
Abstract
Uneven-aged silver fir-European beech forest stands were studied to (1) analyse the dynamics of diameter structure and tree species composition in the past two centuries and (2) determine the impact of red deer on the regeneration and recruitment of silver fir. The study used current data on forest stands, archival data from old forest management plans for the period 1789–2004, and red deer harvesting records for the period 1907–2006. During the observation period, the silver fir population aged and silver fir and European beech alternated in dominance. The study revealed a strong impact of red deer on the composition and recruitment of tree regeneration, especially on silver fir regeneration. The drastic changes in red deer density (from extermination up to 5.8 animals km−2) and past forest management practices were apparently the main factors driving the population dynamics of silver fir (regeneration, recruitment, and diameter structure) in the study area during the past two centuries.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The composition and structure of forest stands are a result of the complex interplay between growth conditions, natural and anthropogenic disturbances (Oliver and Larsen 1996), and inter- and intra-specific relationships (van der Maarel 2005). Natural disturbance—an important driver of stand dynamics—may be one-off events such as windthrows, fires, avalanches, and floods, or continuous events such as the impact of ungulate browsing on tree species regeneration (Pickett and White 1985). One of the key anthropogenic disturbances is forest management, which has a profound impact on the structural and spatio-temporal dynamics of forest stands.
Over the last centuries in Europe, forest management has caused large-scale changes in the spatial distribution, tree species composition, and structure of forest stands (Johann 2007). In the eighteenth and nineteenth centuries, even-aged forestry created large areas of uniform, mainly conifer-dominated forest stands. On the other hand, the application of uneven-aged systems in the nineteenth and the twentieth centuries (Diaci 2006; Johann 2007) has maintained mixed, more structurally diverse, and heterogeneous forest stands. Uneven-aged forest stands are considered stable in terms of structure and composition (e.g. Larsen 1995).
Selection management (Schütz 2001) is a form of uneven-aged forest management and is characterised by continuous forest cover, negligible fluctuations in growing stock and tree species composition, and continuous natural regeneration. Selection forests are widespread, particularly in Switzerland, Germany, France, Italy, Slovenia, and Croatia; they are often mountainous forests of silver fir (Abies alba Mill.), European beech (Fagus sylvatica L.), and Norway spruce (Picea abies (L.) Karst.). Silver fir is frequently the key tree species, and without it, the maintenance of selection structures of forest stands is less successful (Korpel 1995; Schütz 2001). The most important conditions for the maintenance of a selection structure are ongoing regeneration and the recruitment of smaller-diameter trees into larger diameter classes. Silvicultural measures in particular have a profound impact on these conditions, but other factors may be important as well. For example, the selective browsing of large ungulates on regeneration and differences in how tree species recover from browsing co-determine the species composition of regeneration and reduce the recruitment of the most palatable tree species (Gill 1992b).
Due to the palatability and slow juvenile growth of silver fir, and consequently, the longer exposure to browsing, fir is one of the most susceptible European tree species to the impact of large herbivores. In some areas in Central Europe, such as in the Alps and in the Dinaric Mountains, silver fir regeneration may be browsed to the extent that it never passes the germination phase (Ott 1989; Motta 1996; Senn and Suter 2003; Jarni et al. 2004; Heuze et al. 2005). Many researchers emphasise that large herbivores have been a factor driving the profound changes in tree species composition of Central European forests since the beginning of the twentieth century (Gill 1992a; Ammer 1996; Putman 1996). The impact of browsing has varied as the population densities of large herbivores changed over space and time. A frequent obstacle to arriving at more reliable findings about the long-term impact of large herbivores on the development of forest stands has been a lack of research based on long-term datasets. Senn and Suter (2003) established that most existing research provides only indirect evidence of the impact of large herbivores on the long-term dynamics of forest stands and silver fir populations. Moreover, at the landscape level, there are very few studies of the long-term dynamics of selection forest stands in Central Europe (e.g. Gaspersic 1967; Boncina et al. 2003; O’Hara et al. 2007).
Long-term dynamics of forest stands are studied with a variety of methods: (1) palynological methods (e.g. Moore et al. 1997) make it possible to study tree species composition over several millennia; (2) dendrochronological methods (e.g. Schweingruber 1996; Swetnam et al. 1999) allow the reconstruction of the development of trees for periods spanning several centuries; and (3) archival data such as forest management plans, forest maps, land registers, felling records, and game harvesting records, which are a neglected source of information, make it possible to quantify long-term changes in forest structure and to better understand the impacts of a changing natural and anthropogenic disturbance regime over the past few decades or centuries (Axelsson et al. 2002; Chapman et al. 2006).
In Slovenia, regular forest management has a long and rich tradition (Boncina et al. 2003). Selection management, practised in 4% of Slovenian forests (Boncina et al. 2002), is well established in the Dinaric Mountains (Hufnagl 1893; Schollmayer 1906), an area that is home to a large, continuous complex of silver fir-European beech forests. These forests are a suitable object for the study of long-term changes of forest stands: detailed data on the structure and composition of forest stands and felled timber are available for a period spanning more than a century. The area is also suitable for studies of the impact of large herbivores on the development of forest stands: the population density of red deer (Cervus elaphus L.), the most widespread and ecologically the most significant species of large herbivore in the area (Jerina 2006), has changed dramatically over the past two centuries. Preserved archives of the annual harvest of red deer make it possible to estimate its population dynamics with sufficient accuracy. Additionally, the browsing of regeneration has been regularly monitored there for more than 30 years in both fenced and non-fenced areas. The aim of this study was thus (1) to study changes in diameter structure and tree species composition of uneven-aged selection silver fir-European beech forests over the last two centuries and (2) to determine the impact of red deer on regeneration and changes in diameter structure of silver fir.
Methods
Study area
The Dinaric Mountains stretch from Slovenia through Croatia and Bosnia and Herzegovina to Montenegro and Albania. The Dinaric silver fir-European beech (hereafter fir-beech) forests form one of the largest forest areas in Central Europe; in Slovenia, these forests represent 14% of the total forest area (Boncina et al. 2002). There is a long tradition of forest and wildlife management in these forests; the first forest management plans were made in the eighteenth century. A concept of nature-based forestry with single-tree and group selection as the prevalent silvicultural systems has been used from the beginning of the twentieth century, when adaptive forest management similar to Biolley’s concept in Switzerland began (Hufnagl 1893; Schollmayer 1906). Earlier, in the eighteenth and nineteenth centuries, exploitation involved mainly broadleaves, which were used for the production of charcoal and potash; the intensity of exploitation peaked in the second half of the nineteenth century (Perko 2002).
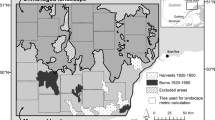
The dynamics of forest stands were studied in the forest management unit (FMU) Leskova dolina (Fig. 1), which is located in southern Slovenia (45°36′N; 14°28′E) and comprises 2,456 ha of uneven-aged fir-beech forests divided into 131 compartments. Elevation ranges between 740 and 1,350 m above sea level. The area has abundant precipitation (2,166 mm year−1), which is evenly distributed through the year. The average annual temperature is 6.5°C, with a mean monthly maximum temperature of 16.4°C in July and a mean minimum of −3.4°C in January. Late spring and early autumn frosts are common, and the snow cover duration averages 90 days (122 days maximum); the area is often not snow-free until late spring. The bedrock is carbonate, mainly limestone and some dolomite. Eutric cambisols of various depths developed on this bedrock, and in some parts, there are rendzinas of various depths. Fir is the dominant species, accounting for 50% of the total growing stock, followed by beech (26%), spruce (18%), and sycamore (Acer pseudoplatanus L.) with 2%. Other tree species (elm Ulmus glabra L., small-leaved lime Tilia cordata Mill., rowan Sorbus aucuparia L., and yew Taxus baccata L.) altogether account for less than 4% of the total growing stock. The average growing stock is 428 m3 ha−1, and the annual increment is 9.3 m3 ha−1 (FMP 2004). Forest stands in every compartment have undergone selection cutting approximately once per decade with an average cutting intensity of 6.9 m3 ha−1.

Intensive (2,456 ha) and extensive (approx. 18,000 ha) study area of FMU Leskova dolina with locations of fenced areas (locations A) and compartments where ages of mature silver firs were determined (locations B)
The study area is home to three species of large herbivores: red deer, roe deer (Capreolus capreolus L.), and chamois (Rupicapra rupicapra L.). The forest cover of the area is high (94%), and there are few pastures, which makes for considerable browsing pressure on forest vegetation (Jerina 2008). In the extensive study area (Fig. 1), red deer density is relatively high (about 3 animals km−2; this study), whereas the density of roe deer (1.3 km−2) and chamois (0.04 km−2) is low (Jerina unpublished). However, in spatial terms, ungulate densities in the extensive study area are heterogeneous: roe deer density is the highest at the foot of the hills, where the forest cover is lower; chamois is found only on peaks in the southern section of the extensive study area; and red deer are the dominant species in the intermediate zone, where fir-beech forests abound. This intermediate fir-beech zone is also where the tree regeneration sampling plots were located in our study. For most of the Holocene up to the March revolution in 1848, red deer was probably a common species (Fabjan 1956; Adamic 1992; Jerina 2006). After 1848, it disappeared due to over-harvesting, and in 1907, it was reintroduced (Adamic 1992). Its density did not start growing rapidly again until after World War II, reaching a maximum in 1976 (5.8 km−2), whereupon it halved in just a few years as intense shooting was mandated to reduce browsing damage to forests; since then, red deer density has fluctuated around this value (2005: 3 km−2) (Jerina 2006; this study).
Archival data acquisition and database creation
The dynamics of forest stands in the period 1789–2004 were studied on the basis of archival data acquired from the land register and old forest management plans kept in the Archives of the Republic of Slovenia and at the Slovenia Forest Service (Table 1). The oldest data on forests in the FMU Leskova dolina were acquired from the Josephinian land register of 1789. Detailed analysis was carried out on data from eight preserved forest management plans (FMP) for the study area, which were made in 1864, 1912, 1936, 1954, 1964, 1974, 1994, and 2004. The plans include the results of forest inventories at the level of compartments (N = 131; average compartment size = 18.75 ha). The intra-compartment boundaries hardly changed between 1864 and 2004, but the methods of forest inventories did. In 1789 and 1864, ocular assessments of the tree species composition of compartments were made. Most subsequent inventories involved full callipering of forest compartments (FMP 1912, 1936, 1954, 1964, 1974). All trees above a diameter at breast height (dbh) of 8 cm (FMP 1912), 10 cm (FMP 1954, 1964, 1974) or 12 cm (FMP 1936) were measured. The number of measured trees was high (e.g. 1,339,611 trees were measured in 1954). In the last two decades, the inventory of forest stands was carried out on permanent sampling plots (grid 200 × 250 m; area = 500 m2; N = 486), where trees ≥10 cm in dbh were measured.
The structure of the forest stand data is not entirely consistent. Typically, the inventory of a forest compartment includes information on the dbh structure (number of trees per dbh class and tree species), tree species composition (share of individual tree species in total growing stock), growing stock (m3 ha−1), and annual increment (m3 ha−1); data for 1789 and 1864 are an exception because they include only assessments of tree species composition in forest stands. In the 1912 and 1936 inventory, data on the dbh structure of forest stands by compartment are not shown for individual species but only aggregated for broadleaves and conifers. Inventory data on the number of trees are provided in non-consistent diameter classes. For 1912 and 1936, the trees were classified into 10-cm dbh classes (FMP 1912, 1936) and for all other years in 5-cm dbh classes (FMP 1954, 1964, 1974, 1994, 2004); data were therefore combined and shown on a single scale—in 10-cm dbh classes. The growing stock of individual forest tree species and forest stands was calculated with Biolley tariffs. Additional useful information was acquired from the text in the FMPs.
Analysis of age of mature silver fir trees
To examine the age and period of unsuppressed growth of mature fir trees in the area covered by this study, 221 mature fir trees (dbh ≥ 40 cm) in five randomly chosen compartments (Fig. 1) were analysed; this analysis was done in the field on freshly cut stumps.
Reconstruction of population dynamics of red deer
The population dynamics of red deer were reconstructed on the basis of data on harvest and loss (found dead animals, roadkill etc.), which were collected for the 1907–1938 period by Fabjan (1956); after 1948, these data were gathered systematically according to law in the official hunting registers. The quality of the available data changed over time: after 1976, data on the sex and age of extracted animals have been gathered, but previously only the data on total annual harvest were collected; during and for a short period after World War I (1914–1918) and World War II (1940–1946), hunting statistics were not kept. Accordingly, the dynamics of the population density were reconstructed with multiple methods: (1) for the period after 1976, with the “population reconstruction” method (see Roseberry and Woolf 1991); this method is based on the number and age of harvested animals, which are then used to reconstruct the number of animals that lived in a specific year. For example, for year X, records show the harvest of N animals aged 0+, in year X + 1, M number of animals aged 1+, in year X + 2, P number of animals aged 2+, etc. In year X, the number of animals aged 0+ was thus N + M + P + etc.; the same method is used to calculate the number of animals aged 1+, 2+, etc. for the individual years and the number of all animals in a given year; (2) for the period 1907–1976, the densities were reconstructed with a multivariate linear regression model which was build based on the data for 1976–2007 (in the model estimated, red deer density is the dependent variable and red deer harvest data are the independent variables); the model forecasts density in year X on the basis of a known number of extractions in year X and several years thereafter; and (3) for the years 1914–1918 and 1940–1946, the numbers were reconstructed with linear interpolation of density values before and after this period.
Browsing on regeneration
To determine the impact of red deer on the composition of the regeneration (dbh < 10 cm), the regeneration in two fenced (one 19 ha and one 2 ha; locations in Fig. 1) and two non-fenced areas was analysed in 2008. The fences were built 35 years ago and have been under permanent control by local professional foresters since then. Stand conditions are similar within and outside the fenced areas: they are dominated by stands with small canopy gaps and a growing stock of about 450 m3 ha−1. A total of 66 plots of 16 m2 were surveyed, which included 33 plots within both fenced areas and 33 outside the fenced areas; the approach (size and number of plots) was adopted by Boncina (2000) and Jarni et al. (2004). The location of the first randomly selected plot within each fenced area defined the origin of the transect that ran parallel to the fence. Plots were placed at 4-m intervals along the transects. Transects outside the fenced areas were parallel to the ones inside and placed at a similar distance from the fence as the transects inside (approximately 50 m from a fence). On each plot, seedlings (h < 130 cm) and saplings (h ≥ 130 cm and dbh < 10 cm) were recorded for each tree species and classified by height class (<20 cm; 20–49 cm; 50–89 cm; 90–129 cm; ≥130 cm and dbh < 10 cm). Additionally, browsing damage of the terminal shoot of each tree was recorded. The differences in the number of seedlings and saplings by tree species between fenced and non-fenced areas were examined using the non-parametric Mann–Whitney U test, since not all of the data were distributed normally (Hollander and Wolfe 1999).
Browsing of fir seedlings for the period 1977–2004 was analysed on the extensive area of FMU Leskova dolina (Fig. 1) based on records from permanent sampling plots. Eight consecutive seedling inventories in 1977, 1981, 1983, 1985, 1987, 1989, 1992, and 1994 were carried out on 147 sampling plots, while three consecutive inventories (1996, 2000, 2004) were carried out on 64 sampling plots (e.g. Veselic 1991; Jerina 2008). On 5 × 5 m plots, all seedlings were tallied by tree species and height class (<15 cm; 15–29 cm; 30–129 cm) and recorded as browsed or unbrowsed. Only summary data (for all plots combined) are available for the first inventory. Consequently, the analysis always considered summary data for individual years. Because variation in red deer density may potentially affect both browsing intensity and fir seedling density, both indicators were analysed. A correlation between browsing intensity and red deer density, and seedling density and red deer density was estimated with the non-parametric Kendall’s tau-b correlation analysis; its advantage is in the sensitivity to all monotonous and not just linear correlations between variables, and it does not presume a normal distribution of variables (Hollander and Wolfe 1999).
Results
Long-term forest stand dynamics
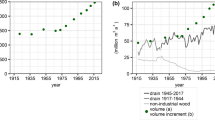
The tree species composition of uneven-aged fir-beech forests in the FMU Leskova dolina underwent profound change during the 1789–2004 period (Table 2). At the end of the eighteenth century, broadleaves accounted for the bulk of the growing stock (76%). Conifers made up 24% of the growing stock and were dominated by fir; few spruce trees were present. By the middle of the nineteenth century, the composition of the forest stands changed substantially: the share of conifers, particularly fir, rose and the share of beech and other broadleaves fell. This trend continued until 1974, when conifers reached the highest share of the growing stock (80%) and fir accounted for as much as two-thirds of the total growing stock. After 1974, the share of broadleaves, especially beech, increased notably, and the share of conifers dropped. Among the conifers, the share of fir in total growing stock dropped from 68 to 50% by 2004. Spruce was very rare in the stands in 1912, whereupon its share of the total growing stock rose continually, doubling between 1954 and 2004.
At the beginning of the twentieth century, the growing stock of the forest stands was low (234 m3 ha−1) due to heavy exploitation at the end of the nineteenth century. After 1912, the total growing stock increased, by as much as 83% by 2004.
Data on tree species composition and total growing stock of the forest stands indicate an ageing of the stands and an alternation between fir and beech; this has been additionally confirmed by an analysis of the dbh structure of the forest stands. In the period 1912–2004, the dbh structure of the forest stands maintained a reversed J-shaped distribution (Fig. 2a). However, significant changes are evident: the number of medium-sized (dbh = 30–49 cm) and large-sized (dbh ≥ 50 cm) trees increased and the number of small-sized trees (dbh = 10–29 cm) dropped constantly.

Diameter structure dynamics of forest stands a and populations of individual tree species, b silver fir, c European beech, and d Norway spruce
The largest changes occurred in the dbh structure of fir (Fig. 2b). In 1912, there were hardly any firs equal to or more than 50 cm in dbh (4 ha−1), but their number grew to 20 trees per ha by 1954, and in 2004, there were 36 large-sized diameter firs per ha. Between 1912 and 2004, the number of small-sized diameter firs (dbh = 10–19 cm) dropped drastically from 190 to 30 ha−1, respectively. The results suggest a successive ageing of the fir population coupled with insufficient recruitment of young firs to the stand canopy (see also Fig. 3b).

a Correlation between red deer density and browsing of silver fir seedlings, 1977–2004; b comparison of red deer density in the period 1900–2006 and number of small-sized diameter silver firs (dbh = 10–19 cm) in studied forest stands
Half of the current generation of mature firs (dbh ≥ 40 cm) germinated in a relatively short period between 1829 and 1846 (Table 3). The period of unsuppressed growth of the studied firs started at the end of the nineteenth century.
The dbh structure of beech in the period 1912–2004 (Fig. 2c) did not indicate such significant changes as that of fir; in the last two decades, the number of large- and medium-sized diameter beeches increased substantially, resulting in a notable increase in its share in the total growing stock (Table 2). The dbh structure of spruce (Fig. 2d), meanwhile, shows a gradual increase in the number of smaller-sized diameter trees in the period 1974–2004, indicating that spruce is being recruited into the stands.
The impact of red deer on regeneration, recruitment, and diameter distribution dynamics of silver fir
Analysis of browsing on regeneration in fenced and non-fenced areas in 2008 showed that selective browsing by large herbivores had a major impact on the density and species composition of the regeneration. No statistically significant differences were found between the fenced and non-fenced areas for total regeneration density (P = 0.078), but there were statistically significant differences in the density of seedlings and saplings in some height classes (Table 4). Statistically significant differences between the two areas were also found for the density of fir regeneration (P = 0.000); in the fenced areas, the density of fir regeneration was 4.8 times higher than in the non-fenced areas. The differences are even more obvious by height classes: no statistically significant differences were found for the density of fir seedlings up to 20 cm in height, but in the 20- to 49-cm class, there were 50 times more fir seedlings in fenced areas; no fir seedlings higher than 50 cm or fir saplings were registered in non-fenced areas. Similar results were found for sycamore: in fenced areas, the number of sycamore seedlings up to 20 cm high and seedlings in the 20- to 49-cm class was significantly lower than in non-fenced areas (P = 0.000 and P = 0.000, respectively), but the recruitment of sycamore seedlings into higher classes was noticeably higher in the fenced areas. The density of beech regeneration was higher in non-fenced than in fenced areas, but the difference was not statistically significant (P = 0.087). Spruce was hardly registered at all in the regeneration layer.
In the years when records of seedling (h < 130 cm) browsing in the extensive area of FMU Leskova dolina were made (1977–2004), the red deer density changed significantly (Fig. 3a). The highest density was recorded in 1977 (5.8 km−2) and the lowest 10 years later, in 1987 (2.5 km−2). The density and browsing of fir seedlings are strongly related to their height. The average density of all fir seedlings in one inventory in 1977–2004 was nearly 24,000 individuals ha−1, the browse rate (the rate of browsed individuals among all individuals) was 8.2%, and fir’s share of total seedlings (the share of seedlings of one tree species in the total number of seedlings) was over 25%. The average density of fir seedlings over 15 cm was 314 individuals ha−1, the browse rate exceeded 38%, and the share of fir in the seedlings of the same height category dropped to 1.6%. Finally, only an average of 44 firs ha−1 was registered in the highest class (30–129 cm), of which 71% on average were browsed, and the share of fir in total seedlings above 30 cm dropped to only 0.6%. In 4 of the 11 monitoring years, all firs above 30 cm were browsed; in one census (in 1994), there were no firs over 30 cm at all. The signs of correlations (Table 5) between the density and browsing intensity of fir seedlings and red deer density were mostly in accordance with expectations, but only the correlation between the browsing rate to all fir seedlings and red deer density was statistically significant (r = 0.49; P = 0.036; n = 11).
Recruitment of fir into small-diameter trees dropped constantly over the studied period (Fig. 3b). The number of small-sized diameter firs (dbh = 10–19 cm) in the forest stands was significantly higher in 1912 than in 1954 or 2004, at 190, 111, and 30 trees, respectively. A similar trend was evident in the share of fir in the total number of small-sized diameter trees (results not shown); both started to drop before the red deer density began to rapidly increase.
Discussion
Archival data enables detailed reconstructions of the development of forest stands and the factors that affect this development (Axelsson et al. 2002; Chapman et al. 2006). However, cautious analysis is indispensable when data from different archive sources are used, because differences between data sets are easily confused with structural changes of forest stands (Radeloff et al. 1999; Agnoletti and Anderson 2000). Archival data describe the state of forest stands at a certain point in time. This is just one of many—albeit perhaps noteworthy different—states in time over the long-term development of forest stands, a fact which needs to be considered in the interpretation of the results. However, with a large enough density of such points (which archival data enable), it is possible to clearly describe the dynamics of forest stands over a long period.
In terms of the data quality on stand parameters and the length of the period the data cover, this study area is one of the best in Slovenia, and the data are of similar quality as in other similar studies (e.g. Linder and Östlund 1998; Axelsson et al. 2002; Montes et al. 2005). Nevertheless, some problems appeared in the acquisition of data and the preparation of the database on the stand parameters: in some inventories, the data were incomplete; the tree species composition was provided by groups of tree species; different inventory methods were used; and the dbh structures of the stands were provided in variously broad dbh classes.
We only examined the influence of red deer on regeneration, although roe deer and chamois also live in the study area. In the extensive study area, population densities, and in particular, biomass (as better indicator of ecological impacts) of roe deer and chamois are very low per se, especially when compared to red deer. The maximum biomass of red deer in the period 1976–2005 was 381 kg km−2, whereas that of roe deer and chamois was 37 and 1.7 kg km−2, respectively (Jerina unpublished). Moreover, the spatial distribution of the three ungulate species in the study area is very spatially heterogeneous: roe deer live mostly in the lowest-lying, less-forested areas; chamois at the highest altitudes; while the intermediate zone, where the sampling plots were located, provides ideal conditions for red deer. Consequently, the density and biomass estimation were underestimated for red deer and overestimated for roe deer and chamois. As a result, estimates of roe deer and chamois density dynamics would likely be too imprecise for use in the analysis of regeneration browsing.
The methods used to reconstruct the red deer density presume that all mortality was registered, but this is never entirely true. However, the reintroduction of red deer to the study area in 1907 was the first successful reintroduction of the species in the wider region (Adamic 1992) and therefore the subject of considerable and constant attention among both resource managers and hunters. Consequently, the data were gathered very meticulously, and there was no poaching because the area was managed by professional hunters. Additionally, due to problems with forest regeneration, foresters have been continuously pushing hunters to reduce the red deer density. Under Slovenian legislation, registered non-hunting mortality is considered as part of the total annual harvest. Because hunters did not agree with a strong reduction in red deer density, they were motivated to record non-hunting mortalities. In the study area, non-hunting mortality occurs mainly due to large predators [grey wolf (Canis lupus L.) and Eurasian lynx (Lynx lynx L.)], and only a portion of this mortality is registered (e.g. Smith et al. 2004; Krofel et al. 2008; Webb et al. 2008). But even if this fact was taken into account, the total estimated population size would not be significantly affected, as large predator-caused mortality represents a very low share of the total harvest (<10%) (Jerina unpublished). Furthermore, the study area is located in a forested mountain massif which is surrounded on most sides with non-forest areas, which makes the red deer population demographically very isolated. All the aforementioned factors are important because our methods assume no (or equalized) emigration and immigration and a 100% detection of dead animals (Roseberry and Woolf 1991). The calculated population densities were thus slightly, but constantly underestimated.
The population density of red deer fluctuated greatly during the study period, and therefore the minor errors in the red deer data should not decrease the ability to detect the impact of red deer on vegetation change. Furthermore, we were only concerned with the relative dynamics of the population parameters rather than absolute values. Therefore, we believe our data were appropriate for examining the research objectives.
Changes in composition and structure are a fundamental part of the natural dynamics of forest stands (Pickett and White 1985; Oliver and Larsen 1996). Management and other anthropogenic influences can only mitigate or intensify them. A comparison of studies based on archival data (e.g. Linder and Östlund 1998; Radeloff et al. 1999; Axelsson et al. 2002; Duchesne et al. 2005; Chapman et al. 2006; O’Hara et al. 2007; Vrska et al. 2009) reveals a variety of forest stand dynamics in different forest types and a wealth of leading impact factors, but a frequent common characteristic is the fluctuation in tree species composition and dbh structure. The traditional view is that composition and structure of selection forests do not alter substantially over a long period of time, yet this study concluded the opposite: during the 215-year observation period of fir-beech forest dynamics, the stand parameters changed a considerable degree.
Substantial changes in growing stock and dbh structure of forest stands were established in the observed period. The total growing stock of the stands increased as a result of a deliberate accumulation of volume increment (FMP 1912, 1954). Consequently, the number of large-sized diameter trees in the dbh structure of the forest stands rose. Boncina et al. (2003) found similar changes in other uneven-aged fir-beech forests in Slovenia. Those changes were the result of the first intensive forest exploitation at the end of the nineteenth century, which reduced the growing stock of forest stands, and the subsequent deliberate increase in growing stock of uneven-aged stands towards target values of about 400 m3 ha−1 (FMP 1954; Gaspersic 1967). Similar changes were observed in Croatian Dinaric fir-beech forests (Cavlovic 2000), and analogous, albeit somewhat smaller, changes occurred also in Swiss plenter spruce-fir-beech stands in past 6–9 decades (O’Hara et al. 2007); this indicates similar dynamics of uneven-aged forests in Slovenia, Croatia, and Switzerland, and probably Central Europe in general.
The tree species composition of the studied forest stands changed drastically between 1789 and 2004. Fir underwent the greatest change, as its share of the total growing stock grew strongly at first, but then dropped in the final decades, from 68% in 1974 to 50% in 2004. At the same time, the share of beech and spruce in the total growing stock increased. Their dbh structures indicate a continued increase of their share in the forest stands over the coming decades. These findings correspond to the process of the alternation of dominant tree species (e.g. Simak 1951; Rabotnov 1992; Korpel 1995), yet our data suggest that this alternation is more a result of anthropogenic influences rather than a natural endogenous process; similar was found in the research in fir-beech forests of the Carpathians (Vrska et al. 2009). Furthermore, the study period is too short to reach a definite conclusion about fir-beech alternation with certainty. Nevertheless, fir has been reported to be frequently replaced by beech (e.g. Gaspersic 1974) and spruce (e.g. Heuze et al. 2005). Similar changes in the composition of fir-beech forests have also been documented in virgin forest remnants (e.g. Korpel 1995; Boncina 1999; Boncina et al. 2003; Diaci 2006), but the magnitude of changes was smaller than in our study area. The drop in the fir share in the growing stock of mixed forests and even a reduction in its distribution have been reported by researchers across Central Europe (e.g. Schütt et al. 1999; Senn and Suter 2003; Ficko and Boncina 2006).
Insufficient recruitment of fir into the stand canopy is the main impediment for the practice of selection forest management in the study area. This problem has been reported in many Central European countries (e.g. Motta 1996; Senn and Suter 2003; Heuze et al. 2005; Cavlovic et al. 2006). Of the factors that affect fir regeneration and recruitment in the study area, two factors stand out, both of them directly or indirectly triggered by man: (1) forest management and (2) ungulate browsing on regeneration.
From the middle of the nineteenth century to the middle of the twentieth century—a period of functionally negligible ungulate population densities—forest management was the main impact factor in the dynamics of the studied forest stands. Fir was present in the study area before the period covered by this study (Sercelj 1996) but the mature firs growing today germinated in the first half of the nineteenth century. The first major exploitation of these forests, in the second half of the nineteenth century, considerably reduced the growing stock of the stands and increased the light influx into the stands, which is reflected in the released growth of fir regeneration. In line with the economic principles of that time, foresters promoted conifers, in particular fir, in regeneration and among trees by weeding out beech (Perko 2002). At the beginning of the twentieth century, plenter management was introduced, but owing to the stands’ origin, they did not have an entirely satisfactory plenter structure (Schollmayer 1906); by removing beech, foresters continued to promote conifers. The bulk of medium-sized diameter trees—mostly fir—in the growing stock of the forest stands and the planned accumulation of volume increment (Perko 2002) slowed down regeneration and recruitment of small-sized diameter trees into the stand canopy. In the first decades of the twentieth century, attempts were made to accelerate recruitment of fir by creating small gaps in stand canopy, but such conditions were more favourable for the regeneration and recruitment of broadleaves (Gaspersic 1967). The fact is that beech successfully germinated in stands dominated by fir, which regenerated abundantly in the mid-nineteenth century, when beech dominated the top layer. This strengthened a belief in the alternation of dominant tree species, ostensibly because of changes in soil conditions caused by tree litter (Gaspersic 1974; Pintaric 1978). In the period 1912–2004, the number of large-sized diameter trees, in particular fir, increased significantly, which indicates the ageing of forest stands and the fir population.
The ageing of fir-beech stands due to an insufficient regeneration and recruitment rate of fir has also been reported in Croatia (Cavlovic et al. 2006). A lack of regeneration and recruitment, and population ageing, has also been recorded for other species, including eastern hemlock (Tsuga canadensis (L.) Carr.) in mixed forests of Michigan, in the USA (Frelich and Lorimer 1985). A reduction in recruitment of small-sized diameter firs to the canopy of the analysed forest stands was recorded already in the period of low red deer density (≤0.1 km−2) at the beginning of the twentieth century. This suggests that some additional factors other than red deer browsing also affect fir recruitment, such as the structure and composition of forest stand, the density of the stand canopy, alelopathic relation between tree species, etc. (e.g. Gaspersic 1967; Pintaric 1978; Prpic et al. 2001).
The impact of large herbivores on forest stand dynamics varied significantly over time. After 1848, when the March revolution changed social relations and legal conditions also regarding hunting rights, red deer was completely exterminated (Fabjan 1956; Adamic 1992), which had a favourable effect on fir regeneration. The first exploitation of the studied forest stands, at the end of the nineteenth century, increased the food supply of shrubs, herbs, and grasses. Consequently, roe deer, which had then been present at very low densities, apparently did not have any noteworthy impact on tree regeneration, including the most sensitive species such as fir. In 1907, red deer was reintroduced to the study area, and after 1950, through a combination of favouring red deer in wildlife management, a low harvesting rate, and an absence of large predators, the population density started growing exponentially. The impact of red deer on vegetation, including tree seedlings, increased dramatically. In the 1980s and 1990s, browsing pressure was singled out by the forest service as the main problem of forest management in this area. After 1976, culling reduced the red deer population from 5.8 to about 3 km−2, but the browsing rate was still high and recruitment into the stand canopy weak (Veselic 1991); obviously the great change in red deer density did not reduce damage to fir seedlings, although a drop in damage to sycamore and beech, for example, was registered (Debeljak et al. 1999).
Like elsewhere in mountainous mixed forests (e.g. Ott 1989; Motta 1996; Senn and Suter 2003; Heuze et al. 2005), in fir-beech forests, fir is one of the most susceptible species to browsing. The question that arises is at what densities of ungulates can fir still be successfully recruited into the stand canopy. A Slovenia-wide study (Jerina 2008) showed that relationships between large herbivore density and degree of fir browsing are explicitly non-linear and weak; a greater drop in the degree of browsing was recorded only for exceptionally low herbivore densities (e.g. red deer <1 km−2), apart from that the intensity of fir browsing was constantly high. Present-day ungulate densities in the study area are generally not high (see Ammer 1996; Motta 1996; Heuze et al. 2005). However, the regeneration in fenced and non-fenced areas undeniably shows that red deer has a fundamental impact on regeneration and recruitment of fir and certain other tree species. The impact is even greater because the food capacity of the area is meagre due to a high forest cover (94%), high growing stock, and dense canopy.
Other factors may also be important for tree seedling recruitment and, consequently, changes in tree species composition, such as growing conditions, inter- and intra-specific relationships, general food availability in the forest stands, resistance of individual species to browsing, silver fir dieback, and climate change. (Gill 1992a, b; Debeljak et al. 1999; Petit and Lambin 2002; Bigler et al. 2004; van der Maarel 2005). Although these factors are complexly inter-related, they were not analysed in this study as it was presumed that they have a small impact compared to the studied factors. Some observations showed that on some sites with similar ungulate densities as in the study area, fir regenerates well (Jerina 2008), which means that the “target” density of ungulates that allows fir to regenerate and recruit to stand canopy varies depending on the other impact factors, among which inter- and intra-specific competition in combination with site and stand (light) conditions may play an important role. Just as Frelich and Lorimer (1985) found for eastern hemlock (Tsuga canadensis (L.) Carr.) in Michigan, USA, and Linder (1998) for Scots pine (Pinus sylvestris L.) and Norway spruce in Sweden, it is possible that fir needs a “window of opportunity” for successful recruitment into the stand canopy (Senn and Suter 2003)—a co-occurrence of appropriate conditions, including a low population density of large herbivores.
Forecasts of the future of fir in Dinaric fir-beech forests, and more broadly in Central Europe, are unreliable: this study and several others (e.g. Motta 1996; Heuze et al. 2005; Ficko and Boncina 2006; Jerina 2008) indicate that its share will continue to decline, probably to the level of the eighteenth century in our study area. The tree species composition before the analysed period cannot be completely determined; instead of hypothesizing a static ratio of tree species, it is more appropriate to understand it in the sense of “natural” fluctuations. Sercelj (1996) found that fir and beech have maintained a constant presence in fir-beech forests in Slovenia for the last 7,000 years, but, presumably due to changes in climate conditions, their relative dominance continuously changes. Anthropogenic impacts have mostly intensified these fluctuations, the extermination and reintroduction of red deer in the study area being such an example. Ungulate density (and its impact) likely went through changes before the period of our analysis as well, which may have contributed to past fluctuations in the tree species composition of forest stands.
Understanding the dynamics of the composition and structure of forest stands and the processes that drive these dynamics is an essential basis for present and future ecosystem-based forest management. Therefore, uneven-aged forest management should be more oriented towards managing the processes in forest ecosystems, and less towards maintaining static structures. Furthermore, uneven-aged forest management should not only be about stand management, since wildlife populations can have a strong influence on stand structure and composition.
References
Adamic M (1992) Red deer management (Cervus elaphus L.) in Republic Slovenia: the problems, recent trends and perspectives. Zbornik gozdarstva in lesarstva 39:47–60
Agnoletti M, Anderson S (eds) (2000) Methods and approaches in forest history. CABI Publishing, Wallingford
Ammer C (1996) Impact of ungulates on structure and dynamics of natural regeneration of mixed mountain forests in the Bavarian Alps. For Ecol Manag 88(1–2):43–53
Axelsson AL, Ostlund L, Hellberg E (2002) Changes in mixed deciduous forests of boreal Sweden 1866–1999 based on interpretation of historical records. Landscape Ecol 17(5):403–418
Bigler C, Gricar J, Bugmann H, Cufar K (2004) Growth patterns as indicators of impending tree death in silver fir. For Ecol Manag 199:183–190
Boncina A (1999) Stand dynamics of the virgin forest Rajhenavski Rog (Slovenia) during the past century. In: Diaci J (ed) Virgin forests and forest reserves in Central and East European countries: history, present status and future development: proceedings of the invited lecturers’ reports presented at the COST E4 management committee and working groups meeting in Ljubljana, Slovenia. Biotechnical faculty, Department of Forestry and Renewable Forest Resources, Ljubljana, pp 95–110
Boncina A (2000) Comparison of structure and biodiversity in the Rajhenav virgin forest remnant and managed forest in the Dinaric region of Slovenia. Glob Ecol Biogeogr 9:201–211
Boncina A, Diaci J, Cencic L (2002) Comparison of the two main types of selection forests in Slovenia: distribution, site conditions, stand structure, regeneration and management. Forestry 75(4):365–373
Boncina A, Gaspersic F, Diaci J (2003) Long-term changes in tree species composition in the Dinaric mountain forests of Slovenia. Forestry Chron 79(2):227–232
Cavlovic J (2000) Novi program gospodarenja za G.J. Belevine (2000–2009)—zaustavljanje nepovoljnih trendova i iniciranje povoljnih procesa u razvoju preborne sume? Sumarski list 124(7–8):450–457
Cavlovic J, Bozic M, Boncina A (2006) Stand structure of an uneven-aged fir-beech forest with an irregular diameter structure: modeling the development of the Belevine forest, Croatia. Eur J For Res 125:325–333
Chapman RA, Heitzman E, Shelton MG (2006) Long-term changes in forest structure and species composition of an upland oak forest in Arkansas. For Ecol Manag 236(1):85–92
Debeljak M, Dzeroski S, Adamic M (1999) Interactions among the red deer (Cervus elaphus L.) population, meteorological parameters and new growth of the natural regenerated forest in Sneznik, Slovenia. Ecol Model 121:51–61
Diaci J (2006) Gojenje gozdov: pragozdovi, sestoji, zvrsti, nacrtovanje, izbrana poglavja (in Slovene). Biotechnical faculty, Department of Forestry and Renewable Forest Resources, Ljubljana
Duchesne L, Ouimet R, Moore JD, Paquin R (2005) Changes in structure and composition of maple-beech stands following sugar maple decline in Quebec, Canada. For Ecol Manag 208(1–3):223–236
Fabjan I (1956) Snezniska jelenjad. Lovec 39(6):161–169
Ficko A, Boncina A (2006) Silver fir (Abies alba Mill.) distribution in Slovenian forests. Zbornik gozdarstva in lesarstva 79:19–35
FMP (1912) Betriebseinrichtung für das Dezennium 1912–1921. Reviere Schneeberg, Leskova dolina, Masun, Georgstal (in German)
FMP (1936) Forest management plan for FMU Leskova dolina 1936 (in Italian)
FMP (1954) Forest management plan for FMU Leskova dolina 1954–1963 (in Slovene)
FMP (1964) Forest management plan for FMU Leskova dolina 1964–1973 (in Slovene)
FMP (1974) Forest management plan for FMU Leskova dolina 1974–1983 (in Slovene)
FMP (1994) Forest management plan for FMU Leskova dolina 1994–2003 (in Slovene)
FMP (2004) Forest management plan for FMU Leskova dolina 2004–2013 (in Slovene)
Frelich LE, Lorimer CG (1985) Current and predicted long-term effects of deer browsing in hemlock forests in Michigan, USA. Biol Conserv 34(2):99–120
Gaspersic F (1967) Razvojna dinamika mesanih gozdov jelke-bukve na Snezniku v zadnjih sto letih (in Slovene). Gozdarski vestnik 7–8:202–237
Gaspersic F (1974) Zakonitosti naravnega pomlajevanje jelovo-bukovih gozdov na visokem krasu Sneznisko-Javorniskega masiva: doctoral dissertation (in Slovene). Biotechnical faculty, Department of Forestry and Renewable Forest Resources, Ljubljana
Gill RMA (1992a) A review of damage by mammals in north temperate forests. 1. Deer. Forestry 65(2):145–169
Gill RMA (1992b) A review of damage by mammals in north temperate forests. 3. Impact on trees and forests. Forestry 65(4):363–388
Heuze P, Schnitzler A, Klein F (2005) Consequences of increased deer browsing winter on silver fir and spruce regeneration in the Southern Vosges Mountains: implications for forest management. Ann For Sci 62(2):175–181
Hollander M, Wolfe DA (1999) Non-parametric statistical methods, 2nd edn. Wiley, New York
Hufnagl L (1893) Der Plenterwald, sein Normalbild, Holzvorrat, Zuwachs und Ertrag. Österr Vierteljahresschr Forstwes 117–132
Jarni K, Robic D, Boncina A (2004) Analysis of the influece of ungulates on the regeneration of Dinaric fir-beech forests in the research site Trnovec in the Kocevje forest management region. Zbornik gozdarstva in lesarstva 74:141–164
Jerina K (2006) Spatial distribution, home range and body mass of red deer (Cervus elaphus L.) in regard to environmental factors: doctoral dissertation (in Slovene). Biotechnical faculty, Department of Forestry and Renewable Forest Resources, Ljubljana
Jerina K (2008) Velika rastlinojeda divjad in razvojna dinamika gozdnih ekosistemov: proucevanje vplivov izbranih okoljskih in populacijskih parametrov ter gozdno-gojitvenih sistemov na zmoznosti naravne obnove: zakljucno porocilo o rezultatih opravljenega raziskovalnega dela na projektu v okviru ciljnega raziskovalnega projekta (CRP) “Konkurencnost Slovenije 2006–2013”: Research report. Biotechnical faculty, Department of Forestry and Renewable Forest Resources, Ljubljana
Johann E (2007) Traditional forest management under the influence of science and industry: the story of the alpine cultural landscapes. For Ecol Manag 249(1–2):54–62
Korpel S (1995) Die Urwälder der Westkarpaten. Gustav Fisher Verlag, Stuttgart
Krofel M, Kos I, Linnell J, Odden J, Teurlings I (2008) Human kleptoparasitism on Eurasian lynx (Lynx lynx L.) in Slovenia and Norway. Varstvo narave 21:93–103
Larsen JB (1995) Ecological stability of forests and sustainable silviculture. For Ecol Manag 73(1–3):85–96
Linder P (1998) Structural changes in two virgin boreal forest stands in central Sweden over 72 years. Scand J For Res 13(4):451–461
Linder P, Östlund L (1998) Structural changes in three mid-boreal Swedish forest landscapes, 1885–1996. Biol Conserv 85(1–2):9–19
Montes F, Sánchez M, del Rio M, Canellas I (2005) Using historic management records to characterize the effects of management on the structural diversity of forests. For Ecol Manag 207(1–2):279–293
Moore PD, Webb JA, Collinson ME (1997) Pollen analysis. Blackwell, London
Motta R (1996) Impact of wild ungulates on forest regeneration and tree composition of mountain forests in the Western Italian Alps. For Ecol Manag 88(1–2):93–98
O’Hara KL, Hasenauer H, Kindermann G (2007) Sustainability in multi-aged stands: an analysis of long-term plenter systems. Forestry 80:163–181
Oliver CD, Larsen BC (1996) Forest stand dynamics. Wiley, New York
Ott E (1989) Verjüngungsprobleme in hochstaudenreichen Gebirgsnadelwäldern. Schweizerische Zeitung Forstwes 140(1):23–42
Perko F (2002) Zapisano v branikah: Gozdovi in gozdarstvo od Sneznika do Nanosa skozi cas. Gozdarsko drustvo Postojna, Postojna
Petit CC, Lambin EF (2002) Long-term land -cover change in the Belgian Ardennes (1775–1929): model-based reconstruction vs. historical maps. Glob Change Biol 8:616–630
Pickett ST, White PS (1985) The ecology of natural disturbance of natural patch dynamics. Academic Press, San Diego
Pintaric K (1978) Urwald Perucica als natürliches Forschungslaboratorium. Allgemeine Forst Zeitschrift 24:702–707
Prpic B, Matic S, Vukelic J, Seletkovic Z (2001) Virgin forests of beech and fir in the Croatian Dinaric Mountain range. In: Prpic B (ed) Silver fir (Abies alba Mill.) in Croatia. Akademija sumarskih znanosti, Hrvatske sume, p.o. Zagreb, Zagreb, Croatia, pp 479–500
Putman RJ (1996) Ungulates in temperate forest ecosystems: perspectives and recommendations for future research. For Ecol Manag 88(1–2):205–214
Rabotnov TA (1992) Phytozönologie: Struktur und Dynamik natürlicher Ökosysteme. Verlag Eugen Ulmer, Stuttgart
Radeloff VC, Mladenoff DJ, He HS, Boyce MS (1999) Forest landscape change in the northwestern Wisconsin Pine Barrens from pre-European settlement to the present. Can J For Res 29(11):1649–1659
Roseberry JL, Woolf A (1991) A comparative evaluation of techniques for analyzing white-tailed deer harvest data. Wildlife monographs 117
Schollmayer H (1906) Direktiven für die Bestandesaufnahme und die Betriebseinrichtung auf der F.C. Herrschaft Schneeberg. Kleinmayr—Bamberg, Ljubljana
Schütt P, Weisberger H, Lang UM, Roloff A, Stimm B (1999) Abies alba Miller, 1768. In: Schütt P, Roloff A (eds) Enzyclopädie der Holzgewächse: Handbuch und Atlas der Dendrologie, 16. Ergänzungslieferung. Wiley-VCH, Weinheim
Schütz JP (2001) Der Plenterwald und weitere Formen strukturierter und gemischter Wälder. Parey, Berlin
Schweingruber FH (1996) Tree rings and environment. Dendroecology. Swiss Federal Institute for Forest, Snow Landscape Research, Birmensdorf, Paul Haupt Verlag, Berne
Senn J, Suter W (2003) Ungulate browsing on silver fir (Abies alba) in the Swiss Alps: beliefs in search of supporting data. For Ecol Manag 181(1–2):151–164
Sercelj A (1996) The origins and development of forests in Slovenia. Slovenska akademija znanosti in umetnosti, Ljubljana
Simak M (1951) Untersuchungen über den naturlichen Bauartenwechsel in der schweizerischen Plenterwälder. Mitteilungen Eidgenosischen Anstalt für Forstlichen Versuchswessen 27:406–468
Smith DW, Drummer TD, Murphy KM, Guernsey DS, Evans SB (2004) Winter prey selection and estimation of wolf kill rates in Yellowstone National Park, 1995–2000. J Wild Manag 68:153–166
Swetnam TW, Allen CD, Betancourt JL (1999) Applied historical ecology: using the past to manage for the future. Ecol Appl 9:1189–1206
van der Maarel E (2005) Vegetation ecology. Blackwell, Malden
Veselic Z (1991) Na Postojnskem prestevilna divjad se naprej hudo ogroza gozdno mladje. Gozdarski vestnik 49:147–157
Vrska T, Adam D, Hort L, Kolár T, Janík D (2009) European beech (Fagus sylvatica L.) and silver fir (Abies alba Mill.) rotation in the Carpathians—a developmental cycle or a linear trend induced by man? For Ecol Manag 258(4):347–356
Webb NF, Hebblewhite M, Merrill EH (2008) Statistical methods for identifying wolf kill sites using global positioning system locations. J Wild Manag 72:798–807
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by C. Ammer.
Rights and permissions
About this article
Cite this article
Klopcic, M., Jerina, K. & Boncina, A. Long-term changes of structure and tree species composition in Dinaric uneven-aged forests: are red deer an important factor?. Eur J Forest Res 129, 277–288 (2010). https://doi.org/10.1007/s10342-009-0325-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-009-0325-z