
Abstract
Within the framework of an interdisciplinary project on the effects of climate and forest management on beech-dominated deciduous forests (Swabian Jura, south-western Germany), forest meteorological investigations are carried out to analyse the influences of exposure and canopy density on the microclimate within various beech stands (Fagus sylvatica L.). This second paper of the series “Microclimate within beech stands” focuses on the thermal conditions that exist mainly in the near-surface layer at different test plots. They were analysed by air temperature, T a, (at 1.5 m a.g.l) and soil temperature, T s, at six depths measured continuously in the period 1999–2003. The main results can be summarized as follows. (1) The thermal situation within the beech stands described by T a depended primarily on the turbulent air-mass exchange conditions which were based on the slope-specific energy balance of the forest floor and advective heat fluxes. (2) The thermal situation within the soil described by T s at 3 and 20 cm depth was governed by the molecular heat transport. Therefore, the heating and cooling rates of T s were always lower than for T a. Higher T a and T s values for the test plots on the SW slope showed that the thermal conditions within the beech stands depended primarily on the exposure. (3) Based on slope-specific differences of daily extremes of T a and T s at 3 cm depth between the silviculturally treated and control plots, the influence of the more pronounced height growth of the understorey vegetation under the near-surface thermal conditions could be clearly verified for the NE slope.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The climatic conditions of a forest site are determined by its radiation, heat and water balance as well as by the balance of nutritional compounds. The main influence is governed by the synoptic situation and its long-term development. In complex terrain, the main quantities of the balance equations are additionally influenced by exposure and elevation, which leads to a specific climate at single sites. Silvicultural treatment, such as thinning, modifies the climatic conditions, and stand-specific microclimates arise below the canopy (e.g. Geiger 1961; Lee 1978; Aussenac 2000). Due to their importance for different forest processes and structures (e.g. stand regeneration, development of understorey vegetation, bio-geochemical processes), the microclimatic conditions below the canopy have been analysed already at various sites (e.g. Mayer 1979; Chen et al. 1993, 1999; Chen and Franklin 1997; Carlson and Groot 1997; Friedland et al. 2003), but these experimental investigations rarely exceeded 1 year. It is known that ecological processes in forests react on long-term microclimatic conditions, which, however, can be modified by extreme weather conditions. Hence investigations on the effects of microclimate on stand development and biodiversity need to last significantly longer than one year (Morecroft et al. 1998; Grimmond et al. 2000).
Beech (Fagus sylvatica L.) is the stand-forming tree species in many natural forest stands throughout central Europe, and forest administrations have demanded a stronger concentration on beech during the past few years (Hanewinkel 2001; Kenk and Guehne 2001). Nevertheless, there is a lack of systematic studies on the microclimate within beech stands, including the influence of regional climatic change as well as the consequences of forest management and topographic effects.
For these reasons, an interdisciplinary research project has been started in beech stands on the Swabian Jura, a mountain range in south-western Germany. The objective of this project is to quantify the influences of climate and forest management on beech-dominated deciduous forests (Rennenberg 2001). As part of the project, experiments are being carried out to investigate the dependence of the microclimatic conditions within the beech stands on the exposure and density of the canopy. The first paper of the series “Microclimate within beech stands” dealing with photosynthetically active radiation (PAR) (Mayer et al. 2002) showed a distinct dependence of PAR on the density of the canopy quantified by the plant area index (PAI). This paper on the thermal microclimate within different beech stands is based on air temperature, T a, and soil temperature, T s. The objective of this paper is to analyse the spatio-temporal variability of T a and T s within the beech stands on two opposite slopes which differ in exposure and in canopy density. As in the study on PAR, temporal variability of T a and T s is discussed by mean daily (February: month within the leafless period, August: month within the fully leaved period) and annual cycles for the year 2001. In addition, the thermal consequences of the development of the understorey vegetation caused by a strong shelterwood felling in 1999 are analysed at two test plots for the period 1999–2003.
Physical basics
The temporal changes of the potential temperature \( \frac{{\partial \Theta }} {{\partial {\text{t}}}} \) within an air volume, i.e. heating or cooling, can be described by thermodynamic laws as follows (Freytag 1988; Stull 1988; Kraus 2000):
where (I) is the three-dimensional advection of sensible heat (u, v and w are the components of the wind vector in the directions x, y and z of a three-dimensional Cartesian coordinate system), (II) is the three-dimensional turbulent exchange of sensible heat (u′, v′, w′ and Θ′ are the turbulent fluctuations of the three wind-vector components and the potential temperature, respectively) and (III) is the vertical divergence of the net all-wave radiation Q*. Near the surface, the potential temperature Θ can be approximated by the air temperature, T a (Stull 1988):
where g is the gravity acceleration, c p is the specific heat of the air at constant pressure (g/c p=0.0098 K/m), and z is the elevation above the 1,000 hPa-level. Therefore, the temporal changes of \( \frac{{\partial T_{a} }} {{\partial t}} \) are also determined by the processes of Eq. (1).
Assuming that soil heat flux Q s is the only heat source and sink within the soil, temporal changes of soil temperature \( \frac{{\partial T_{s} }} {{\partial t}} \) can be determined by the vertical gradient of Q s (e.g. Stull 1988; Foken 2003):
with c s being the volumetric heat capacity of the soil. Considering the approach for Q s
where a s is the molecular heat conductivity of the soil, Eq. (3) can be transformed into:
where ν T (=a s/c s) is the thermal diffusivity coefficient of the soil. Mean values of c s, a s and ν T for different soil types are compiled in Stull (1988) and Foken (2003), considering the dependence on soil moisture.
Therefore, temporal changes of T s are the result of molecular heat transport within the soil, whereas temporal changes of T a in complex terrain (van Gorsel et al. 2003) depend on turbulent air-mass exchange caused by the energy exchange at the surface and advective processes due to topographically induced thermal circulations (e.g. combination of catabatic and anabatic winds as well as mountain and valley winds).
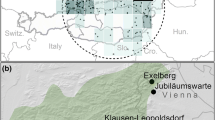
Site
The area under investigation is located near Moehringen (about 100 km east of Freiburg) and is described in detail by Rennenberg (2001), Geßler et al. (2001) and Mayer et al. (2002). In the Kraehenbach valley (about 690 m a.s.l., 8°45′E, 47°59′N), two main investigation sites were set up on the NE-exposed slope of the “Moehringer Berg” (up to 200 m above the bottom of the valley) and on the opposite SW-exposed slope of the “Muehlberg” (up to 180 m above the bottom of the valley). The slopes of the valley are covered with 70–80-year-old single-layer, beech-dominated (>90% Fagus sylvatica L.) stands. In the year 2001, the mean stand height, H, was about 28 m on the NE-exposed slope and about 26 m on the SW slope. Adjacent test plots of about 0.5 ha treated in silviculturally different ways (untreated control plots, weak shelterwood felling and strong shelterwood felling) were established at both main investigation sites. All plots consist of a 30×30 m core zone, where various measurements were carried out, and a peripheral zone of 20 m width, which was treated like the corresponding core zone.
At both investigation sites, the soil layer is shallow and can be characterised as terra fusca-rendzina, derived from limestone with high fractions of rocks and stones. On the NE-exposed slope, the soil contains 15% rocks and stones in the uppermost 20 cm of the soil and approximately 30% below 50 cm (Fotelli et al. 2003; Keitel et al. 2003). On the SW-exposed slope, the soil profile is particularly rocky, containing more than 40% (volumetric basis) rocks and stones in the top 20 cm of the soil and increasing up to 80% below 50 cm. The specific heat capacity of the dry soil determined experimentally in the year 2002 amounted to 0.83±0.044 J/(g K) on the NE slope and 0.93±0.034 J/(g K) on the SW slope. Gravimetrical analyses of the soil water potential during measurement campaigns showed higher levels of the soil moisture on the NE than the SW slope (Heidenfelder 2002). On a monthly basis, the highest differences of the soil moisture between test plots of the same silvicultural treatment on the NE and SW slope were lower than 10%. Differences of the soil moisture as mean values over 1999 to 2001 amounted to 2% between the control plots and 8% between the strong shelterwood plots on both slopes. In the investigation by Heidenfelder (2002), the local climate is classified as cool-humid on the NE slope and warm-dry on the SW slope.
Methodology
Measuring method
The transmission of climate data from official stations of the German Weather Service to the small-scale pattern of the specific test plots using geo-statistical methods (e.g. Xia et al. 1999, 2001; Hasenauer et al. 2003) was not possible in the required spatio-temporal resolution. Therefore, it was necessary to install specific stations on the test plots, where the measured meteorological data ensure the required resolution. Moreover, they enable the recording of more climate variables, which are fundamental for ecological research (e.g. PAR) but are not measured at official climate stations.
At each of the test plots on both slopes, air temperature, T a, and soil temperature, T s, are recorded continuously by an identical set of four forest-meteorological near-surface stations. They are located at two control plots (NE1.0 and SW2.0) and at two strong shelterwood plots (NE2.2 and SW1.2). Table 1 contains some features of these test plots. T a is measured at z=1.5 m a.g.l. by use of a Pt-100 sensor, which is mounted together with a Humicap-probe for the measurement of the relative humidity into a sensor-head, type HMP45D produced by Vaisala (Helsinki, Finland). For the comparative analysis of the thermal conditions above the canopy, T a values from two meteorological towers installed at both control plots were used. Measured T a values were taken at the relative height of z/H=1.30, because it was also the reference height for the investigation of the PAR conditions above the canopy. T s is measured by Pt-100 sensors at depths of 1, 3, 5, 10, 20 and 40 cm. At each test plot, all sensors for T a and T s are scanned every 30 s by a micrologger (type CR23X, Campbell Scientific Inc., Logan, UT, USA) and 30-min mean values are calculated automatically. More details about the micrometeorological near-surface stations and measurement towers, e.g. with respect to all measured variables and sensors, are given by Holst et al. (2000, 2001).
The plant area index (PAI), determined by measurements with two LAI-2000 Plant Canopy Analyzers (LI-COR Inc., Lincoln, NB, USA), is used to quantify the canopy density of each stand. The determination of PAI was carried out at specific days with homogeneous sky conditions (uniformly overcast) within the leafless period and in summer when the canopy is fully leaved (Holst et al. 2004). The difference in PAI between the leafless and fully leaved period can be interpreted as leaf area index (LAI). The PAI results presented in Table 2 are mean values based on up to nine single PAI measurements at 1.3 m a.g.l.
The results of the discontinuously determined PAI values from February 2000 to May 2003 show only a slight increase for the canopy of the test plots, which lies within the measuring accuracy. This may be interpreted as an indication that the canopy density at the test plots did not change noticeably during this period. Starting in July 2002, the additional reference height of 3 m a.g.l. was introduced for the determination of PAI at the strong shelterwood plots, because the understorey vegetation which developed significantly after the felling in 1999 had exceeded the first measurement height of 1.3 m a.g.l. The different development of the understorey vegetation at both strong shelterwood plots can be estimated from PAI differences between both reference heights. The understorey vegetation is dominated mainly by ash seedlings (Fraxinus excelsior L.).
Observation of the understorey development
Data from observations and measurements of the maximum height of the understorey vegetation layer are available for the description of the development of the understorey vegetation at the test plots. The height of the understorey vegetation was below 0.5 m at all test plots at the beginning of the research project in 1999. It has remained nearly constant at the control plots on both slopes up to now.
However, a pronounced growth of the understorey vegetation is typical of the strong shelterwood plots due to the pronounced availability of PAR (Mayer et al. 2002; Weisberg et al. 2003). For the first season after the silvicultural treatments, Keitel et al. (2003) and Fotelli et al. (2003) reported that at the strong shelterwood plots, the understorey vegetation had increased by 25% on the NE slope and by 8% on the SW slope, which was caused by different amounts of PAR. In June 2000, some shoots reached about 1 m at NE2.2 and about 0.75 m at SW1.2. In the year 2001, the understorey vegetation became denser at NE2.2 and reached a height of 1.6 m at the whole plot in the growing season, while it was distinctly lower at the SW1.2-plot with heights of about 1 m.
A height of 2.5 m was observed for the understorey vegetation at NE2.2 in summer 2002. Since the sensors of the near-surface stations were installed at 1.5 m a.g.l., the measurement of T a took place well within the understorey. While the understorey vegetation at NE2.2 could be characterised as a closed and very dense layer, it was composed of single shoots reaching heights of about 1.5 m at the SW1.2-plot. The different growth of the understorey vegetation at the NE2.2 and SW1.2-plots could be observed also in the year 2003. The understorey at NE2.2 was dense and reached a height of 2.8 m, while it was lower (about 2.5 m) at SW1.2. Compared to the year 2002, the understorey vegetation at SW1.2 was more dense and closed in the year 2003.
Results
Mean daily courses within the leafless and leaved period
Air temperature, T a
In the leafless season, T a differences between the test plots on the NE and SW slopes may result from their elevation differences, which is not more than 70 m (Table 1), and from their opposite aspect. The dominating effect of the exposure in the leafless period is shown by mean daily courses of T a within and above different beech stands in February 2001 (Fig. 1a, b). It caused higher mean maximum values of T a at the SW slope (SW2.0: 4.4 °C; SW1.2: 4.6 °C; SW (z/H=1.30): 3.5 °C) than at the NE slope (NE1.0: 2.8 °C; NE2.2: 2.7 °C; NE (z/H=1.30): 2.8 °C). Obviously, the difference of the mean T a maximum between z=1.5 m a.g.l. and z/H=1.30 was higher on the SW than the NE slope, which was caused by the increased irradiance on the SW slope and, therefore, the higher mean T a maximum near the energy-exchange surface. The T a differences above the canopy between NE and SW slope were lower than near the surface due to the more intensive turbulent air-mass exchange. A surface inversion developed during the night on both slopes, leading to higher values of T a above the stands than near the surface. Owing to a less dense canopy and, therefore, higher amounts of net long-wave radiation, the mean T a values in the night were lower at both strong shelterwood plots than at the control plots.

Hourly mean values of the air temperature, T a, within and above different beech stands (1.5 m a.g.l. and at z/H=1.30) on the NE slope (a) and on the SW slope (b) in February 2001
As indicated by the mean daily courses of T a in August 2001 (Fig. 2a, b), i.e. in the fully leaved period, the thermal conditions in the below-canopy space were dominated by the differences caused by the exposition of the sites. The mean maximum T a values were explicitly higher on the SW slope [SW2.0: 21.9 °C; SW1.2: 22.8 °C; SW (z/H=1.30): 21.6 °C] than on the NE slope [NE1.0: 20.2 °C; NE2.2: 20.2 °C; NE (z/H=1.30): 20.9 °C]. On both slopes, the dense canopy in the growing season caused pronounced differences of the extreme T a values between the strong shelterwood and the control plots, while they were not significant for mean T a values. According to the exposure of the slopes and the processes responsible for the heating of the air, the mean maximum of T a above the canopy on the SW slope was lower than the mean maximum T a values at SW2.0 and SW1.2. On the NE slope, however, NE1.0 and NE2.2 had lower mean maximum T a values than above the canopy.

Hourly mean values of the air temperature, T a, within and above different beech stands (1.5 m a.g.l. and at z/H=1.30) on the NE slope (a) and on the SW slope (b) in August 2001
The influence of the different exposure on the near-surface T a conditions can also be seen when analysing the extreme mean temporal changes of T a taken from Figs. 1 and 2. In February and August 2001, both mean maximal and minimal T a changes were lower for similar test plots on the NE than on the SW slope (Table 3). Due to the fully leaved canopy in the growing season (August 2001), the differences of the mean values of \( \frac{{\partial T_{a} }} {{\partial t}}_{{\max }} \;{\text{and}}\;\frac{{\partial T_{a} }} {{\partial t}}_{{\min }} \) between the strong shelterwood and the control plots were more pronounced on both slopes than in the leafless season (February 2001). Higher values of mean heating based on more intense solar irradiance could be found with decreasing canopy density (decreasing PAI values). In addition, net long-wave radiation in the nighttime increased with decreasing PAI values, which led to higher rates of cooling of the near-surface layer at the test plots. On both slopes, the mean values of \( \frac{{\partial T_{a} }} {{\partial t}}_{{\max }} \;{\text{and}}\;\frac{{\partial T_{a} }} {{\partial t}}_{{\min }} \) occurred almost at the same time for both months (February and August 2001): \( \frac{{\partial T_{a} }} {{\partial t}}_{{\max }} \) between 10 and 11 CET and \( \frac{{\partial T_{a} }} {{\partial t}}_{{\min }} \) between 18 and 19 CET.
Soil temperature, T s
The mean daily courses of T s at 20 cm depth showed almost no variability at all test plots in February 2001 (Fig. 3a, b), while they had the form of a distinct sinusoidal wave in August 2001 (Fig. 4a, b). At 3 cm depth, i.e. closer to the energy-exchange surface, the mean daily courses of T s could be characterised by a marked daily pattern, not only in the growing season (August 2001) but also in the leafless period (February 2001). In August 2001, the higher canopy density caused a more distinct daily T s course, with higher amplitudes at the strong shelterwood plots than at the control sites. Comparing both slopes, these differences between the test plots were more pronounced on the SW than on the NE slope.

Hourly mean values of the soil temperature, T s (depths: 3 and 20 cm) for different beech stands on the NE slope (a) and on the SW slope (b) in February 2001

Hourly mean values of the soil temperature, T s (depths: 3 and 20 cm) for different beech stands on the NE slope (a) and on the SW slope (b) in August 2001
In comparison to the values of Table 3, the mean extreme temporal changes of T s at 3 and 20 cm depth (Table 4) show that heating and cooling of the soil by molecular energy transport proceeded much more slowly than those of the air in the near-surface layer. In contrast to February 2001, in August 2001 the mean absolute values of \( \frac{{\partial T_{s} }} {{\partial t}}_{{\max }} \;{\text{and}}\;\frac{{\partial T_{s} }} {{\partial t}}_{{\min }} \) at 3 cm depth were significantly higher at the strong shelterwood than at the control plots on both slopes. This was mainly caused by the less dense structure of the canopy (lower PAI) at the strong shelterwood plots. The differences of the mean extreme heating and cooling of the soil were less pronounced at 20 cm depth. Compared to the mean extreme temporal changes of T a, mean extreme temporal changes of T s occurred after a 1–4 h-delay at the single depths, depending on the specific silvicultural treatment.
Mean T s tautochrones enable clear information to be gathered on the direction of the heat transport within the soil and the mean daily amplitude of T s at single depths. In February 2001 (Fig. 5a, b), heat was transported from the soil towards the soil surface at all plots. Vertical gradients of T s did not vary significantly between the plots. In February 2001, T s was up to 0.8 °C lower at NE2.2 than at NE1.0 at all observed times (Fig. 5a). For all depths at the test plots on the opposite slope (Fig. 5b), T s was slightly higher at SW1.2 than at SW2.0. In August 2001, the heat transport was directed from the soil surface into the soil (Fig. 6a, b). As in winter, vertical gradients of T s did not vary significantly between the plots in the deeper soil levels. Differences of the canopy density led to higher T s values at the strong shelterwood plots. Despite comparable PAI differences, T s differences between the strong shelterwood plot and the control plot were less pronounced on the NE slope (Fig. 6a) than on the SW slope (Fig. 6b).

Tautochrones of the soil temperature, T s, for different beech stands on the NE slope (a) and on the SW slope (b) in February 2001, where z is the depth

Tautochrones of the soil temperature, T s, for different beech stands on the NE slope (a) and on the SW slope (b) in August 2001, where z is the depth
Altogether, the influence of the exposure on T s could be demonstrated in several ways. (1) In both months discussed, the thermal level was higher on the SW slope than for comparable depths on the NE slope. Thus mean maximum T s values at 3 cm depth were higher on the SW slope (February 2001: SW2.0: 2.9 °C, SW1.2: 3.0 °C; August 2001: SW2.0: 16.8 °C, SW1.2: 18.7 °C) than on the NE slope (February 2001: NE1.0: 1.2 °C, NE2.2: 0.4 °C; August 2001: NE1.0: 16.1 °C, NE2.2: 17.0 °C). (2) In February 2001, hourly means of T s at the test plots on the NE slope were always higher at 20 cm than at 3 cm depth. On the SW slope, however, hourly means of T s at 3 cm depth were higher than at 20 cm depth within a short time in the evening. (3) In August 2001, differences of the mean maximum T s values in similar depths between the strong shelterwood and the control plots were caused by the different canopy densities. Due to the opposite exposure of both slopes, those differences were more pronounced on the SW than on the NE slope.
Mean annual courses
Daily mean values of T a
The strong shelterwood plots showed increased amplitudes of the mean daily courses of T a compared to similar test plots. Therefore, it could be assumed that daily mean values of T a differ only by tenths of degrees C between different plots. This assumption was confirmed by results from correlation analysis performed for daily mean values of T a in the year 2001. The comparatively high values for the coefficient of determination were mainly caused by the annual course of daily mean T a values, which was similar for all test plots. Compared to PAR (Mayer et al. 2002), the effects by exposure and canopy density were distinctly lower.
Due to the different exposure of the slopes, daily mean T a values were higher at SW2.0 than NE1.0 (Table 5 in consideration of the regression coefficient b). Regressions between mean daily T a values measured at the test plots SW2.0 and NE1.0 near Moehringen and those from the nearby Tuttlingen climate station of the German Weather Service (hereafter referred as to DWD) point out influences of different elevations (DWD Tuttlingen climate station: 648 m a.s.l.), exposure and canopy density. These regression analyses have been performed to investigate the alternative of a long-term data transfer from an official climate station.
The effect of different canopy densities on daily mean T a values could be identified only for the SW slope, where they were somewhat higher at SW1.2 than at SW2.0 (Table 6). A partitioning of daily mean T a values into daily mean T a values for the daytime and those for the nighttime proved to be more convenient to show the physical effects of different canopy densities on the thermal conditions in the near-surface layer. The regression coefficients a and b (Table 6) indicate that for higher thermal levels, i.e. in the growing season, the mean daytime T a values were higher at the strong shelterwood than at the control plots. The differences depended on the exposure and were more pronounced on the SW than NE slope. In contrast, the differences of the mean nighttime T a values between the warmer control plots and the cooler strong shelterwood plots reached higher amounts on the NE than on the SW slope. The main reason was the comparatively higher difference of stand density (Table 1) between NE1.0 and NE2.2 than between SW2.0 and SW1.2, which induced a higher difference of the net long-wave radiation between the strong shelterwood plot and the control plot on the NE slope.
Daily mean values of T s
Similar to the results for the daily mean T a values, daily mean T s values in the year 2001 differed slightly between the test plots. Caused by the same physical processes (e.g. annual pattern of solar irradiance), linear regression analyses of daily mean T s values between the single test plots showed a marked coefficient of determination (Table 7). The effects of exposure and canopy density were summarized into the regression coefficients a and b. Throughout the year 2001, the daily mean T s values in both depths (3 and 20 cm) were higher on the SW than on the NE slope. Considering both control plots as example, the differences between SW2.0 and NE1.0 decreased with increasing daily mean T s values at both depths.
On the NE and SW slope, different canopy densities caused higher mean daily T s values at the strong shelterwood than the control plot (Table 7), but this was limited for T s recorded at 20 cm depth on the NE slope to mean daily T s values of over 1 °C. In contrast to the influence of the exposure, the differences of the daily mean T s values between the strong shelterwood and control plots increased with higher daily mean T s values for both slopes. This was caused mainly by the pronounced PAI differences between both types of test plots during the growing season.
Monthly mean values of T a
The analysis of the monthly mean values of T a in the year 2001 is limited to the effects of different exposure. At the control plots, the mean T a values above the canopy (z/H=1.30) were up to 0.5 °C higher on the SW than on the NE slope from January to March and from September to December (Table 8). With respect to the strong shelterwood plots, T a reached higher monthly mean values throughout the year 2001 on the SW than on the NE slope. This difference was generally higher in the wintertime and peaked with 0.8 °C in March. In June and July, no differences of monthly mean T a values occurred between both control plots on the opposite slopes, which points out the influence of the closed canopy in the growing season. Except for June and July, monthly mean T a values at SW2.0 were up to 0.7 °C (January) higher than those at NE1.0. Comparing plots with different canopy densities on the NE slope, monthly mean T a values were up to 0.2 °C higher below the dense canopy (NE1.0) than at the thinned plot NE2.2, while on the SW slope, mean T a values from May to September were up to 0.2 °C higher below the less dense canopy (SW1.2) than at the control plot SW2.0.
Monthly mean values of T a, calculated separately for nighttime (T a,n) and daylight hours (T a,d), strengthen the effects of different exposure and canopy densities (see also Figs. 1 and 2) because the influences of solar irradiance as a source of energy during daylight hours and net long-wave radiation as a sink of energy during the night are much more pronounced than for monthly mean T a values calculated for the whole day. Due to the favourable exposure, monthly mean T a,d values were generally higher on the SW than on the NE slope (Table 8). The differences reached higher values for the strong shelterwood plots (up to 1.2 °C in October 2001) than for the control plots (up to 0.8 °C in January, February and October 2001). With respect to the SW slope, mean T a,d values were generally higher at SW1.2 than at SW2.0. The peak difference of 0.5 °C in July 2001 was caused mainly by the increased influence of the fully leaved canopy. On the NE slope, mean T a,d values at NE2.2 exceeded those at NE1.0 (up to 0.3 °C) only during the growing season (May to September), when the influence of the PAI differences revealed its maximum. In the leafless season (January to March and October to December), however, mean T a,d values were higher at NE1.0 (up to 0.2 °C).
The effect of different canopy densities (i.e. PAI values) was more pronounced for monthly mean T a,n values than for monthly mean T a,d values (Table 8). Due to an increased net long-wave radiation in the near-surface layer at the strong shelterwood plots, monthly mean T a,n values were higher at the control plots for almost the whole of 2001. On the NE slope, the peak difference of 0.8 °C between the control and strong shelterwood plot was obtained during the growing season from June to August, while the corresponding difference on the SW slope occurred with 0.7 °C in October. A systematic effect of the exposure on monthly mean T a,n values could not be verified, since the mean T a,n values at NE1.0 were up to 0.6 °C (August) higher than those at SW2.0 from May to August. At the strong shelterwood plots, a tendency of higher monthly mean T a,n values during winter (up to 0.5 °C in January and March) at SW1.2 than for NE2.2 could be found.
Comparing monthly mean T a,d and T a,n values (Table 8), peak differences were determined for all test plots in summer. In August 2001, they amounted to 4.4 °C at SW1.2, 3.8 °C at SW2.0, 3.5 °C at NE2.2 and 2.5 °C at NE1.0, i.e. they were higher (1) at the SW slope and (2) at the strong shelterwood plots on both slopes. In the leafless period, the differences between monthly mean T a,d and T a values reached higher amounts at all test plots than for the differences between monthly T a and T a,n values. On average over the leafless period 2001, these differences were slightly lower on the NE slope (NE2.2: 0.8 °C) than on the SW slope (SW1.2: 1.2 °C). In the growing season, however, the differences between the monthly mean T a and T a,n values were dominating. On average over the growing season 2001, they reached 3.5 °C at SW1.2, 2.8 °C at SW2.0, 3.0 °C at NE2.2 and 2.1 °C at NE1.0. The main reasons for these results were the imbalance between nighttime and daylight hours during summer and the increased net long-wave radiation at the strong shelterwood plots which led to lower night T a values than at the control plots.
Monthly mean values of T s
In the year 2001, the monthly mean values of T s at 3 and 20 cm depth were higher on the SW slope (Table 9) due to its favourable exposure. Analysing similar test plots on both slopes, mean peak T s differences occurred in February 2001 (2.2 °C between SW1.2 and NE2.2 at 3 and 20 cm depth). In accordance with these results, the differences of monthly mean T s values (at 3 and 20 cm depth) between similar test plots were up to 0.3 °C higher (between SW2.0 and NE1.0 at 3 cm depth and between SW1.2 and NE2.2 at 20 cm depth) in the leafless period (November to April) than in the growing season (June to September).
Differences of mean T s values between the strong shelterwood and control plots caused by the opposite exposure were (1) higher in the growing season (SW slope: 1.1 °C at both depths; NE slope: 0.8 °C at both depths) than in the leafless period (SW slope: 0.3 °C at 3 cm depth, 0.4 °C at 20 cm depth; NE slope: 0.2 °C at 3 cm depth, no difference at 20 cm depth), (2) slightly higher on the SW slope (maximum: 1.3 °C in August and September at both depths) than on the NE slope (maximum: 1.1 °C in May at 3 cm depth; 1.0 °C in June at 20 cm depth) and (3) reached its maximum on the NE slope earlier in the year than on the SW slope. Higher monthly mean T s values during the growing season at the strong shelterwood plots on both slopes could be interpreted as an influence of the less dense canopy than at the control plots, leading to an intensified solar irradiance in combination with prolonged daylight hours in summer compared to winter. As the exchange of heat within the soil is caused by molecular transport, the daily amplitude of T s at the strong shelterwood plots was less pronounced than for the near-surface T a.
Impact of the dynamic of the understorey vegetation
Besides canopy density and exposure, the vertical distance from the energy-exchange surface is important for the thermal conditions below the canopy. At the strong shelterwood plots, this surface was shifted away from the forest floor due to the fast development of the understorey vegetation. Slope-specific differences of daily extreme T a and T s values are suitable for the analysis of the impact on the thermal conditions within the beech stands on the opposite slopes resulting from the different increase of the understorey height.
Based on the differences of the daily extreme T a values between the strong shelterwood and control plot on the NE slope from September 1999 to July 2003 (Fig. 7a), the following results can be derived. (1) In spite of the annual course, a tendency of increasing differences of daily T a maxima between both types of test plots could be detected until the growing season 2001, which was caused by the decreasing distance between the active surface for the energy exchange and the measuring height of T a (1.5 m a.g.l.) at NE2.2. (2) During the fully leaved season 2002, the T a sensor was located within the regularly dense understorey, i.e. below the active surface. As a consequence, the differences of the daily T a maxima between NE2.2 and NE1.0 remained unchanged. (3) A decline of these differences was observed in the growing season 2003. (4) Due to the same reason as for (2), the amounts of the differences of the daily T a minima between NE2.2 and NE1.0 increased until the growing season 2001, remained unchanged during the fully leaved period 2002 and decreased slightly in the growing season 2003.

Differences of daily highest and lowest value of air temperature T a (1.5 m a.g.l.) between the test plots NE2.2 and NE1.0 (a) and between the test plots SW1.2 and SW2.0 (b) between 1 September 1999 and 31 July 2003
The results for the test plots on the SW slopes can be summarized as follows (Fig. 7b). (1) Due to the delayed growth of the understorey at SW1.2, the differences of the daily T a maxima between SW1.2 and SW2.0 increased until the growing season 2002 and showed a decreasing tendency during the growing season 2003, when the T a sensor was located within the understorey. (2) The differences of daily T a minima between SW1.2 and SW2.0 revealed neither an annual course nor a dependence on the height of the understorey until the fully leaved period 2002.
Since the understorey became higher and denser at NE2.2, the differences of the daily T s maxima at 3 cm depth between NE2.2 and NE1.0 decreased during the growing seasons (Fig. 8a). The differences of the daily T s minima between NE2.2 and NE1.0, however, did not show any trend. While their maximum amounts were lower than the comparable values of the daily T s maxima during the growing seasons 1999, 2000 and 2001, the amounts of the differences of the T s minima corresponded approximately to the differences of the daily T s maxima between NE2.2 and NE1.0 during the growing seasons 2002 and 2003. The distinct negative differences of the daily T s maxima in February/March were caused by snow at NE2.2, while at NE1.0 it melted some days before. Similar effects could not be observed on the SW slope, because the more intensive solar irradiance led to higher soil temperatures and, therefore, a faster melting of snow.

Differences in daily highest and lowest value of soil temperature T s (3 cm depth) between the test plots NE2.2 and NE1.0 (a) and between the test plots SW1.2 and SW2.0 (b) between 1 September 1999 and 31 July 2003
The results for the SW slope (Fig. 8b) show negative differences of the daily T a maxima between SW1.2 and SW2.0 at the beginning of spring 2000, which were a consequence of different effects of the solar irradiance at test plots with various canopy densities. Since the elevation angle of the sun is already high in March and April, solar irradiance at SW2.0 was large enough to warm the soil despite the more dense, but still leafless, canopy. At the same time, nighttime cooling at SW1.2 was higher than at SW2.0 due to the lower canopy density. With the unfolding of the leafs of the beech canopy at the beginning of May, solar irradiance at SW2.0 was reduced significantly leading to a positive difference T s,max (SW1.2)−T s,max (SW2.0). Similar conditions occurred in spring 2002 and 2003 in a less pronounced way. Kirchgäßner (2001) observed the positive influence of a higher canopy density in terms of increased near-surface air and soil temperatures on the phenological evolution of the herbaceous vegetation on both test plots. As a consequence, the development of the herbaceous vegetation at SW2.0 was about 1 week ahead compared to SW1.2. As on the NE slope, the SW slope showed also the tendency that the differences of the daily T s maxima between SW1.2 and SW2.0 decreased in the growing seasons.
Discussion
In contrast to similar investigations at other forest sites (e.g. Geiger 1961; Mayer 1979; Chen et al. 1993, 1999; Chen and Franklin 1997; Morecroft et al. 1998; Grimmond et al. 2000; Friedland et al. 2003), this experimental study is designed for long-term forest-meteorological measurements at different test plots in complex terrain. Therefore, it was possible to investigate not only the impacts of varying canopy densities, but also the consequences of two opposite exposures and the effects of the understorey dynamics under differing silvicultural treatments.
The results for different cycles of T a and T s (mean daily courses, mean annual courses based on daily means, daily extremes, monthly means and monthly means for the daylight and nighttime hours, respectively) attained in this study can be characterised by a generally higher thermal level at the test plots on the SW slope. Hence it follows that the thermal microclimate within the beech stands depends strongly on the exposure throughout the year. This was expected regarding the physical processes for heating and cooling the near-surface air in complex terrain. The additional influence of the different canopy densities during the fully leaved period could be proved only for results of T a and T s, which are separately related to periods with dominating solar irradiance (daylight hours) or net long-wave radiation (nighttime). Since the thermal impacts of different canopy densities in both periods are similar, daily and monthly means of T a and T s showed almost no influence of the canopy density on the thermal near-surface microclimate. However, it could be clearly detected when mean daily courses per month, daily extremes and monthly means for the daylight and nighttime hours of both thermal variables are analysed (see also Mayer 1979; Aussenac 2000; Grimmond et al. 2000; Potter et al. 2001). The contributions of the canopy density on T a and T s were always lower than differences of T a and T s between similar types of test plots caused by the orientation of the slopes.
Compared to the magnitude of order of PAR differences due to different canopy densities, T a and T s differences between the single test plots were distinctly lower, even if their dependence on the timescale is considered. Based on the physical laws of the energy exchange within the near-surface atmosphere in complex terrain, the differences of T a and T s at the test plots were (1), with respect to the influence of the canopy density, comparable to similar results from other investigations (e.g. Mayer 1979; Carlson and Groot 1997; Morecroft et al. 1998; Potter et al. 2001; Prévost and Pothier 2003) and (2), with respect to the significance of the exposure, typical of the topoclimate in mountainous regions in central Europe covered with forests (e.g. Freytag 1988).
One reason for forest-meteorological studies lies in the great importance of the microclimate for the biological environment, i.e. soil-biological and plant-physiological processes and conditions. In general, differences of the microclimatic conditions support biodiversity in forests (e.g. de Freitas and Enright 1995). Structural, anatomic, morphological and physiological properties of trees depend on the respective microclimatic conditions (e.g. Aussenac 2000); these properties are adjusted to the microclimate, at least partially. The thermal microclimate controls biochemical conversions and, therefore, the forest’s supply of nutrition. Air temperature acts as a catalytic converter, which controls the rates of processes such as respiration, transport of water and nutrition, soil formation and soil transformation. Different thermal conditions influence the development of seedlings and shoots (e.g. Frey et al. 2003). An experimental investigation on beech trees (Hurst 1988) showed that lower soil temperatures restricted the uptake of water as well as the growth of the roots and led to less foliage mass as well as decreased radial growth. Grimmond et al. (2000) pointed out that the climatic conditions are decisive for litter decomposition and soil respiration. They emphasised their significance for ecosystem research and the carbon cycles of forests.
Contemporaneous biological investigations at the test plots near Moehringen indicated the significance of the microclimatic results in a complex context. The additive effect of varying growth conditions at the single plots, which include the different thermal conditions, was analysed by Kirchgäßner (2001) applying phenological observations on the test plots during the years 1999 and 2000. With respect to the soil, Heidenfelder (2002) ascertained that the nitrogen transformation rates and the content of nitrogen in the microbial biomass were higher on the warmer SW slope than on the cooler NE slope. Evaluating the consequences of the silvicultural treatments on the microbial processes in the soil, which are involved in the nitrogen transformation rates, the combination of the variables of soil temperature and soil moisture, which are related to each other, turned out to be an important complex of factors. Fotelli et al. (2003) found a statistically significant dependence between δ13C in the wood of beech seedlings and soil temperature. Keitel et al. (2003) obtained a strong link between stomata conductance and air temperature, which resulted in a relationship between air temperature and transpiration of beech stands. Analysing the dynamics of radial stem variance and radial growth of beech trees at the test plots, Hauser (2003) observed that air temperature and soil temperature were relevant for the explanation of the varying daily radial stem variances.
Concluding remarks
Summarising the results on the thermal microclimate and PAR (Mayer et al. 2002) within different beech stands, the working hypothesis of the interdisciplinary research project on beech-dominated deciduous forests near Moehringen (Rennenberg 2001) can be preliminarily reviewed. (1) A specific contribution was made to eliminate the lack of systematic investigations on the microclimate within beech stands. (2) The general estimation of the SW slope as the warmer one could be confirmed by the near-surface air temperature and soil temperature. (3) Differences of the microclimate conditions in beech stands between control plots and silviculturally treated plots caused by differing canopy densities could be verified dependent on exposure. (4) The simulation of expected future climatic conditions was possible to some extent by the use of test plots on two opposite slopes with differing aspect. On the spatial scale of topoclimate, the differences of the near-surface air temperature T a due to this experimental design were in the lower range of retrospective changes of annual means of T a at climate stations of the German Weather Service on the Swabian Jura during the past 50 years (Kirchgäßner 2001). However, new results of regional climate simulations by suited models show a higher increase in T a for the coming decades, which can be simulated more precisely by use of experimental sites in different regional climates.
Abbreviations
- PAR:
-
Photosynthetically active radiation
- PAI:
-
Plant area index
- LAI:
-
Leaf area index
References
Aussenac G (2000) Interactions between forest stands and microclimate: ecophysiological aspects and consequences for silviculture. Ann For Sci 57:287–301
Carlson DW, Groot A (1997) Microclimate of clear-cut, forest interior, and small openings in trembling aspen forest. Agric For Meteorol 87:313–329
Chen J, Franklin JF (1997) Growing season microclimate variability within an old-growth Douglas-fir forest. Clim Res 8:21–34
Chen J, Franklin JF, Spies TA (1993) Contrasting microclimates among clearcut, edge, and interior of old-growth Douglas-fir forest. Agric For Meteorol 63:219–237
Chen J, Saunders SC, Crow TR, Naiman RJ, Brosofske KD, Mroz GD, Brookshire BL, Franklin JF (1999) Microclimate in forest ecosystem and landscape ecology. Bioscience 49:288–297
Foken T (2003) Angewandte Meteorologie. Springer, Berlin Heidelberg New York
Fotelli MN, Rennenberg H, Holst T, Mayer H, Geßler A (2003) Carbon isotope composition of various tissues of beech (Fagus sylvatica) regeneration is indicative of recent environmental conditions within the forest understorey. New Phytol 159:229–244
de Freitas CR, Enright NJ (1995) Microclimatic differences between and within canopy gaps in a temperate rainforest. Int J Biometeorol 38:188–193
Frey BR, Lieffers VJ, Munson AD, Blenis PV (2003) The influence of partial harvesting and forest floor disturbance on nutrient availability and understorey vegetation in boreal mixed forests. Can J For Res 33:1180–1188
Freytag C (1988) Atmosphärische Grenzschicht in einem Gebirgstal bei Berg- und Talwind. Wiss Mitt Meteor Inst Univ München No 60
Friedland AJ, Boyce RL, Vostral CB, Herrick GT (2003) Winter and early spring microclimate within a mid-elevation conifer forest canopy. Agric For Meteorol 115:195–200
Geiger R (1961) Das Klima der bodennahen Luftschicht. Vieweg-Verlag, Braunschweig
Geßler A, Schrempp S, Matzarakis A, Mayer H, Rennenberg H, Adams MA (2001) Radiation modifies the effect of water availability on the carbon isotope composition of beech (Fagus sylvatica). New Phytol 150:653–664
Grimmond CSB, Robeson SM, Schoof JT (2000) Spatial variability of micro-climate conditions within a mid-latitude deciduous forest. Clim Res 15:137–149
Hanewinkel M (2001) Economic aspects of the transformation from even-aged pure stands of Norway spruce to uneven-aged mixed stands of Norway spruce and beech. For Ecol Manage 151:181–193
Hasenauer H, Merganicova K, Petritsch R, Pietsch SA, Thornton PE (2003) Validating daily climate interpolations over complex terrain in Austria. Agric For Meteorol 119:87–107
Hauser S (2003) Dynamik hochaufgelöster radialer Schaftveränderungen und des Dickenwachstums bei Buchen (Fagus sylvatica L.) der Schwäbischen Alb unter dem Einfluss von Witterung und Bewirtschaftung. PhD Thesis, Fak Forst-Umweltwiss Univ Freiburg
Heidenfelder A (2002) Einfluß von Klima und Bewirtschaftung auf am N-Umsatz beteiligte mikrobielle Prozesse in Waldböden. PhD Thesis, Forstwiss Fak Univ Freiburg
Holst T, Rost J, Schindler D, Matzarakis A, Mayer H (2000) Mikroklimatische Untersuchungen in südwestdeutschen Buchenbeständen. Rep Meteor Inst Univ Freiburg No. 5, 123–135
Holst T, Matzarakis A, Mayer H, Rost J, Schindler D (2001) Mikroklima in Buchenbeständen auf gegenüberliegenden Hängen in der Schwäbischen Alb. In: Beitr zur 1. Deutsch-Österreichisch-Schweizerischen Meteorologentagung Wien, September 2001. Österreich Beitr Meteor Geophys Heft 27, Session 7a, pp 1–18
Holst T, Hauser S, Kirchgäßner A, Matzarakis A, Mayer H, Schindler D (2004) Measuring and modelling plant area index in beech stands. Int J Biometeorol (in press)
Hurst A (1988) Wirkung von Bodenabkühlungen auf Wasserbilanz und Wachstum von Buchen (Fagus sylvatica L.). Mitt Eidg Anst Forstl Versuchswes, vol 64
Keitel C, Adams MA, Holst T, Matzarakis A, Mayer H, Rennenberg H, Geßler A (2003) Carbon and oxygen isotope composition of organic compounds in the phloem sap provides a short-term measure for stomatal conductance of European beech (Fagus sylvatica L.). Plant Cell Environ 26:1157–1168
Kenk G, Guehne S (2001) Management of transformation in central Europe. For Ecol Manage 151:107–119
Kirchgäßner A (2001) Phänoklimatologie von Buchenwäldern im Südwesten der Schwäbischen Alb. Rep Meteor Inst Univ Freiburg No. 7
Kraus H (2000) Die Atmosphäre der Erde—Eine Einführung in die Meteorologie. Vieweg-Verlag, Braunschweig
Lee R (1978) Forest microclimatology. Columbia University Press, New York
Mayer H (1979) Mikroklimatische Untersuchungen im ostbayerischen Bergmischwald. Arch Met Geoph Biokl Ser. B, 27:247–262
Mayer H, Holst T, Schindler D (2002) Mikroklima in Buchenbeständen—Teil I: Photosynthetisch aktive Strahlung. Forstwiss Centralbl 121:301–321
Morecroft MD, Taylor ME, Oliver HR (1998) Air and soil microclimates of deciduous woodland compared to an open site. Agric For Meteorol 90:141–156
Potter BE, Teclaw RM, Zasada JC (2001) The impact of forest structure on near-ground temperatures during two years of contrasting temperature extremes. Agric For Meteorol 106:331–336
Prévost M, Pothier D (2003) Partial cuts in a trembling aspen-conifer stand: effects on microenvironmental conditions and regeneration dynamics. Can J For Res 33:1–15
Rennenberg H (2001) Buchendominierte Laubwälder unter dem Einfluss von Klima und Bewirtschaftung: Ökologische, waldbauliche und sozialwissenschaftliche Analysen. Abschlussbericht des Sonderforschungsbereiches 433. Univ Freiburg
Stull RB (1988) An introduction to boundary layer meteorology. Kluwer, Dordrecht
van Gorsel E, Christen A, Feigenwinter C, Parlow E, Vogt R (2003) Daytime turbulence statistics above a steep forested slope. Boundary-Layer Meteorol 109:311–329
Weisberg PJ, Hadorn C, Bugmann H (2003) Predicting understorey vegetation cover from overstorey attributes in two temperate mountain forests. Forstwiss Centralbl 122:273–286
Xia Y, Winterhalter M, Fabian P (1999) A model to interpolate monthly mean climatological data at Bavarian forest climate stations. Theor Appl Climatol 64:27–38
Xia Y, Fabian P, Winterhalter M, Zhao M (2001) Forest climatology: estimation and use of daily climatological data for Bavaria, Germany. Agric For Meteorol 106:87–103
Acknowledgements
The authors are indebted to the German Research Foundation (DFG) for grants under SFB 433 (subproject A1), MA-749/17-1 and MA-749/17-2, the national forest office at Tuttlingen and the city administration of Tuttlingen for permission to use the forest sites, as well as to Prof. Dr. Heinz Rennenberg, Institute of Tree Physiology and Forest Botany of the University of Freiburg, for his scientific initiative to interdisciplinary investigations on beech stands near Moehringen.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Holst, T., Mayer, H. & Schindler, D. Microclimate within beech stands—part II: thermal conditions. Eur J Forest Res 123, 13–28 (2004). https://doi.org/10.1007/s10342-004-0019-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-004-0019-5