
Abstract
In order to improve the efficiency of the known floral attractant of Epicometis hirta [(E)-anethol and (E)-cinnamyl alcohol in a ratio of 1:1], candidate synergist compounds for field tests were selected through electroantennographic tests using the antennae of female and male E. hirta adult beetles. Among synthetic floral compounds 4-methoxyphenethyl alcohol and methyl salicylate evoked high responses from the antennae and were chosen for further field studies. In trapping tests in Bulgaria, the addition of 4-methoxyphenethyl alcohol to the E. hirta bait in the ratio of 1:1:1, in Hungary in ratios of 1:1:0.3 and 1:1:1 significantly increased catches. The addition of methyl salicylate was without effect in both countries. A high-capacity trap supplied with this ternary attractant could be more efficient for mass trapping purposes of E. hirta in environment-friendly plant protection.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Adults of Epicometis hirta Poda (Coleoptera: Scarabaeidae, Cetoniinae) damage the reproductive parts of the flowers of several ornamental plants and orchard trees all over Europe (Hurpin 1962). The adult beetles cause damage to flowers of apple, cherry, and many other spring-blossoming fruit trees. Also, they frequently damage cereals, primarily ears of rye. The beetles can feed on many flowering weeds, i.e. different spp. of Compositae and Cruciferae. They chew the petals, staminae and stigmae, thus rendering the flower infertile. They not only damage flowers in full blossom, but also in the bud stage (Hurpin 1962). In Central Europe the overwintering adults emerge from the soil in April; the peak of the swarming period is in May. The grub (larva) lives in the soil, feeds on rotting plant material, it causes no damage (Endrödi 1956).
Control of this pest is difficult, since most insecticides cannot be applied during flowering without affecting honeybees or other beneficials. Mass trapping of adult beetles by high-capacity traps baited with highly effective attractants could be an excellent alternative control method, as it could reduce damage to below an economically acceptable threshold and, at the same time, would be non-toxic, posing no health or environmental risk to other organisms, and so could be used throughout the blossoming period.
Epicometis hirta, similar to many other day-active flower-feeding insects, uses both chemical and visual cues in locating host plants. It has a strong preference towards blue colour (Schmera et al. 2004). A binary chemical attractant consisting of (E)-cinnamyl alcohol and (E)-anethol has been optimised previously which proved to be attractive towards adult beetles of E. hirta (Tóth et al. 2003, 2004). These compounds are common flower scent constituents (Knudsen et al. 1993). When applying both the chemical attractant and the blue colour, catches of E. hirta were significantly higher than in traps containing only one of the cues (Schmera et al. 2004). Such traps are in use today in Central and Eastern Europe for monitoring and mass trapping of E. hirta (Mircheva et al. 2004; Sivcev et al. 2006; Tóth et al. 2005).
The objective of the present research was to improve the efficiency of the binary E. hirta bait in order to make it more effective for mass trapping of this scarab pest. To keep the number of candidate compounds to be tested within practical limits, we screened synthetic floral compounds in electroantennogram (EAG) assays on the antennae of female and male E. hirta. It was thought that high EAG responses showed that the antenna was sensitive to the given compound, and chances to find field-active co-attractants were higher among compounds eliciting high EAG activity. Of course, if a compound elicits high EAG responses, this does not necessarily mean that it will have also behavioural activity, nor does it indicate what type of behavioural activity (attractancy, repellency, etc.) is to be expected (Roelofs 1977). Compounds selected on the basis of the EAG response spectra were field tested to see whether their addition improved the activity of the known binary E. hirta bait.
Materials and methods
Chemicals
Altogether 27 synthetic compounds (mostly floral) described earlier to be attractive to Scarabaeidae and to other species in related insect taxa or characterized to be components of fermenting natural insect bait liquids were chosen for electrophysiological tests (i.e. Donaldson et al. 1986, 1990; Landolt et al. 2000; Landolt 2000). Synthetic compounds were obtained from Sigma-Aldrich Kft. (Budapest, Hungary). All compounds were >95% pure as stated by the supplier.
Electroantennograms (EAGs)
For recording EAGs, a stainless steel tube (teflon-coated inside) with a constant humidified airflow of ca. 0.7 l/min was set up. An antenna freshly amputated at the base from a live beetle was mounted between two glass capillaries containing 0.1 M KCl solution, and the mounted antenna was placed at ca. 3 mm distance from the outgoing airflow. One of the electrodes was grounded, while the other was connected to a high impedance DC amplifier (IDAC-232, Syntech, Hilversum, The Netherlands). Test compounds (10 μg each) were administered in hexane solution to a 10 × 10-mm piece of filter paper inside a Pasteur pipette. This dosage was thought to exceed the saturation level for all compounds and testing compounds at such a high dose would balance for differences in volatility of the compounds as suggested for similar EAG screening studies by Roelofs (1977).
Stimuli consisted of pushing 1 ml of air through the Pasteur pipette into the airstream flowing towards the antenna. Stimuli were administered in random order. Response amplitudes were normalized against the mean of responses to 1-phenylethanol (eliciting medium high responses from antennae), which was tested before and after the test compounds. Stimuli were administered at ca. 20- to 30-s intervals. EAG-measurements were conducted in the lab at 21 °C.
Experimental insects were collected from the edge of an oak forest at Julianna major (Pest county, Hungary) in the middle of April 2006 by random sampling of E. hirta specimens feeding on flowers. Insects collected were kept in cages (40 × 30 × 30 cm) under natural conditions and fed apple pieces. One cage contained ca. 20 specimens altogether. Sexes were separated based on the presence or absence of penis.
For EAG-analysis, data were subjected to ANOVA and the means were separated by Fisher’s protected LSD. In comparison of responses evoked by highly EAG-active compounds from female or male antennae Student t test was used. Statistical procedures were conducted using the software packages StatView® v4.01 and SuperANOVA® v1.11 (Abacus Concepts Inc., Berkeley, CA).
Baits
For preparing the bait dispensers, a 1-cm piece of dental roll (Celluron®, Paul Hartmann AG, Heidenheim, Germany) was placed into a tight polythene bag made of 0.02-mm linear polyethylene foil. The dimensions of the polyethylene sachets were ca. 1.5 × 1.5 cm. The dispenser was attached to a plastic strip (8 × 1 cm) for easy handling when assembling the traps. For making up the baits, each compound was administered onto the dental roll and the opening of the polythene bag was heat-sealed. Earlier experience showed that the bait did not loose its activity during several weeks of field exposure; hence, we decided to replace the lures at 2- to 3-week intervals.
Dispensers were wrapped singly in pieces of alufoil and were stored at −30°C until use.
Traps
In the tests, CSALOMON® VARb3 modified funnel traps (produced by Plant Prot. Inst., HAS, Budapest, Hungary) were used, which proved to be excellent for the capture of related scarabs (Imrei et al. 2001; Schmera et al. 2004). Photos of VARb3 traps can be viewed at http://www.julia-nki.hu/traps.
Field tests
Experiments were conducted at several sites in Bulgaria and in Hungary in 2007 and 2008. Traps were set up in a randomized complete block design (1 block consisted of 1 trap of each treatment). The distance between traps was 10–15 m. Traps were set up in sunny places, attached to poles at a 30–40 cm height above ground. Traps were inspected twice weekly, when captured insects were recorded and removed.
The objective of experiments 1 and 2 was to study the effect of the addition of 4-methoxyphenethyl alcohol or methyl salicylate to the known binary attractant.
In case of experiments 3 and 4, the objective was to study a dose/response of 4-methoxyphenethyl alcohol when added to the binary bait. Details of the experiments and the specific treatments are shown in Tables 1, 2.
The data units for the field experiments were number of insects caught/trap/inspection. Capture data were analysed by Kruskal–Wallis followed by pairwise comparisons by Mann–Whitney U test, using the software package StatView® v4.01 (Abacus Concepts Inc., Berkeley, CA).
Results
Electroantennographic measurements
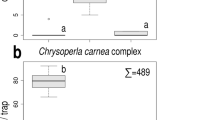
When screening synthetic floral compounds on the antennae of E. hirta, methyl salicylate, 4-methoxyphenethyl alcohol, (E)-cinnamyl acetate, eugenol, geraniol, phenethyl acetate and phenylacetaldehyde elicited highest EAG responses from the antennae of both sexes (Figs. 1, 2). (E)-Cinnamyl alcohol and (E)-anethol (the two components of the known bait) evoked medium high responses in males and females. Responses of female or male antennae showed similar trends with possibly the exceptions of (E)-cinnamyl acetate, which evoked significantly higher responses from male antennae (P = 0.0133), and geraniol, which also elicited higher responses from male antennae (P = 0.0435).

EAG responses of antennae of E. hirta males to synthetic compounds. Responses were normalized to response evoked by the standard 1-phenylethanol. Columns show means of responses from 12 males. Mean columns not significantly different at P = 5% (ANOVA, Fisher’s Protected LSD) are below the same group line (drawn above the columns) labelled with different letters

EAG responses of antennae of E. hirta females to synthetic compounds. Responses were normalized to response evoked by the standard 1-phenylethanol. Columns show means of responses from 18 females. Mean columns not significantly different at P = 5% (ANOVA, Fisher’s Protected LSD) are below the same group line (drawn above the columns) labelled with different letters
Field experiments
Experiments 1, 2
Significantly more E. hirta were recorded at both sites in traps baited with 4-methoxyphenethyl alcohol and the known binary attractant together than in traps baited with the binary attractant alone (Fig. 3). The addition of methyl salicylate to the binary attractant was without effect at either site.

Mean catches of E. hirta in traps baited with the binary and ternary mixtures of (E)-anethol, (E)-cinnamyl alcohol, 4-methoxyphenethyl alcohol and methyl salicylate (total catch in experiment 1, 222 beetles; in experiment 2, 73 beetles). P values from Mann–Whitney U test (paired comparisons) following Kruskal–Wallis test
Experiments 3, 4
In ratio tests at both sites, baits containing 4-methoxyphenethyl alcohol showed a tendency of catching more than the known binary attractant (Fig. 4). In Hungary, traps baited with the mixtures of (E)-anethol:(E)-cinnamyl alcohol:4-methoxyphenethyl alcohol in 1:1:0.3 or 1:1:1 ratios caught significantly more E. hirta than the binary mixture alone, while catches with the 1:1:0.1 ratio did not increase significantly. In Bulgaria, traps baited with the ternary mixture in 1:1:1 ratio caught significantly more E. hirta than the binary attractant on its own (Fig. 4). Catches in traps baited with the ternary blends did not differ from each other neither in Bulgaria nor in Hungary.

Mean catches of E. hirta in traps baited with the binary lure of (E)-anethol + (E)-cinnamyl alcohol and the binary lure plus 4-methoxyphenethyl alcohol in three different ratios (total catch in experiment 3, 243 beetles; in experiment 4, 3,250 beetles). P values from Mann–Whitney U test (paired comparisons) following Kruskal–Wallis test
Discussion
Among compounds eliciting highest responses from antennae of both sexes methyl salicylate and 4-methoxyphenethyl alcohol were chosen for further field tests since (E)-cinnamyl acetate, eugenol, geraniol, phenethyl acetate and phenylacetaldehyde in earlier field tests showed only low or no activity in attracting E. hirta (Tóth et al. 2003; unpublished data).
The present results clearly show the positive effect of the addition of 4-methoxyphenethyl alcohol to the E. hirta known bait [consisting of (E)-cinnamyl alcohol and (E)-anethol]. It appears from the ratio tests (Fig. 4) that the most effective dose of 4-methoxyphenethyl alcohol should range between 1:1:0.3 and 1:1:1. Further field tests need to be carried out in order to determine the exact optimal dose of 4-methoxyphenethyl alcohol.
In the above experiments, 4-methoxyphenethyl alcohol was not tested on its own. However, the fact that in other tests (aimed at other target insects and conducted at sites where E. hirta was also present) we never observed catches of this scarab in traps baited with 4-methoxyphenethyl alcohol (Tóth et al. unpubl.), suggests that the compound may not have a strong attractive effect on E. hirta when presented alone. The improved ternary attractant described in the present study may improve efficiency of mass trapping efforts aimed to control E. hirta. Studies to support this notion are underway and will be published in the future.
From the two EAG-active compounds which were tested in field experiments, methyl salicylate is a very widespread floral scent constituent, and has been found in many plant families (Knudsen et al. 1993; Knudsen and Tollsten 1993, 1995; Jakobsen et al. 1994; Flamini et al. 2002; Grison-Pigé et al. 2002). Methyl salicylate has been described as an attractant for several beetles (Wolde-Havariat et al. 2007; Maekawa et al. 1999; Rutledge and Young 2007; Hammack 2001; Kamm and Buttery 1984; Dickens 2006).
The field-active compound in this study, 4-methoxyphenethyl alcohol is present in the plant families of Hyacinthaceae (Knudsen et al. 1993), Araceae (Kite and Hetterscheid 1997), Orchidaceae (Lawrence 1993) and Annonaceae (Jürgens et al. 2000). 4-Methoxyphenethyl alcohol is exploited as an attractant by Diabrotica barberi Smith and Lawrence (Coleoptera: Chrysomelidae) (Metcalf 1998).
References
Dickens JC (2006) Plant volatiles moderate response to aggregation pheromone in Colorado potato beetle. J Appl Entomol 130:26–31
Donaldson JMI, McGovern TP, Ladd TL Jr (1986) Trapping techniques and attractants for Cetoniinae and Rutelinae (Coleoptera: Scarabaeidae). J Econ Entomol 79:374–377
Donaldson JMI, McGovern TP, Ladd TL Jr (1990) Floral attractants for Cetoniinae and rutelinae (Coleoptera: Scarabaeidae). J Econ Entomol 83:1298–1305
Endrödi S (1956) Lamellicornia. Fauna Hungariae IX/4. Akadémiai, Budapest
Flamini G, Cioni PL, Morelli I (2002) Analysis of the essential oil of the aerial parts of Viola etrusca from Monte labbro (South Tuscany, Italy) and in vivo analysis of flower volatiles using SPME. Flavour Fragr J 17:147–149
Grison-Pigé L, Hossaert-McKey M, Greeff JM, Bessière J-M (2002) Fig volatile compounds—a first comparative study. Phytochem 61:61–71
Hammack L (2001) Single and blended maize volatiles as attractants for diabroticite corn rootworm beetles. J Chem Ecol 27:1373–1390
Hurpin B (1962) Super-Famille des Scarabaeoidea. In: Balachowsky AS (ed) Entomologie Appliquée a l’Agriculture. Masson et Cie, Paris, pp 24–204
Imrei Z, Tóth M, Tolasch T, Francke W (2001) 1,4-Benzoquinone attracts males of Rhizotrogus vernus Germ Z Naturforsch 57c 177–181
Jakobsen HB, Friis P, Nielsen JK, Olsen CA (1994) Emission of volatiles from flowers and leaves of Brassica napus in situ. Phytochemistry 37:695–699
Jürgens A, Webber AC, Gottsberger G (2000) Floral scent compounds of Amazonian Annonaceae species pollinated by small beetles and thrips. Phytochem 55:551–558
Kamm JA, Buttery RG (1984) Root volatile components of red clover: identification and bioassay with the clover root borer (Coleoptera: Scolytidae). Environ Entomol 13:1427–1430
Kite GC, Hetterscheid WLA (1997) Inflorescence odours of Amorphophallus and Pseudodracontium (Araceae). Phytochemistry 46:71–75
Knudsen JT, Tollsten L (1993) Trends in floral scent chemistry in pollination syndromes: floral scent composition in moth-pollinated taxa. Bot J Linn Soc 113:263–284
Knudsen JT, Tollsten L (1995) Floral scent in bat-pollinated plants: a case of convergent evolution. Bot J Linn Soc 119:45–57
Knudsen JT, Tollsten L, Bergström LG (1993) Floral scents—a checklist of volatile compounds isolated by head-space techniques. Phytochem 33:253–280
Landolt PJ (2000) New chemical attractants for trapping Lacanobia subjuncta, Mamestra configurata, and Xestia c-nigrum (Lepidoptera, Noctuidae). J Econ Entomol 93:101–106
Landolt PJ, Smithhisler CS, Reed HC, Mcdonough LM (2000) Trapping social wasps (Hymenoptera, Vespidae) with acetic-acid and saturated short-chain alcohols. J Econ Entomol 93:1613–1618
Lawrence BD (1993) The scents of orchids. Elsevier, Amsterdam
Maekawa M, Imai T, Tsuchiya S, Fujimori T, Leal WS (1999) Behavioral and electrophysiological responses of the soybean beetle, Anomala rufocuprea Motchulsky (Coleoptera: Scarabaeidae) to methyl anthranilate and its related compounds. Appl Entomol Zool 34:99–103
Metcalf RL (1998) Ultramicrochemistry of insect semiochemicals. Mikrochim Acta 129:167–180
Mircheva A, Subchev M, Sredkov I, Tóth M (2004) Seasonal flight of Epicometis hirta Poda (Coleoptera, Scarabaeidae) established by attractant traps. Annuaire de l’Université de Sofia “St. Kliment Ohridski” 96:201–204
Roelofs WL (1977) The scope and limitations of the electroantennogram technique in identifying pheromone components. In: McFarlane NR (ed) Crop protection agents—their biological evaluation. Academic Press, New York, pp 147–165
Rutledge CE, Young DK (2007) First report of an attractant for a tumbling flower beetle (Coleoptera: Mordellidae). Environ Entomol 36:894–989
Schmera D, Tóth M, Subchev M, Sredkov I, Szarukán I, Jermy T, Szentesi Á (2004) Importance of visual and chemical cues in the development of an attractant trap for Epicometis (Tropinota) hirta Poda (Coleoptera: Scarabaeidae). Crop Prot 23:939–944
Sivcev I, Tóth M, Tomasev I (2006) Application of attractants in control of the sugar-beet weevil and the blossom feeder scarab (Bothynoderes punctiventris, Epicometis hirta). Poljoprivedni Kalendar 2006:138–140 (in Serbian)
Tóth M, Klein MG, Imrei Z (2003) Field screening for attractants of scarab pests in Hungary (Coleoptera: Scarabaeidae). Acta Phytopathol Entomol Hung 38:323–331
Tóth M, Schmera D, Imrei Z (2004) Optimization of a chemical attractant for Epicometis (Tropinota) hirta Poda. Z Naturforsch 59c:288–292
Tóth M, Imrei Z, Szarukán I, Voigt E, Schmera D, Vuts J, Harmincz K, Subchev M (2005) Chemical communication of fruit- and flower-damaging scarabs: results of one decade’s research efforts. Növényvédelem 41:581–588 (in Hungarian)
Wolde-Havariat Y, Seyoum E, Jembere B, Negash M, Hansson BS, Hillbur Y (2007) Behavioural and electrophysiological response of sorghum chafer Pachnoda interrupta (Coleoptera: Scarabaeidae) to plant volatiles. Int J Trop Sci 27:53–61
Acknowledgments
The present research was partially supported by the grant of KUKBOGMV OM-00063/2008JÁP and by the Grant DOO2-244/2008 of the Bulgarian National Scientific Fund. The GC-EAD apparatus was financed by GVOP 3.2.1-2004-04-0153/3.0 (major part), by OTKA T37355 (minor part) and by Dr. Jan van der Pers (SYNTECH) to Dr. G. Szöcs.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by M. Traugott.
Rights and permissions
About this article
Cite this article
Vuts, J., Szarukán, I., Subchev, M. et al. Improving the floral attractant to lure Epicometis hirta Poda (Coleoptera: Scarabaeidae, Cetoniinae). J Pest Sci 83, 15–20 (2010). https://doi.org/10.1007/s10340-009-0263-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10340-009-0263-z