typical of these latitudes contribute to a rather tight N cycle in nearly all systems, characterized by an efficient internal cycling and a small loss in runoff. Tree growth in temperate forests has therefore been considered to be highly regulated by the availability of N (Mitchell and Chandler 1939). Increased pollution-derived atmospheric N loadings into an N-limited system may enhance the uptake of N by vegetation, leading to canopy expansion, increased primary production and an accelerated internal N cycle. Accelerated base cation uptake, which follows enhanced growth rates, may lead to nutritional imbalances and deficiencies of macronutrients such as phosphorus (P) and calcium (Ca), which will inhibit the vegetation growth (e.g. Burstrom 1968; Chapin 1980). When the input of N exceeds the total nutritional demand of plants and microbes within the ecosystem, N saturation will occur (Aber et al. 1989). The first sign of saturation is increased nitrate (NO3 −) leaching from below the rooting zone, accompanied by a net proton production and, subsequently, acidification. Leaching of NO3 − may be followed by leaching of base cations, which, in the long term, may reduce the site fertility and result in ecosystem malfunction such as disturbances in the mycorrhiza development and function, reduced root growth and increased susceptibility to damage by animals, fungi, bacteria and viruses (Nihlgard 1985). Acidification may further cause mobilization of aluminium, particularly the Al3+ ion, leading to root damage and aluminium toxicity of many plants.
Even though the atmospheric deposition of N compounds has decreased by approximately 20 % during the period 1989–1998 over southern Scandinavia (Wright et al. 2001), there are still concerns for nitrogen saturation in forest and aquatic ecosystems in Sweden. The ecosystem-scale experiments in the area of Lake Gårdsjön, on the west coast of Sweden, were designed to obtain a better understanding of the relationship between the conditions of coniferous forest ecosystems, pollution and other stress factors (Wright and van Breemen 1995). Field experiments, including nitrogen fertilization, were conducted in several sub-catchments within the Lake Gårdsjön area as a part of the project. To overview the influence of the experimental treatments, responses of runoff (Moldan and Wright 1998a, b; Moldan et al. 2006), ground vegetation, soils and soil solution (Stuanes et al. 1995), N2O emission (Klemedtsson et al. 1997) and the internal nitrogen cycle (Kjønaas et al. 1998), fine root (Clemensson-Lindell and Persson 1995) and mycorrhiza (Brandrud 1995) have been assessed.
The present work examines the radial growth of Norway spruce (Picea abies (L.) Karst.) and Scots pine (Pinus sylvestris L.) in the experimentally treated watersheds at the Gårdsjön research site. The growth of a tree is a function of a range of environmental parameters, including climate, stand dynamics and exposure to pollutants (Fritts 2001). Thus, annual radial growth increments can provide valuable historical records of past local environmental conditions and be used to link alterations of various environmental parameters to tree health (Cook et al. 1987). Dendroecological methods have frequently been used to describe growth reductions associated with pollutant exposure on local scale (Ashby and Fritts 1972; Nash et al. 1975; Nöjd and Reams 1996; Hirano and Morimoto 1999; Long and Davis 1999; Boone et al. 2004), as well as regional scale (McLaughlin and Percy 1999; Dittmar et al. 2003). Here, dendroecological techniques were used to examine the effects on conifer growth of (1) 16 years of chronic nitrogen fertilization and (2) 11 years of exclusion of N deposition. The experimental effect was estimated by comparing the tree growth at the impact sites with that of an adjacent, non-treated reference site.
Materials and methods
Study site and treatment descriptions
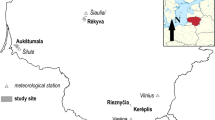
The radial tree growth of Norway spruce and Scots pine was studied in the area of Lake Gårdsjön (58°04′N, 12°03′E, 135–145 m a.s.l.), located approximately 10 km east of the Swedish west coast and 50 km north of Gothenburg City (Fig. 1). Proximity to the sea yields a maritime climate in this region, characterized by mild winters and cool summers. Mean annual temperature and total annual precipitation are 6.4 °C and 1,100 mm, respectively. The area receives moderate depositions of sulphate, nitrate and ammonium; mean throughfall inputs (1989–1996) were estimated to be 25 kg SO4–S ha−1 year−1, 7.3 kg NO3–N ha−1 year−1 and 4.8 kg NH4–N ha−1 year−1 (Moldan et al. 2006). The bedrock in the region is dominated by granites and granodiorites. Soils are dominantly thin (<0.5 m) sandy and silty podzols with inclusions of bedrock outcrops and thicker organic soils in the valley bottoms. The C/N ratio ranges between 34 and 38 (whole soil, mol/mol), and the soil N content is approximately 8,600 kg ha−1 (Moldan et al. 2006). The forest in the Gårdsjön area consists of mixed mature conifer trees, where Norway spruce dominates the stands and inclusions of Scots pine are found in the drier areas.

Map showing the location of the experimental area of Lake Gårdsjön, on the west coast of Sweden. Black triangles in the southern part of the lake mark the position of the manipulated (G1 and G2) and the reference (F1) plots
For this study, three sub-catchments, G1, G2 and F1, all situated at the southern end of the lake in the Gårdsjön basin, were chosen (Fig. 1). All the three catchments had relatively similar valley-shaped topographies, with flat or gently sloping bottoms and steep sides. As a part of the European NITREX project (NITrogen saturation EXperiments; Wright and van Breemen 1995), the G2 NITREX catchment (0.52 ha) was experimentally manipulated by the addition of ammonium nitrate (NH4NO3) from April 1991 onwards (Moldan et al. 1995). NH4NO3 was dissolved in deionized water and applied to the whole, originally N-poor, catchment area by means of a sprinkling system. The nitrogen was distributed in weekly and biweekly doses, in proportion to the volume of ambient throughfall. The volumes of the additional water corresponded to 5 % of the natural precipitation, and the experimental addition of N was approximately 40 kg ha−1 year−1 (Moldan et al. 2006). The second catchment, G1 ROOF (0.63 ha), was entirely covered between April 1991 and 2001 by a transparent roof, constructed at a height of 2–4 m beneath the forest canopy (Moldan et al. 2004). Here, the acidic throughfall was intercepted by the roof and replaced by water pumped from Lake Gårdsjön, deionized, spiked with sea salt and sprinkled by means of an irrigation system underneath the roof construction to simulate rain, typical of the region but under less-polluted conditions. The artificial throughfall contained small amount of added KNO3 as the only N compound and had a lower concentration of sulphur (18–29 μM) compared with natural throughfall (125 μM) in the region (Moldan et al. 2004). The frequency and duration of each sprinkling event were dependent on the natural precipitation in the area. The litter intercepted by the roof was collected and placed on the soil underneath the construction (Bishop and Hultberg 1995). The third catchment chosen for this study, F1 CONTROL (3.7 ha), was not subjected to any experimental manipulation, but received ambient levels of atmospheric deposition and was consequently serving as a reference site for this study.
Chronology construction
In each of the three catchment areas, trees of Norway spruce and Scots pine, representing a fairly uniform age class of 93–121 years (Table 1), were randomly selected for tree-growth measurements. Using an increment borer, two-core samples, 4 mm in diameter, were sampled at a height of about 1.3 m from 12 to 16 trees from each species and at each site. To avoid reaction wood, the samples were taken in opposite directions of the trunk, parallel to the topographic contour. All the tree-ring data were collected in the early summer of 2007.
The tree cores were subsequently dried and mounted, and the cores were surfaced with a razor blade to enhance the appearance of ring boundaries and the cell structure. Annual tree-ring widths were measured with a precision of ±0.001 mm using a stereomicroscope coupled to a Lintab measurement table connected to the Time Series Analysis Program (TSAP) software (Rinntech, Heidelberg, Germany). To exclude possible measurement biases and to correctly ascertain the year in which each tree ring was formed, the samples were cross-dated through the matching of ring-width patterns of cores from each tree and among different trees of the same species. This procedure was performed visually and verified statistically using the COFECHA software (Holmes 1999). Since it was desirable to remove low-frequency age-related growth changes from the data, where the growth rate commonly declines monotonically with tree maturation (Fritts 2001), all individual tree-ring series were standardized using the ARSTAN software (Cook and Holmes 1986). The standardization procedure was performed by fitting a negative exponential curve, or a regression line, to the ring-width series and then dividing each observed ring-width value by the corresponding curve value. Standard-version chronologies were produced for each site and species as a biweight robust mean of the remaining dimensionless tree-ring indices and retained for the tree-growth assessment. For climate–growth response, residual chronologies were constructed after removing autocorrelation from individual series.
Tree-growth response
It is well established that climate is one of the most important factors influencing tree growth (Fritts 2001). Due to the close proximity between studied catchments, the same local climatic forcing most likely influenced the tree growth at each site, leading to the assumption that the tree-growth pattern among sites should be similar would the site treatments not have been introduced. This hypothesis was tested through an evaluation of the relationship between climate and tree growth. Long-term environmental disturbances, such as air pollution, likely manifest as a low-frequency variation in the ring-width data (e.g. McClenahen and Dochinger 1985; Sutherland and Martin 1990). In order to omit any potential low-frequency variations in the tree growth caused by the different manipulations and to enhance the climate-related year-to-year tree-ring signal, prewhitened residual chronologies were used to assess the climate–growth relationship. Response function analysis was performed between tree-ring indices and monthly and seasonal precipitation and temperature data from Säve meteorological station located c. 30 km from Gårdsjön (57°48′N, 11°54′E) and covering the 1911–2005 period. Analysis was carried out in DENDROCLIM2002 software (Biondi and Waikul 2004), which uses 1,000 bootstrapped samples to compute response coefficients and to test their significance at a 0.05 level.
The growth of pine and spruce at the control site represents natural (i.e. unaffected) conditions. If manipulation at sites G1 and G2 has affected the tree-ring chronologies, they would differ from control site in lower frequencies. Standard-version chronologies, retaining the time-persistence in the tree-ring data, were thus used to compare radial tree growth between sites. The control chronologies were compared with the G1 and G2 site chronologies by means of product–moment correlation coefficient and paired t test to determine whether the mean difference between tree-ring series was significantly greater than 0. The statistics were calculated for two 16-year periods (1959–1974 and 1975–1990) before each treatment initiation and one 16-year period (1991–2006) after the onset of the manipulation. Furthermore, to highlight the low-frequency variability and to compare the chronologies graphically with each other, each chronology was smoothed using locally weighted regression (loess filter with a 10-year window; Cleveland and Devlin 1988).
Randomized intervention analysis
Randomized intervention analysis (RIA) (Carpenter et al. 1989) was used to investigate whether a non-random change had occurred in tree growth as a response to the artificial N addition and acidic throughfall exclusion in the G2 and G1 sites, respectively. This statistical test has especially been designed for ecosystem experiments, where it is desirable to detect possible impacts of an intervention. The RIA null hypothesis assumes that all observed alterations in a manipulated ecosystem are random, that is, any relationships apparent in the data are simply due to chance in the sampling process. Unlike many other statistical tests, RIA is not affected by moderate serial autocorrelation, heterogeneity of variance or non-normality. To estimate the test statistic, paired tree-growth index time series from both the control and the manipulated sites were plotted. The time sequence used in the analysis covered the two 16-year periods before (1959–1990) and the 16-year period after (1991–2006) the treatment initiations at the G1 and G2 sites. Differences, [(I t (exp.) − (I t (ref.)] = D t , for the time-paired tree-growth observations, I t , were calculated, where exp. and ref. refers to the manipulated and the control sites, respectively. Mean intersystem differences before and during each intervention, \( \bar{D}\,({\text{pre}}) \) and \( \bar{D}\,({\text{post}}) \), respectively, were then computed. The absolute value of the difference between \( \bar{D}\,({\text{pre}}) \) and \( \bar{D}\,({\text{post}}) \) is the test statistic. Distribution of the test statistic was approximated by a Monte Carlo simulation, where the time series of differences, D t , were randomly shuffled 1,000 times. For each shuffling, \( \left| {\bar{D}\,({\text{pre}}) - \bar{D}\,({\text{post)}}} \right| \) was examined. The proportion of the \( \left| {\bar{D}\,({\text{pre}}) - \bar{D}\,({\text{post)}}} \right| \) that exceeded the observed value was the P value (Carpenter et al. 1989).
Results
Radial growth
The spruce and pine chronologies, obtained from 12 to 16 trees from each site, ranged from 102 to 138 years (Table 1). Average sample age was just above 100 years, with the oldest trees found at the G2 site. Mean, unstandardized ring-width chronologies and their standard deviations for each site and species are given in Fig. 2 (to cover a common period, all chronologies were truncated before 1910). The tree-growth curves of individual series showed different trends (figure not shown), some exhibiting an exponential-like decay and some showing the effects of suppression–release common for a mesic, closed-canopy forest environment. Ring widths were averaged for two periods, corresponding to a pre-treatment (1910–1990) and a treatment time interval (1991–2006; Table 1). Almost all sites showed the narrowest ring widths during the latter period, likely caused by the tree-ageing effect (Fritts 2001). A general decline in the standard deviations of the averaged ring widths during the treatment period provided an indication of a decrease in the sensitivity of each individual tree, suggesting a more homogenous response of the trees with increasing maturation (Table 1; Fig. 2). The mean sensitivity ranged from 0.212 to 0.263, with slightly higher values for pine.

Mean radial growth of spruce and pine from the G1 and G2 sites for the period 1910–2006. Standard deviations are illustrated by vertical bars
Comparison with the reference site
Similar tree-growth responses to climate were found at all sites; the growth of spruce was benefitted from enhanced June precipitation, while pine was positively affected by mild late-winter temperatures (Table 2). By establishing that the tree-growth variations are controlled by the same climate forcing, it can be concluded that the growth pattern in the treated sites would have been similar to that in the control site would the treatments not have been initiated. By reintroducing the persistence in the tree-ring data, the control and manipulated sites can be compared and intervention-related discrepancies in tree growth detected.
When comparing standardized chronologies developed for the manipulated sites with the reference chronologies, a rather synchronous growth pattern emerges (Fig. 3). The growth of spruce exhibited peaks in the 1960s and 1980s and a depression in the 1970s, while increased pine growth was conspicuous in the 1960s and in the early 1990s. Long-term growth trends were generally consistent among the experimental and control plots during the last decades (Fig. 4). The time period starting from 1990s onwards was characterized by a continuously decreasing radial pine growth at all sites. A similar negative trend was apparent for spruce. However, in contrast to pine, the starting point for the spruce growth reduction was in the middle of the 1980s, while a gradual recovery was somewhat apparent from the middle of the 1990s.

Comparison of standardized tree-ring indices (TRI, standard version) of spruce and pine from the G1 and G2 sites (black lines) with growth indices of spruce and pine from the untreated F1 site (grey lines)

Standard-version spruce and pine chronologies from the G1 and G2 sites. Data are been smoothed with a 10-year low-pass (first-order) loess filter (Cleveland and Devlin 1988)
Changes in the tree growth at the treated sites were evaluated by statistically comparing the growth series with those of the control site (Table 3). In the pre-manipulation periods, as well as in post-manipulation interval, the correlation between the chronologies was high and statistically significant at all sites (P < 0.01). The paired t test revealed that the mean pine growth at the G2 site, during the treatment period, was significantly (P < 0.01) lower than that at the control chronology, while the growth of spruce was slightly stronger (P < 0.05). At the G1 plot, only the mean growth of pine differed significantly (P < 0.01), in this case exceeding the reference site.
To further establish whether a non-random tree-growth change had occurred following the site treatments, RIA was performed on paired data sets from the treatment and the reference sites (Fig. 5). The probability of rejection of the RIA null hypothesis is shown in Table 4. Pine growth at the G1 and G2 sites had a rejection limit exceeding 99 % (P < 0.01). Hence, the pine growth could be rejected safely as exhibiting a non-random behaviour following the initiation of each experiment. Autocorrelation in the time series of intersystem differences might cause RIA to underestimate the true P value. It has been suggested that the P value from RIA should be less than 0.01 to reject the null hypothesis if a time series is autocorrelated (Carpenter et al. 1989). The spruce growth in the G1 and G2 sites had a rejection limit below 99 %. Hence, it could not safely be assumed that spruce exhibited a non-random growth change in either of the sites.

Calculations for randomized intervention analysis. The thick lines represent the difference, [(I t (exp.) − (I t (ref.)], between standardized tree-ring indices (I t ) from each manipulated site (exp.) and the untreated F1 site (ref.). Mean differences for the pre-treatment period (1959–1990) and the treatment period (1991–2006), \( \bar{I}_{t} \,({\text{pre}} .) \) and \( \bar{I}_{t} \,({\text{post}} .) \), are represented by dashed lines
Discussion
Since the G2 plot was initially N–limited, enhanced tree growth was expected to accompany the increased N input. However, while no prominent discrepancy between the growth of spruce at the fertilized and the reference sites was observed, the pines at the G2 plot seemed, on the contrary, to show a N-saturation effect by significantly decreasing the wood productivity during fertilization with NH4NO3. Previous N-fertilization studies on the stemwood growth of conifer and deciduous trees have showed various results: increased and decreased growth (Mälkönen et al. 1990; Nilsson and Wiklund 1992; Nohrstedt et al. 1993; Pettersson 1994; Sikström 1997; Magill et al. 2004; Wallace et al. 2007) and small or no effects (Sikström 1997; Persson et al. 1995) have been observed. Reduced growth of Norway spruce in German forests (Wright et al. 1995) has been attributed to ambient input of N, reaching levels of approximately 50 kg N ha−1 year−1. Binkley and Högberg (1997) suggested that this deterioration might be associated with nutritional problems such as magnesium (Mg) deficiency. Reduction in the pine growth at the G2 site is not likely related to Mg shortage. Mg concentration at the G2 plot was optimal at the start of the experiment (Kjønaas and Stuanes 2008), and, due to the proximity to the sea, the area received moderate input of Mg from the sea salt deposition throughout the whole treatment period (Binkley and Högberg 1997).
Previous results from the area suggest that the system of the G2 plot reached an N-saturation point in a relatively short time after the onset of the experiment. Increased NO3 − concentrations were observed in the runoff immediately after the treatment initiation (Hultberg et al. 1994; Moldan et al. 1995; Wright et al. 1995) and proceeded to rise throughout the treatment period from <1 μeq L−1 in April 1991 to c. 70 μeq L−1, accounting for approximately 10 % of the total input, in 2004 (Moldan et al. 2006). Moreover, the N content in litter and foliage increased within a few years of the treatment (Boxman et al. 1998; Kjønaas et al. 1998). These observed responses, typical for an N-saturated system, indicate that the N cycle had been affected in the G2 plot. Even though the N treatment might have caused a fertilizing effect in the G2 plot in the short run, our data suggest that the duration of this effect was not long-lasting enough to enhance the radial wood production. Kjønaas and Stuanes (2008) proposed that the slow tree-growth response at the G2 plot might partly be explained by the fast immobilization of incoming N into the soil pool; only about 9 % of the added N was incorporated into the spruce biomass during the first years of the experiment. The increased NO3 − leaching shortly after the initiation of the treatment has likely caused alterations in the nutrient status of the system so that the tree growth is now more or less inhibited by a nutrient shortage and therefore presumably more susceptible to secondary stress factors such as drought, frost, fungi and parasites (Nihlgard 1985). Tree growth in the Gårdsjön area was already limited at the start of the experiment by the restricted availability of major nutrient elements (N, P, K, Ca). The decrease in the N/nutrient ratios (Mg/N, P/N, K/N) in the spruce needles at the G2 plot throughout the experimental period, especially in the P/N ratio that, at end of the experiment, reached proportions falling well below the critical level (<10–12; Kjønaas and Stuanes 2008), has proved that the nutrient uptake has indeed become even more limited at the G2 site.
Previous findings from the G1 site demonstrate an improved quality of the runoff following the roof construction; the concentrations of S, Al and base cations declined, while the pH slightly rose (Moldan et al. 2004). Fine roots are sensitive to changes in the soil environment. Thus, different root parameters such as tree fine root biomass and fine root Ca/Al molar ratio have been used as indicators of plant nutritional status (e.g. Lamersdorf and Borken 2004; Helmisaari et al. 2007; Vanguelova et al. 2007; Zang et al. 2011). Increased percentage of living fine roots in the organic soil layer within the G1 plot (Clemensson-Lindell and Persson 1995) and better mineral nutrient status of fine roots (increased K, P and Ca levels in relation to N; Persson et al. 1998) suggest that tree-growth conditions had improved in the G1 site following the roof construction. Indeed, pine growth at this site significantly exceeded that of the reference site throughout the treatment period. The G1 site was N poor before and during the experiment. The improved growth at the site might therefore indicate that changes in the acidification rather than in the N status of the system are important for the tree growth in the area.
In contrast to the results obtained for pine and despite observed signs of reduced and increased amounts of spruce fine roots in the G2 and G1 sites (Clemensson-Lindell and Persson 1995), respectively, the growth of spruce showed no distinct tendency towards either improved or reduced growth following the site interventions. Fine roots may be the first biological components to respond to changes in the soil environment (Persson and Ahlström 1999). The increased amount of fine roots in the G1 plot may have improved the tree water uptake, thereby increasing the assimilation rate, especially on dry summer days. However, the energy cost associated with the root growth may, on the other hand, have ceased the growth rate of the aboveground tree structures.
Our results could be interpreted as an indication of a somewhat stronger sensitivity of pine trees to changes in the N status of the forested ecosystem, which, on the other hand, may seem contradictory since it has been showed that Scots pine is well adapted to conditions of poor nutrient availability and that the productivity of pine exceeds that of Norway spruce at sites with low fertility (Ilvessalo 1927). The lack of any response in the case of spruce could also simply be attributed to the site characteristics of each individual tree-growth spot. The sensitivities of individual trees and microsite conditions may partly determine whether fertilization with N will cause tree mortality or increased wood productivity (Wallace et al. 2007). It is important to remember that trees were sampled randomly in the whole watersheds with no regard to the soil moisture status, topography, soil depth, ground vegetation or other factors potentially influencing the sequestration and leaching of N and thereby the tree exposure to N and the nutrient availability. For instance, trees growing in the lower parts of the terrain in the G2 plot were probably more exposed to N, since any added N into the system was eventually drained through this region.
Conclusions
Earlier studies have suggested that elevated N input in previously N-limited temperate forests often results in initially increased tree N uptake and subsequently enhanced productivity. Our results showed, on the contrary, that 16 years of fertilization with 40 kg N ha−1 year−1 did have a significant negative impact on radial pine growth (N-saturation effect), but not on spruce. N saturation of the system shortly after the treatment initiation followed by a reduction in site fertility was a probable cause of the decline in pine growth.
Excluding inorganic N and other acidifying substances significantly enhanced the productivity of pine, but not spruce, which, in consistent with previous findings from the site, suggests that the growth conditions for pine improved following the acidic exclusion. The absence of any significant tendency of either improved or deteriorated growth of spruce following the treatment initiations suggests that the radial growth of spruce is less susceptible to changes in the N status of the forest system, compared with that of pine. However, the lack of any significant responses to the interventions may also be explained by the characteristics of the sites.
References
Aber JD, Nadelhoffer KJ, Steudler P, Melillo JM (1989) Nitrogen saturation in northern forest ecosystems. Bioscience 39:378–386
Arnold G, van Diest A (1991) Nitrogen supply, tree growth and soil acidification. Fertilizer Res 27:29–38
Ashby WC, Fritts HC (1972) Tree growth, air pollution, and climate near LaPorte, Indiana. Bull Am Meteorol Soc 53(3):246–251
Binkley D, Högberg P (1997) Does atmospheric deposition of nitrogen threaten Swedish forests? For Ecol Manag 92:119–152
Biondi F, Waikul K (2004) DENDROCLIM2002: a C++ program for statistical calibration of climate signals in tree-ring chronologies. Comput Geosci 30:303–331
Bishop HK, Hultberg H (1995) Reversing acidification in a forest ecosystem: the Gårdsjön covered catchment. Ambio 24(2):85–91
Boone R, Tardif J, Westwood R (2004) Radial growth of oak and aspen near a coal-fired station, Manitoba, Canada. Tree-Ring Res 60(1):45–58
Boxman AW, Blanck K, Brandrud TE, Emmett BA, Gundersen P, Hogervorst RF, Kjønaas OJ, Persson H, Timmermann V (1998) Cross site comparison of vegetation and soil fauna response to experimentally changed nitrogen inputs in coniferous ecosystems in the EC-NITREX project. For Ecol Manag 101:65–80
Brandrud TE (1995) The effects of experimental nitrogen addition on the ectomycorrhizal fungus flora in an oligotrophic spruce forest at Gårdsjön, Sweden. For Ecol Manag 71:111–122
Burstrom HG (1968) Calcium and plant growth. Biol Rev 43(3):287–316
Carpenter RS, Frost MT, Heisey D, Kratz KT (1989) Randomized intervention analysis and the interpretation of whole-ecosystem experiments. Ecology 70(4):1142–1152
Chapin FS (1980) The mineral nutrition of wild plants. Annu Rev Ecol Syst 11:233–260
Clemensson-Lindell A, Persson H (1995) The effects of nitrogen addition and removal on Norway spruce fine-root vitality and distribution in three catchment areas at Gårdsjön. For Ecol Manag 71:123–131
Cleveland WS, Devlin SJ (1988) Locally weighted regression: an approach to regression analysis by local fitting. J Am Stat Assoc 83:596–610
Cook ER, Holmes RL (1986) Users manual for program ARSTAN. In: Holmes RL, Adams RK, Fritts HC (eds) Tree-ring chronologies of Western North America: California, eastern Oregon and northern Great Basin. Chronology Series 6. The University of Arizona, Tucson, pp 50–65
Cook ER, Johnson AH, Blasing TJ (1987) Forest decline: modelling the effect of climate in tree rings. Tree Physiol 3:27–40
Dawdy DR, Matalas NC (1964) Statistical and probability analysis of hydrologic data, part III: analysis of variance, covariance and time series. In: Te Chow V (ed) Handbook of applied hydrology, a compendium of water-resources technology. McGraw-Hill, New York, pp 8.68–8.90
Dittmar C, Zech W, Elling W (2003) Growth variations of common beech (Fagus sylvatica L.) under different climatic and environmental conditions in Europe—a dendroecological study. For Ecol Manag 173:63–78
Fritts HC (2001) Tree rings and climate. Blackburn Press, Caldwell
Helmisaari H-S, Derome J, Nöjd P, Kukkola M (2007) Fine root biomass in relation to site and stand characteristics in Norway spruce and Scots pine stands. Tree Physiol 27:1493–1504
Hirano T, Morimoto K (1999) Growth reduction of the Japanese black pine corresponding to an air pollution episode. Environ Pollut 106:5–12
Holmes RL (1999) Users manual for program COFECHA. Laboratory of Tree-Ring Research, University of Arizona, Arizona
Hultberg H, Dise NB, Wright RF, Andersson I, Nyström U (1994) Nitrogen saturation induced during winter by experimental NH4NO3 addition to a forested catchment. Environ Pollut 84:145–147
Ilvessalo Y (1927) The forests of Finland. Results of the general survey of the forests of the country carried out during the years 1921–1924. Commun Inst For Fenn 11:1–192
Kjønaas OJ, Stuanes AO (2008) Effects of experimentally altered N input on foliage, litter production and increment in a Norway spruce stand, Gårdsjön, Sweden over a 12-year period. Int J Environ Stud 65:433–465
Kjønaas OJ, Stuanes AO, Huse M (1998) Effects of chronic nitrogen addition on N cycling in a coniferous forest catchment, Gårdsjön, Sweden. For Ecol Manag 101:227–250
Klemedtsson L, Kasimir Klemedtsson Å, Moldan F, Weslien P (1997) Nitrous oxide emission in Swedish forest soils in relation to liming and simulated increased N-deposition. Biol Fertil Soils 25:290–295
Lamersdorf PN, Borken W (2004) Clean rain promotes fine root growth and soil respiration in a Norway spruce forest. Glob Change Biol 10:1351–1362
Long RP, Davis DD (1999) Growth variation of white oak subjected to historical levels of fluctuating air pollution. Environ Pollut 106:193–202
Magill AH, Aber JD, Currie WS, Nadelhoffer KJ, Martin ME, McDowell WH, Melillo JM, Steudler P (2004) Ecosystem response to 15 years of chronic nitrogen additions at the Harvard Forest LTER, Massachusetts, USA. For Ecol Manag 196:7–28
Mälkönen E, Derome J, Kukkola M (1990) Effects of nitrogen inputs on forest ecosystems estimation based on long-term fertilization experiments. In: Kauppi P, Anttila P, Kenttanries K (eds) Acidification in Finland. Springer, Berlin, pp 325–347
McClenahen JR, Dochinger LS (1985) Tree ring response of white oak to climate and air pollution near the Ohio River Valley. J Environ Qual 14:274–280
McLaughlin S, Percy K (1999) Forest health in North America: some perspectives on actual and potential roles of climate and air pollution. Water Air Soil Pollut 116:151–197
Mitchell HL, Chandler RF (1939) The nitrogen nutrition and growth of certain deciduous trees in northeastern United States. The Blackrock Forest bulletin no. 11. Cornwall-on-the-Hudson, NY
Moldan F, Wright FR (1998a) Changes in runoff chemistry after five years of N addition to a forested catchment at Gårdsjön, Sweden. For Ecol Manag 101:187–197
Moldan F, Wright FR (1998b) Episodic behaviour of nitrate in runoff during six years of nitrogen addition to the NITREX catchment at Gårdsjön, Sweden. Environ Pollut 102(S1):439–444
Moldan F, Hultberg H, Nyström U, Wright RF (1995) Nitrogen saturation at Gårdsjön, SW Sweden, induced by experimental addition of nitrogen. For Ecol Manag 71:89–97
Moldan F, Skeffington RA, Mörth C-M, Torssander P, Hultberg H, Munthe J (2004) Results from covered catchment experiment at Gårdsjön, Sweden, after ten years of clean precipitation treatment. WASP 154(1):371–384
Moldan F, Kjønaas JO, Stuanes OA, Wright FR (2006) Increased nitrogen in runoff and soil following 13 years of experimentally increased nitrogen deposition to a coniferous-forested catchment at Gårdsjön, Sweden. Environ Pollut 144:610–620
Nash TH, Fritts HC, Stokes MA (1975) A technique for examining non-climatic variation in widths of annual tree rings with special reference to air pollution. Tree-Ring Bull 35:15–24
Nihlgard B (1985) The ammonium hypothesis-an additional explanation to the forest dieback in Europe. Ambio 14:2–8
Nilsson L-O, Wiklund K (1992) Influence of nutrient and water stress on Norway spruce production in south Sweden-the role of air pollutants. Plant Soil 147:251–262
Nohrstedt H-G, Sikström U, Ring E (1993) Experiments with vitality fertilisation in Norway spruce stands in southern Sweden. The Forestry Research Institute of Sweden, report no. 2, Uppsala, p 38
Nöjd P, Reams AG (1996) Growth variation of Scots pine across a pollution gradient on the Kola Peninsula, Russia. Environ Pollut 93(3):313–325
Persson H, Ahlström K (1999) Effect of nitrogen deposition on tree roots in boreal forests. In: Rastin N, Bauhus J (eds) Going underground—ecological studies in forest soils. Research Signpost, Trivandrum, pp 221–238
Persson O, Eriksson H, Johansson U (1995) An attempt to predict long-term effects of atmospheric nitrogen deposition on the yield of Norway spruce stands (Picea abies (L.) Karst.) in southern Sweden. Plant Soil 168(169):249–254
Persson H, Ahlström K, Clemensson-Lindell A (1998) Fine-root response to nitrogen addition and removal at Gårdsjön—results from ingrowth cores. For Ecol Manag 101:199–205
Pettersson F (1994) Predictive functions for impact of nitrogen fertilization on growth over five years. The Forestry Research Institute of Sweden, report no. 3, Uppsala, p 56
Sikström U (1997) Effects of low-dose liming and nitrogen fertilization on stemwood growth and needle properties of Picea abies and Pinus sylvestris. For Ecol Manag 95:261–274
Stuanes AO, Kjønaas OJ, van Miegroet H (1995) Soil solution response to experimental addition of nitrogen to a forested catchment at Gårdsjön, Sweden. For Ecol Manag 71:99–110
Sutherland EK, Martin B (1990) Growth response of Pseudotsuga menziestii to air pollution from copper smelting. Can J For Res 20:1020–1030
Vanguelova EI, Hirano Y, Eldhuset TD, Sas-Paszt L, Bakker MR, Püttsepp Ü, Brunner I, Lõhmus K, Godbold D (2007) Tree fine root Ca/Al molar ratio—indicator of Al and acidity stress. Plant Biosyst 141:460–480
Wallace PZ, Lovett MG, Hart EJ, Machona B (2007) Effects of nitrogen saturation on tree growth and death in a mixed-oak forest. For Ecol Manag 243:210–218
Wright RF, van Breemen N (1995) The NITREX project: an introduction. For Ecol Manag 71:1–5
Wright RF, Brandrud T-E, Clemensson-Lindell A, Hultberg H, Kjönaas J, Moldan F, Persson H, Stuanes AO (1995) NITREX project: ecosystem responses to chronic additions of nitrogen to a spruce-forested catchment at Gårdsjön, Sweden. Ecol Bull 44:322–334
Wright RF, Alewell C, Cullen JM, Evans CD, Marchetto A, Moldan F, Prechtel A, Rogora M (2001) Trends in nitrogen deposition and leaching in acid-sensitive streams in Europe. Hydrol Earth Syst Sci 5(3):299–310
Zang U, Lamersdorf N, Borken W (2011) Response of the fine root system in a Norway spruce stand to 13 years of reduced atmospheric nitrogen and acidy input. Plant Soil 339:435–445
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by A. Merino.
Rights and permissions
About this article
Cite this article
Seftigen, K., Moldan, F. & Linderholm, H.W. Radial growth of Norway spruce and Scots pine: effects of nitrogen deposition experiments. Eur J Forest Res 132, 83–92 (2013). https://doi.org/10.1007/s10342-012-0657-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-012-0657-y