
Abstract
How actions are chosen, and what they are influenced by, has been the focus of several research traditions. Influences on actions are often studied using compatibility paradigms, such as response priming. Here, a first stimulus (i.e., the prime) is presented shortly before a second stimulus (i.e., the target) which has to be classified. Reaction times to the target are often reduced when primes and targets are compatible compared to incompatible primes and targets—i.e., a positive compatibility effect (PCE). There are, however, some conditions in which reliably negative compatibility effects (NCEs), with faster reactions to incompatible targets, are found. Actions in real life are often influenced by perceived motion and are less determined by following (target) stimuli as it is the case in typical response priming studies with predetermined stimulus–response mappings. Thus, in the current experiment we used motion primes in forced-choice trials (with >> and << as targets) as well as in free-choice trials (with <> and >< as targets). Essentially, we found PCEs in the short-SOA condition and NCEs in the long-SOA condition. The pattern was not qualified by task (i.e., forced choice/free choice). The results provide evidence that NCEs with motion primes are found even without strong links between target stimuli and responses and that especially PCEs can be found with simpler and smaller targets than have been used in previous experiments.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
How actions are chosen, and what they are influenced by, has been the focus of several research traditions, from basic research on cognition (see below) to applied sciences, such as research on purchasing choices and consumer behavior. Influences on actions are often studied using compatibility paradigms. Participants are required to respond to a target stimulus or a target feature while ignoring preceding or simultaneously presented irrelevant stimuli or stimulus features (i.e., primes or distracters). Targets and primes/distracters either call for the same response (i.e., they are compatible, congruent, or consistent), or call for different responses (i.e., they are incompatible, incongruent, or inconsistent). One successful paradigm for studying influences (or pre-activations) of previous stimuli on responses to a current target is the response priming paradigm.
Response priming
In response priming, a first stimulus (i.e., the prime) is presented shortly before a second stimulus (i.e., the target) which has to be classified. Reaction times (RTs) to the target are often reduced when primes and targets are compatible compared to incompatible primes and targets—i.e., a positive compatibility effect (PCE; for a review see Schmidt et al. 2011a; for a review on mechanisms of response priming see Kiesel et al. 2007). However, there are some conditions in which reliably negative compatibility effects (NCEs), with faster reactions to incompatible targets, are found, for example with masked primes and longer stimulus onset asynchronies (typically, SOAs above 200 ms) between prime and target (e.g., Eimer and Schlaghecken 2002, 2003). Schlaghecken et al. (2006) argued that NCEs (with masked primes) reflect an inhibition mechanism in low-level motor control; however, there is a continuing debate as to whether this pattern of effects is a result of (such) motor, central (e.g., attentional), or perceptual (e.g., object updating) mechanisms and how many processes (e.g., self-inhibition, mask-triggered inhibition, object updating, counting processes within different “evaluation windows” accumulating evidence for one stimulus/category/response, attentional processes) contribute to this effect (e.g., Cole and Kuhn 2010; Klauer and Dittrich 2010; Lleras and Enns 2004; Verleger et al. 2004; for reviews see Kiesel et al. 2007, 2008). Additionally, NCEs can be the consequence of bottom-up or top-down influences (e.g., Schmidt et al. 2015). The most common current view seems to be that several processes contribute to NCEs, perhaps to different extents in different situations, tasks, conditions, and so on (see, for example, Krüger et al. 2013).
An example of evidence for the multiple-processes view comes from a nice recent experiment by Liu and Wang (2014). In their experiment, the authors used digital numbers as primes, masks, and targets. The primes/targets were the digital numbers 1 and 3, or 2 and 4. The mask was always the digital number 8. The key here was that the numbers were presented in such a way that combining 1 and 3 resulted in 8. Thus, when 1 was masked by 8, the new (i.e., added) information was 3, and vice versa. In these cases, during the prime–mask sequence, the perceptual information was updated by another piece of meaningful information (i.e., another number), and further, the new information in these cases was associated with another response. In contrast, when 2 or 4 was masked by 8, the added information did not result in another number; thus, this update was not associated with any response. In utilizing these facts, the authors showed that a NCE occurred earlier when the mask added exactly those elements to the prime which call for the non-primed response compared to trials in which the new information was not associated with any response. This is evidence that object updating—or the process by which the mental representation of individual objects is updated over time under the influence of new information (Lleras and Enns 2004)—contributes to the NCE. Further, depending on training (and thus the strength of S–R links of stimuli and requested response), the NCE primarily originates more from object updating, a perceptual-level process, or more from automatic motor inhibition, a motor-level process.
Free-choice priming
Response priming—as introduced above—has mainly been investigated using shape (e.g., arrows or diamonds vs. circles) or color stimuli (e.g., red vs. green dots). Often, simple arrows (< or >) or double arrows (<< or >>) are used as primes and targets (e.g., Eimer and Schlaghecken 2002). In the standard case of response priming, as in most other compatibility paradigms, participants are requested to respond with one reaction (e.g., “press X key”) to stimulus 1 and with another reaction (e.g., “press Y key”) to stimulus 2 (i.e., forced choice). Shortly after the novel result of NCEs in response priming with masked static primes, there evolved a variant which combined the priming procedure with a free-choice task. (Free-choice tasks were originally introduced by Berlyne 1957.) That is, participants could freely choose which of two buttons they want to press in response to a target. For these free-choice trials, the targets to which the response has to be made are often ambiguous, or at least in no manner related to one of the two buttons, responses, or directions. For example, a circle (Schlaghecken et al. 2009), the number zero (Kiesel et al. 2006), a diamond (Klapp and Hinkley 2002), a hash tag (Ocampo 2015), or bidirectional arrows (most often <>, e.g., Bodner and Mulji 2010; O’Connor and Neill 2011; Schlaghecken and Eimer 2004; or block arrows pointing in each direction, e.g., Parkinson and Haggard 2014) are used as (go) stimuli for free-choice responses.
The consistent finding with free-choice priming is that it in most cases closely resembles the pattern found with forced choices. That is, when a PCE is found with forced choices there are also faster and/or more prime-congruent button presses with free choices, and vice versa with a NCE and incongruent responses. Free-choice priming effects are found especially with high proportions of compatible trials in forced-choice trials (Bodner and Mulji 2010) and are larger with directional than arbitrary stimuli (e.g., Mattler and Palmer 2012), which matches findings of forced-choice priming effects that are larger when S–R links are more established. Free-choice priming effects are even found with go/no-go tasks (e.g., Parkinson and Haggard 2014; Schlaghecken et al. 2009), primes which are not used as targets (Ocampo 2015), spatial location primes (Mattler and Palmer 2012), and also if the response mapping is contrary to the usual mapping (i.e., left/right responses are mapped to right/left pointing arrows; O’Connor and Neill 2011). However, free-choice priming depends on context or task set: First, it can only be found in experimental blocks in which free-choice trials are interspersed into forced-choice trials (Klapp and Haas 2005; Schlaghecken and Eimer 2004); second, it can be found if the local context, i.e., the preceding trial, is compatible even in a global context with a low number of compatible forced-choice trials (Bodner and Mulji 2010). Further, there are some reports of positive correlations between forced-choice and free-choice priming effects. Even for response priming research, this is interpreted as evidence that the two forms have a common origin (Bodner and Mulji 2010; Klapp and Hinkley 2002; Ocampo 2015).
Taken together, the results of free-choice priming studies with masked static primes show that our actions are influenced by (subliminal) primes even when we can choose our responses freely. The influences can be either facilitative (PCE) or detrimental (NCE). Influences on forced choices as well as on free choices, however, depend on context, with effects being larger when the S–R strength between prime and response is high.
Motion
The question as to “what influences our actions” is closely connected with a core element of human information processing, that is, the ability to adjust ongoing activity to the changing environment (e.g., Burle et al. 2005). Changing environments often contain various kinds of motions. Some of them are more important than others. The crux is to quickly decide between important and less important information, and to quickly adapt own actions to the changing information and/or motion where applicable. The ability to perceive and discriminate different types of motions is vastly important for this and is even present in newborns (e.g., Farroni et al. 2004; Simion et al. 2008). Highlighting its importance, the special role of motion and moving things in perception and attention has been the focus of several research traditions (e.g., Bosbach et al. 2005; Egeth and Yantis 1997; O’Craven et al. 1997; Treue and Maunsell 1996).
Although motion is highly important in guiding own actions and influencing our behavior in everyday life, until now it has been seldom used in response priming experiments (for exceptions with somewhat uncommon versions of response priming see Mattler and Fendrich 2007, and Sarkheil et al. 2008). To bridge this gap, Bermeitinger (2013) introduced a motion variant of response priming in which moving dot rows (henceforth “row-of-dots primes”) were used as primes for static arrow targets. In further experiments, we tested other motion types (e.g., single dots as primes; Bermeitinger and Wentura 2016). Similar to the above findings with masked primes, the sign of the compatibility effects crucially depended on the SOA between prime and target when utilizing motion primes. Essentially, with short SOAs, positive compatibility effects emerged; with longer SOAs, we found negative compatibility effects. The exact point of time at which the reversal from a PCE to a NCE occurred depended on the kind of motion.
The negative compatibility effect using moving primes (Bermeitinger 2013; Bermeitinger and Wentura 2016) is especially noteworthy because clearly visible primes (motion stimuli) were used without any further constraints (i.e., there was no intervening stimulus; no further stream of information had to be processed, e.g., Klauer and Dittrich 2010; primes did not have to be presented at a randomly varying non-target location, e.g., Wyatt and Machado 2013). The negative compatibility effect with motion primes was interpreted as being evidence for some kind of counter-regulation in response to unwarranted or overshot activations, perhaps at the motor level. The conclusion of the previous research with motion primes is that consciously perceived motion seems to influence our actions in a special way, most probably by quickly activating corresponding motor responses with the need to quickly (counter-)regulate them. Some explanations for negative compatibility effects with static (masked) primes can be excluded for the NCEs with motion primes, especially those which assume that a mask or intervening stimulus triggers an inhibitory process as there is no such event in the experiments with motion primes. However, the exact processes are still under debate. Further, it is unclear in which aspects priming with motion primes and priming with (masked) static primes are comparable.
Current research
In spite of the consistency of the findings reported above—separate for response priming with motion primes and free-choice priming with static masked primes—there are still unanswered questions, especially when combining both lines of research. First, in our previous experiments on response priming with motion primes (Bermeitinger 2013; Hacklaender et al. 2015; Bermeitinger and Wentura 2016; Eckert and Bermeitinger 2016), we used rather large target arrows. The arrows used for row-of-dots primes were approximately 3.34° visual angle (3.5 cm) in length and 0.96° visual angle (1.0 cm) in height. For single-dot primes we already had to change the arrows (they were approximately 2.63° visual angle, i.e., 2.75 cm, in length and 0.96° visual angle, i.e., 1.0 cm, in height) as the original arrows would have masked the visibility of the motion of a single moving dot (Bermeitinger and Wentura 2016). Thus, it also could have been that the original negative compatibility effects were influenced by the fact that the arrows were able to reduce the visibility of the primes by masking them (at least in part)—they were presented after the motion primes, in part at the same position, and with high intensity. This could have resulted in backward masking of the primes and in (more or less) reducing the further conscious access to them (e.g., Enns and Di Lollo 2000). Then, we would have a similar situation as in the response priming studies with intentionally masked primes. Besides the possible problem of reducing prime visibility, the large targets can be categorized by solely attending the tip of the arrow. If the primes are able to shift attention (and/or eye gaze) toward the motions’ goal, the distinguishing feature in compatible trials is closer to the focus of attention. This might explain the faster responses in compatible trials in short SOAs. Thus, the first aim of the current experiment is to use the same targets as used in previous studies (e.g., by Eimer and Schlaghecken 2002)—that is, simple double arrows at the center of the screen for which the problems of strong benefits due to attentional shifts near to the arrows tip and potential issues with the targets masking the primes should be reduced (although not completely eliminated).
Second, it is unclear whether the pattern found with motion primes necessarily needs a clear pre-specified response for the targets. Thus, the question arises: Are (re)actions only influenced by motion primes if there are instructed and defined motor responses for each target and strong S–R links between a given target stimulus and its assigned response? Or, in contrast, do we find the same pattern even without such clear affordances but when subjects could freely choose their responses to ambiguous targets (i.e., <> or ><)? Besides the generally interesting question (= how perceived motion influences free-choice reactions), this line of research has potential theoretical implications: Without an instructed response and with ambiguous targets, there is no conflict between prime (response) and target (response) and accounts which explain compatibility effects by such a conflict (cf., Botvinick et al. 2001; Schmidt et al. 2011b—for their rapid chase theory) might not be applicable.
In order to investigate these two lines of questions and to replicate and extend the pattern found with motion primes, we simply mixed forced-choice and free-choice trials and presented in a randomly chosen manner forced-choice and free-choice targets after motion primes. We used simple double arrows as targets, and SOAs for which in previous experiments a positive compatibility effect (i.e., short SOA = 147 ms) and a negative compatibility effect (i.e., long SOA = 360 ms) were found. Demonstrations of the prime motions also used in the current experiment can be found as movie clips in the supplementary online material to Bermeitinger and Wentura (2016).
Methods
Participants
The sample consisted of 49 students from the University of Hildesheim. Participants had normal or corrected-to-normal vision. They were paid for their participation or participated in exchange for partial course credit. Informed consent was obtained from all individual participants included in the study. One participant from the 147-ms SOA condition had to be excluded due to equipment failure (The refresh rate was automatically set to 60 Hz instead of maintaining 75 Hz.) Four further participants (2 from each SOA condition) were excluded as they were outliers (more than 1.5 interquartile ranges above the third quartile) regarding the mean error rate in the forced-choice trials (Their error rates were between 18.75 and 43.75%.) Of the remaining participants, 21 (19 females; median age of 20 years, ranging from 18 to 31 years) were assigned to the 147-ms SOA condition and 23 (19 females; median age of 21 years, ranging from 19 to 26 years) to the 360-ms SOA condition.
Positive and negative compatibility effects in Bermeitinger (2013) were quite substantial: The positive effect at SOA = 150/147 ms varied from d = 0.59 to 1.06 (M = 0.83) across experiments (compatibility effects in mean RT varied from 11 to 17 ms); the negative effect at SOA = 350/360 ms varied from d = 0.61 to 1.05 (M = 0.78) across experiments (compatibility effects in mean RT varied from − 10 to − 17 ms). With respect to the mean effect sizes, it can be stated that the compatibility effect in our 147-ms SOA condition—if we concede its existence—can be detected with probability 1−β = 0.80 (α = 0.05, two-tailed) if one uses a sample size of at least N = 14. The corresponding effect in the 360-ms SOA condition can be detected with 1−β = 0.80 (α = 0.05, two-tailed) if one uses a sample of at least N = 15, respectively. (Power calculations were computed with G*Power; Faul et al. 2007.)
Design
A 3 (motion direction: rightward, leftward, neutral) × 2 (response: left, right) × 2 (task: forced choice vs. free choice) × 2 (SOA: 147, 360 ms) design was used. The factors motion direction, response, and task were varied within participant (please note that in the case of free-choice task, the response was not varied but chosen by participants); the factor SOA was varied between participants. The main focus lies on the compatibility/congruency of motion direction and response, which were either compatible/congruent (dots moved rightward and the right key was requested in the case of forced-choice trials or pressed in the case of free-choice trials; dots moved leftward and the left key was requested/pressed), incompatible/incongruent (dots moved rightward and left key was requested/pressed; dots moved leftward and right key was requested/pressed), or neutral (dots moved inward or outward). In the tradition of priming experiments, for forced-choice trials, the compatibility effect was computed as the difference (in target RTs or error rates) between incompatible and compatible trials; for free-choice trials, the congruency effect was computed as the difference (in target RTs) between prime-incongruent and prime-congruent responses. Additionally, in free-choice trials we analyzed the percentage of prime-congruent/prime-incongruent choices.
Materials
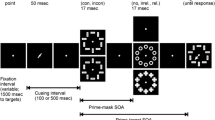
For the forced-choice task, two non-ambiguous double arrows, composed of two less-than or greater-than symbols, respectively, were used as target stimuli—one pointing to the left (<<) and one to the right (>>). For the free-choice task, two ambiguous targets were used, consisting of one greater-than and one less-than symbol, respectively (i.e., >< and <>). At a viewing distance of about 60 cm the targets were approximately 0.67° visual angle (0.7 cm) in length and 0.38° visual angle (0.4 cm) in height. For the primes, the same material was used as in Bermeitinger (2013). Essentially, for the prime event, there was a row of 11 black dots; each dot was approximately 0.38° visual angle (0.4 cm) in diameter. The whole row measured approximately 13.78° visual angle (14.5 cm) in length; the distance from one dot to the next dot was approximately 0.96° visual angle (1.0 cm). The prime event started with the presentation of the row at the center of the screen. To instantiate the movement, the dots were shifted from their original position in steps of 0.16° visual angle (0.17 cm) leftward or rightward. After six steps, a dot had reached the original position of its neighboring dot and the movement started again from the screen’s center (original row position). For the compatible and incompatible conditions, the dots (i.e., the whole row) were moved rightward or leftward. For the neutral conditions, the dots were either moved outward (i.e., the 5.5 left dots of the row moved leftward and the 5.5 right dots of the row moved rightward, meaning that the central dot was split into two semicircles that drifted apart) or inward (i.e., the 5.5 left dots of the row moved rightward, and the 5.5 left dots of the row moved leftward, meaning that the central dot was split into two semicircles that progressively superimpose; for more details see Bermeitinger 2013). Arrows and dots were presented in black on a white background.
Procedure
Participants were individually tested in sound-attenuated chambers. The experiment was run using E-Prime software (version 2) with standard PCs and 17’’ CRT monitors with a refresh rate of 75 Hz. Stimulus presentation was synchronized with the vertical retrace signal of the monitor. Instructions were given on the CRT screen. Participants were requested to quickly and accurately categorize each non-ambiguous arrow with regard to its direction (by pressing the right/left key with their right/left index finger for right/left arrows, respectively; response keys were the “3” and “1” key on the numeric pad on which a right or left arrow was pasted, respectively). Participants were further requested to quickly press the left or right key in response to ambiguous targets according to free choice. They were instructed to distribute the left and right key responses approximately evenly.
The sequence of each trial was as follows: First, a fixation stimulus (+) appeared at the center of the screen for 1000 ms. It was followed by the first row of dots, which was presented for one refresh cycle (i.e., approximately 13 ms) at the center of the screen. Then, the next row was presented for the next refresh cycle and so on. Prime presentation duration was 147 ms (i.e., 11 dot presentations * refresh cycle). For the 147-ms SOA condition, the prime event was followed immediately by the target, which remained on the screen until a response was given. For the 360-ms SOA conditions, the prime event was followed by a blank screen which remained for 213 ms; this was immediately followed by the target, which again remained on the screen until a response was given. The intertrial interval was 400 ms.
Each participant worked through four blocks with 72 trials each. Each block consisted of 36 forced-choice trials: 12 compatible trials (6 with dots moving rightward and leftward, respectively), 12 incompatible trials (6 with dots moving rightward and leftward, respectively), and 12 neutral trials (6 with dots moving inward, 6 with dots moving outward); half of the trials had right arrow targets and the other half had left arrow targets. The other 36 trials of each block were free-choice trials: 12 trials with rightward moving dots, 12 trials with leftward moving dots, 6 trials with inward moving dots, and 6 trials with outward moving dots; half of the trials had >< targets and half of the trials had <> targets. The order of trials within each block was chosen randomly by the computer. There was a short pause after each block. Before the first experimental block, there was a practice phase with 24 trials (half forced choice). Each experiment took about 15 min.
Results
Mean RTs in the forced-choice trials were derived from correct responses only. Mean error rates were 3.94% (SD = 2.58) and 2.96% (SD = 3.23) for the 147- and 360-ms SOA conditions, respectively. Outlying RTs that were 1.5 interquartile ranges above the third quartile with respect to the individual distribution (Tukey 1977), were above 1500 ms, or were below 200 ms were discarded for the forced-choice as well as the free-choice trials. Based on the outlier criterion, 1.59% (SD = 1.19) of all forced-choice and 4.63% (SD = 1.94) of all free-choice trials from the 147-ms SOA condition, and 2.17% (SD = 1.67) of all forced-choice and 5.40% (SD = 2.82) of all free-choice trials from the 360-ms SOA condition were discarded. Mean RTs and mean error rates of the forced-choice trials and mean RTs and mean rates of the prime-congruent and prime-incongruent responses in the free-choice trials are shown in Table 1, separately for each SOA condition.
RTs
Mean RTs from the compatible/prime-congruent, incompatible/prime-incongruent, and neutralFootnote 1 conditions were subjected to an ANOVA for repeated measures with the within-participant factors compatibility/congruency and task (forced choice; free choice) and the between-participants factor SOA (147, 360 ms). Most importantly, there was a significant interaction of compatibility/congruency and SOA, F(2, 84) = 7.68, p = 0.001, \(\eta_{p}^{2}\) = 0.15, which was not further qualified by task (p = 0.368) (Fig. 1). Follow-up analyses with a focus on compatible/prime-congruent and incompatible/prime-incongruent trials confirmed the significant interaction of compatibility/congruency and SOA for the forced-choice trials, F(1, 42) = 10.33, p = 0.003, \(\eta_{p}^{2}\) = 0.20, as well as for the free-choice trials, F(1, 42) = 6.35, p = 0.016, \(\eta_{p}^{2}\) = 0.13. In order to more directly measure the compatibility/congruency effects, we calculated the corresponding effects as explained in Design section, i.e., the difference scores (RT incompatible/incongruent—RT compatible/congruent) for each subject, and submitted these to one-sample t tests against the test value of 0. These t tests indicated that subjects performed slower in incompatible/prime-incongruent than compatible/prime-congruent trials in the 147-ms SOA condition (Mboth_tasks = 8 ms, SEM = 3.16, t(20) = 2.41, p = 0.026, d = 0.53; Mforced_choice = 13 ms, SEM = 3.92, t(20) = 3.22, p = 0.004, d = 0.70; Mfree_choice = 3 ms, SEM = 4.36, t(20) = 0.60, p = 0.555, d = 0.13) and slower in compatible/prime-congruent than incompatible/prime-incongruent trials in the 360-ms SOA condition (Mboth_tasks = − 8 ms, SEM = 2.37, t(22) = 3.30, p = 0.003, d = 0.49; Mforced_choice = − 5 ms, SEM = 3.98, t(22) = 1.35, p = 0.189, d = 0.28; Mfree_choice = − 10 ms, SEM = 2.82, t(22) = 3.63, p = 0.001, d = 0.76).

Reaction times of the compatible, incompatible and neutral conditions in forced-choice trials and of the prime-congruent responses, prime-incongruent responses and neutral trials of the free-choice trials (error bars represent the standard error of the mean)
In addition to the results above, the ANOVA also revealed a significant main effect of task, F(1, 42) = 60.30, p < 0.001, \(\eta_{p}^{2}\) = 0.59, showing faster responses in forced-choice (M = 408, SEM = 6.58) than free-choice trials (M = 455, SEM = 10.75), and an interaction of task and SOA, F(1, 42) = 4.12, p = 0.049, \(\eta_{p}^{2}\) = 0.09, which is of no further interest. All other effects including within-subject variables as well as the main effect of SOA were not significant, ps > 0.12 and p = 0.871, respectively.
There was no significant correlation between compatibility effects and congruency effects, neither in the 147-ms SOA condition (r = 0.17, p = 0.476) nor in the 360-ms SOA condition (r = − 0.06, p = 0.785).
Error/response rates
Mean error rates from the compatible, incompatible, and neutral conditions of the forced-choice trials were subjected to an ANOVA for repeated measures with the within-participant factors compatibility and the between-participants factor SOA. Error rates differed significantly between compatibility conditions, F(2, 84) = 4.24, p = 0.018, \(\eta_{p}^{2}\) = 0.09, with more errors in incompatible than compatible trials (contrast: F(1, 42) = 4.57, p = 0.038) and more errors in neutral than compatible trials (contrast: F(1, 42) = 10.15, p = 0.003), but no difference between errors in incompatible and neutral trials (F < 1, p = 0.678). This main effect was not further qualified by SOA, F < 1, p = 0.636, and there was no main effect of SOA, F = 1.21, p = 0.278.
For the free-choice trials, the mean percent of prime-congruent responses was tested against the rate of random choice (i.e., 50%) using a series of one-sample t tests. These t tests revealed that rate of prime-congruent responses did not differ from the random rate in either SOA condition (Mboth_SOAs = 49.43%, SEM = 1.69, M150 = 48.71%, SEM = 2.54, M360 = 50.09%, SEM = 2.28), ts < 1.02, ps > 0.32. A further independent-samples t test revealed that the prime-congruent response rate did not differ between the two SOA conditions, t = 0.81, p = 0.422.
Discussion
The general question for the current research was as follows: What influences our actions? In particular, we were interested in the question of how motions influence our actions and with the question of how forced-choice and free-choice responses are (differentially) influenced. For investigating these questions, we used a response priming paradigm with motion primes, as motions have a special impact in triggering and influencing our daily behavior. Our experiment, thus, was unique in two ways: First, we used motion primes, and second, the motion primes were clearly visible. In contrast to the material most often used for static primes—i.e., shapes, colors, words which are symbols that had to be transformed to the task-specific requirements—the perception of motion might directly trigger movements without the detour via some kind of transformation.
We found further evidence for compatibility and congruency effects with motion primes, in this instance by utilizing simple double arrows in forced-choice as well as free-choice trials. Essentially, as in our previous research (Bermeitinger 2013; Bermeitinger and Wentura 2016; Hackländer et al. 2015), in RTs we observed a switch from a positive to a negative compatibility/congruency effect from a short (147-ms) to a long (360-ms) SOA. The pattern was present in forced-choice as well as in free-choice trials; however, when looking at simple effects, the PCE was especially present in forced-choice trials and the NCE was especially present in free-choice trials. As there was no interaction with task, we did not further interpret these (merely) numerical differences between forced- and free-choice trials.
In our previous experiments with motion primes, we used rather large arrows as targets. These could have reduced the visibility of the primes by covering a larger area and acting as a mask of the primes. This, in turn, might have influenced the compatibility effects. We now used the same targets as used in many other response priming studies (e.g., Eimer and Schlaghecken 2002), namely simple double arrows consisting of greater-than and less-than symbols at the center of the screen. The utilization of these simple targets ensures that the primes were, spatially, hardly covered and their visibility should not be impaired. With these targets, we found the same general pattern of positive effects with short and negative effects with long SOAs as with larger targets (Bermeitinger 2013; Bermeitinger and Wentura 2016; Hackländer et al. 2015). We see this as evidence that the concrete target and its size do not cause the observed PCE–NCE pattern, although it might be that the targets moderate the effect.
In relation to target properties there may be, especially in the short-SOA condition, a strong benefit, first if the prime causes an attentional shift and, second, if the decisive target property is located near the locus of this shift (Cole and Kuhn 2010). This constellation might be given in compatible trials with large arrow targets which would make it specifically easy to categorize the target (for which the target’s tip might be sufficient), leading to faster responses in compatible than incompatible trials. With the simple and much smaller double arrows, an attentional shift—if involved—caused by the primes should be of lesser influence, first because it takes longer to return to the arrows tip and, second, because the spatial (and attentional) difference between the arrows tips in compatible and incompatible trials is smaller. However, in the current study we found a PCE of M = 13 ms (d = 0.70) that is highly comparable to previously found effects with our motion primes with large arrow targets (PCEs at SOA = 150/147 ms varied from M = 11 to 17 ms, d = 0.59–1.06 across experiments; Bermeitinger 2013). Taken together, it seems rather unlikely that attentional shifts as outlined above caused the compatibility effects found here and in previous research with motion primes. Instead, facilitation due to motor pre-activations caused by the prime seems to be a more appropriate candidate for explaining the observed effects (see, for example, Kiesel et al. 2006 for a more detailed review on several possible motor explanations of (subliminal) response priming).
We should note that, in the current experiment, with longer SOAs the NCE on forced-choice trials (in RTs) was not on its own significant, even though we found a negative difference between incompatible and compatible trials which did not significantly differ from the negative congruency effect in free-choice trials. Furthermore, on the forced-choice trials we found a PCE in terms of error rates. We here want to discuss several possible reasons for this.
First of all, the compatibility effect (in RTs at the longer SOA) clearly tended toward the expected NCE. Second, it is not atypical to fail to replicate the significance of each single case, even though we did replicate the pattern of results (in terms of RTs). Third, certain aspects of the current experiment differed from those of the normal setting. Most importantly, we added free-choice trials which (1) prolonged the experiment, (2) have changed some timing parameters, (3) have changed the proportion of compatible trials, and/or (4) might have changed the strength of the S–R links between target and response in forced-choice trials. Any or all four of these differences could have caused a reduction in the NCE in forced-choice trials as (1) some kind of training might also reduce some kind of influence on the NCE (cf., Liu and Wang 2014; Schlaghecken et al. 2008), (2) the general RTs are associated with the exact time points when PCEs turn into NCEs (and possibly back to PCEs again; e.g., Bermeitinger and Wentura 2016; Panis and Schmidt 2016 ) and there are longer RTs in the current experiment than in previous experiments (e.g., Bermeitinger 2013), (3) compatibility effects are stronger with a larger amount of compatible trials (Bodner and Mulji 2010) and the adding of free-choice trials reduced the overall proportion of compatible trials in the current experiment, and/or (4) the strength of S–R links has been shown to modulate compatibility effects (e.g., Klauer and Dittrich 2010). Finally, it might be that the larger targets most often used in our previous experiments (e.g., Bermeitinger 2013; Bermeitinger and Wentura 2016; Eckert and Bermeitinger 2016; Hacklaender et al. 2015) actually reduced the primes’ visibility. In turn, the targets might moderate the degree of the compatibility effect—larger targets might lead to earlier and more pronounced NCEs. This could also explain the findings of Bermeitinger and Wentura (2016) who found earlier and larger compatibility effects with the larger targets (and row-of-dots primes) compared to smaller targets (and single-dot primes; but see, for example, Mattler and Palmer 2012, who found larger compatibility effects with open than full targets).
Turning to free-choice responses, we found similar results to our previous research (Bermeitinger 2013; Bermeitinger and Wentura 2016; Hackländer et al. 2015). Specifically, the NCE of M = − 10 ms (d = 0.76) at the longer SOA of 360 ms was highly comparable to previous findings (NCEs at SOA = 350/360 ms varied from M = −10 to − 17 ms, d = 0.61–1.05 across experiments; Bermeitinger 2013). That is, we found slower responses after congruent than incongruent primes, even when participants could freely choose which key they wanted to press in response to the ambiguous target. This seems to be clear evidence that the response to the target does not have to be instructed and strong S–R links between a given target and a response are not necessary for the negative effect. In general, the pattern also matches previous studies with free-choice priming in which in almost all cases the results of free-choice priming complied with the results of forced-choice priming (e.g., Perry and Lupker 2010). Without an instructed response and with ambiguous targets, there is no conflict between prime (response) and target (response) and accounts which explain compatibility effects by such a conflict (cf., Botvinick et al. 2001; Schmidt et al. 2011b) might not be applicable. Mattler and Palmer (2012) formulated a simple accumulator model for forced- as well as free-choice priming. The model assumes that response selection is based on and/or influenced by the integration of information. Information is coming from various sources of evidence. In the case of forced-choice priming, information from the prime and the target is integrated. In free-choice priming, information from the prime and internal response tendencies is integrated. The model does not accommodate negative effects. Thus, for NCEs additional processes have to be assumed. In general, negative compatibility effects can be seen as reflecting coping strategies to deal with inappropriately elicited activations (see also Machado et al. 2007, 2013) or as over-corrections for the influence of the prime (Klauer et al. 1997). These strategies or correction processes seem to be at work even in free-choice situations.
There is evidence that negative compatibility effects specifically occur with slower responses (e.g., Ocampo and Finkbeiner 2013; Schmidt et al. 2015; but see Bermeitinger and Wentura 2016). This has been interpreted as evidence that regulatory feedback processes take some time before they can influence pre-activations and facilitations that are caused by the prime and processed in a pure feedforward manner. Thus, one might wonder whether participants might adapt their response strategies and respond generally slower in longer-SOA conditions, thereby driving the negative compatibility effect. However, there is no indication for a general increase in response times with SOA, as can be seen from the neutral condition (see Table 1), indicating that participants did not generally use different response strategies. However, one might argue that the generally slower responses in free-choice trials allow slower regulatory (feedback) processes to come into play more than in forced-choice trials which are generally responded to faster.
We generally asked what differences and similarities are between forced-choice and free-choice trials and between the results from our experiment and the results from others on comparisons of forced- and free-choice trials. In the current experiment, we found no correlation between compatibility and congruency effects. This lack of a correlation fails to confirm previous findings with static masked primes (Bodner and Mulji 2010; Klapp and Hinkley 2002; Ocampo 2015) that were interpreted as evidence of a common origin of the two forms of priming. This lack of a correlation also fits with a recent experiment that used masked primes and compared forced- and free-choice trials (Le Bars et al. 2016, Experiment 1). In this case the authors found a different pattern of compatibility effects between forced- and free-choice trials (in terms of RTs) when the prime preceded the target by 132 ms. Despite these findings, we would not necessarily interpret this as evidence that different processes underlie responses in forced- and free-choice trials. For one, we clearly found the same pattern of results on both types of trials in our experiment, which is typical in the literature (e.g., Perry and Lupker 2010). Furthermore, even though free- and forced-choice trials lead to differences in absolute RTs, the fact that they are both susceptible to negative (congruency/compatibility) effects fits nicely with the literature suggesting that both types of responses are based on the same basic mechanisms (Janczyk et al. 2014; Naefgen et al. 2017), a notion that has partially been evidenced by equal susceptibility to interference effects (Janczyk et al. 2014).
As a separate point of discussion, one might wonder why we did not find any evidence for influences of the prime’s direction on the frequency of chosen responses in free-choice trials. This seems to be in strong contrast to previous experiments which even found influences on the frequency of the chosen responses after subliminal primes (e.g., Kiesel et al. 2006). However, there are also some other studies in which no influence of masked primes on the chosen response in free-choice trials was found (e.g., Mattler and Palmer 2012, Exp. 2 with arbitrary shapes). Bodner and Mulji (2010) reported an influence of the proportion of compatible trials in forced-choice trials, even on the effects in free-choice trials. Interestingly, they found no significant influence on the frequency of the chosen response in free-choice trials with a low proportion (0.2) of compatible trials, but only with a high proportion (0.8). However, they did not test a medium proportion (0.5) and they also did not use neutral trials. The current experiment included only a low proportion of compatible trials, which could explain the lack of an effect on the frequency of chosen responses in free-choice trials. Further, the fact that our primes are visible might lead to differences between our results and the results of previous studies with masked primes. It is known from studies on mere exposure that the influence of a prime stimulus is more pronounced if the prime is presented subliminally than when participants are aware of the prime (for a meta-analysis see Bornstein 1989). It seems to be that the conscious awareness of stimuli leads to conscious processes which counteract their influences by intentions (see also the comparably longer reaction times in free-choice than forced-choice trials).
Finally, the question remains, as to which theoretical accounts may explain the general pattern of results from the current experiment. As already introduced above, mask-triggered inhibition as well as object-updating accounts can be excluded. In contrast, self-inhibition by the motor system (e.g., Eimer and Schlaghecken 2003) might be one possible mechanism behind the pattern. That is, in response to inappropriately elicited activations, the motor system releases inhibitory activations in order to cope with these activations (see also Machado et al. 2007, 2013).
In addition to an account based on motor inhibition, accounts focusing on attentional processes may also help to explain the observed results. For example, it could be that motion primes, in comparison with static primes, lead to longer and more sustained activations, possibly because of the greater relevance and salience of motion. This could result in a longer temporal frame in which PCEs occur. What is more, apparent motion and spatial attention are closely related (e.g., Xu et al. 2013). At the neural level, spatial attention can modulate inputs in terms of their location (but not their identity) at early stages of processing (i.e., P1/N1 components with onsets from 70 to 80 ms). On the other hand, analyses on the basis of non-spatial features, such as shape or color, are associated with later components (i.e., “selection negativity” SN with onsets between 140 and 180 ms; for a review, see, for example, Hillyard and Anllo-Vento 1998). Given that, it follows that spatial attention may be rapidly shifted in the direction of a motion, which, in the current situation, could enhance or speed processing of the target in compatible trials. Note, however, that this attentional mechanism cannot be the sole cause of the effect (see discussion above), but rather only a moderator.
The compatibility effects may also be explained as a product of evidence accumulation for one of two response options (e.g., Klauer and Dittrich 2010). In this case, the motion primes, which may involve spatial attention shifts, allow for more time for evidence accumulation than do static primes. This could lead to more sustained compatibility effects with motion than static primes. It is worth noting that an explanation of compatibility effects such as this, which relies on shifts of spatial attention, would not be available for shape or color primes and could not account for effects in other priming paradigms (e.g., semantic priming).
In summary, in the present research response priming with clearly visible directional motion primes and static arrow targets was examined. Specifically, the targets were adapted to the targets often used in response priming with static primes. Further, we were interested in the question as to how motion influences responses even in free-choice situations. Thus, we used ambiguous targets (<>, ><) to which participants could freely choose to respond with the left or right key, interspersed into instructed (forced-choice) targets (<<, >>) to which participants had to press the assigned key (left; right). The general pattern of effects (in RTs) was highly comparable to previous research, even in free-choice trials: Positive compatibility effects occurred with a short SOA and negative compatibility effects with a long SOA. Several theoretical accounts have been proposed to explain response priming effects. Some accounts can be excluded for the present data; this especially includes accounts referring to mask-triggered inhibition or object updating, as well as some attentional accounts which assume strong differences between larger and smaller targets. In contrast, the current pattern of results is most likely best explained by a combination of (1) self-inhibition of the motor system, (2) the evaluation window account, and (3) some specific attentional mechanisms moderating the effects.
Notes
There were no differences between the inward and outward prime motion (ps between 0.082 and 0.588).
References
Berlyne DE (1957) Conflict and choice time. Br J Psychology 48:106–118
Bermeitinger C (2013) Response priming with apparent motion primes. Psychol Res 77:371–387
Bermeitinger C, Wentura D (2016) Moving single dots as primes for static arrow targets: negative effects at very long SOAs. Exp Psychol 63:127–139
Bodner GE, Mulji R (2010) Prime proportion affects masked priming of fixed and free-choice responses. Exp Psychol 57:360–366
Bornstein RF (1989) Exposure and affect: overview and meta-analysis of research, 1968–1987. Psychol Bull 106:265–289
Bosbach S, Prinz W, Kerzel D (2005) Movement-based compatibility in simple response tasks. Eur J Cogn Psychol 17:695–707
Botvinick MM, Braver TS, Barch DM, Carter CS, Cohen JD (2001) Conflict monitoring and cognitive control. Psychol Rev 108:624–652
Burle B, Allain S, Vidal F, Hasbroucq T (2005) Sequential compatibility effects and cognitive control: Does conflict really matter? J Exp Psychol Hum Percept Perform 31:831–837
Cole GG, Kuhn C (2010) What the experimenter’s prime tells the observer’s brain. Atten Percept Psychophys 72:1367–1376
Eckert D, Bermeitinger C (2016) Response priming with more or less biological movements as primes. Atten Percept Psychophys 78:1414–1433
Egeth HE, Yantis S (1997) Visual attention: control, representation, and time course. Annu Rev Psychol 48:269–297
Eimer M, Schlaghecken F (2002) Links between conscious awareness and response inhibition: evidence from masked priming. Psychon Bull Rev 9:514–520
Eimer M, Schlaghecken F (2003) Response facilitation and inhibition in subliminal priming. Biol Psychol 64:7–26
Enns JT, Di Lollo V (2000) What’s new in visual masking? Trends Cogn Sci 4:345–352
Farroni T, Massaccesi S, Pividori D, Simion F, Johnson MH (2004) Gaze following in newborns. Infancy 5:39–60
Faul F, Erdfelder E, Lang A-G, Buchner A (2007) GPower 3: a flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav Res Methods 39:175–191
Hacklaender R, Eckert D, Bermeitinger C (2015) Visual processing: the role of importance. In: Heinen T (ed) Advances in visual perception research. Nova Publishing, New York, pp 1–28
Hillyard SA, Anllo-Vento L (1998) Event-related brain potentials in the study of visual selective attention. Proc Natl Acad Sci 95:781–787
Janczyk M, Dambacher M, Bieleke M, Gollwitzer PM (2014a) The benefit of no choice: goal-directed plans enhance perceptual processing. Psychol Res 79:206–220. https://doi.org/10.1007/s00426-014-0549-5
Janczyk M, Nolden S, Jolicoeur P (2014b) No differences in dual-task costs between forced- and free-choice tasks. Psychol Res 79:463–477. https://doi.org/10.1007/s00426-014-0580-6
Kiesel A, Wagener A, Kunde W, Hoffmann J, Fallgatter A, Stöcker C (2006) Unconscious manipulation of free choice in humans. Conscious Cogn 15:397–408
Kiesel A, Kunde W, Hoffmann J (2007) Mechanisms of subliminal response priming. Adv Cogn Psychol 3:307–315
Kiesel A, Berner MP, Kunde W (2008) Negative congruency effects: a test of the inhibition account. Conscious Cogn 17:1–21
Klapp S, Haas BW (2005) Nonconscious influence of masked stimuli on response selection is limited to concrete stimulus-response associations. J Exp Psychol Hum Percept Perform 31:193–209
Klapp ST, Hinkley LB (2002) The negative compatibility effect: unconscious inhibition influences response selection. J Exp Psychol Gen 131:255–269
Klauer KC, Dittrich K (2010) From sunshine to double arrows: an evaluation window account of negative compatibility effects. J Exp Psychol Gen 139:490–519
Klauer KC, Roßnagel C, Musch J (1997) List-context effects in evaluative priming. J Exp Psychol Learn Mem Cogn 23:246–255
Krüger D, Klapötke S, Bode S, Mattler U (2013) Neural correlates of control operations in inverse priming with relevant and irrelevant masks. NeuroImage 64:197208
Le Bars S, Hsu Y, Waszak F (2016) The impact of subliminal effect images in voluntary versus stimulus-driven actions. Cognition 156:6–15
Liu P, Wang Y (2014) Perceptual and motor contributions to the negative compatibility effect. Acta Physiol (Oxf) 153:66–73
Lleras A, Enns JT (2004) Negative compatibility or object updating? A cautionary tale of mask-dependent priming. J Exp Psychol Gen 133:475–493
Machado L, Wyatt N, Devine A, Knight B (2007) Action planning in the presence of distracting stimuli: an investigation into the time course of distractor effects. J Exp Psychol Hum Percept Perform 33:1045–1061
Machado L, Guiney H, Struthers P (2013) Identity-based inhibitory processing during focused attention. Q J Exp Psychol 66:138–159
Mattler U, Fendrich R (2007) Priming by motion too rapid to be consciously seen. Percept Psychophys 69:1389–1396
Mattler U, Palmer S (2012) Time course of free-choice priming effects explained by a simple accumulator model. Cognition 123:347–360
Naefgen C, Dambacher M, Janczyk M (2017) Why free choices take longer than forced choices: evidence from response threshold manipulations. Psychol Res. https://doi.org/10.1007/s00426-017-0887-1
O’Connor PA, Neill WT (2011) Does subliminal priming of free response choices depend on task set or automatic response activation? Conscious Cogn 20:280–287
O’Craven K, Rosen BR, Kwong KK, Triesman A, Savoy RL (1997) Detecting the effects of voluntary attention on a visual motion processing region in human cortex. Neuron 18:591–598
Ocampo B (2015) Unconscious manipulation of free choice by novel primes. Conscious Cogn 34:4–9
Ocampo B, Finkbeiner M (2013) The negative compatibility effect with relevant masks: a case for automatic motor inhibition. Front Psychol 4:822. https://doi.org/10.3389/fpsycg.2013.00822
Panis S, Schmidt T (2016) What is shaping RT and accuracy distributions? Active and selective response inhibition causes the negative compatibility effect. J Cogn Neurosci 28:1651–1671
Parkinson J, Haggard P (2014) Subliminal priming of intentional inhibition. Cognition 130:255–265
Perry JR, Lupker SJ (2010) A prospective view of the impact of prime validity on response speed and selection in the arrow classification task with free choice trials. Attention Percept Psychophysics 72:528–537
Sarkheil P, Vuong QC, Bülthoff HH, Noppeney U (2008) The integration of higher order form and motion by the human brain. NeuroImage 42:1529–1536
Schlaghecken F, Eimer M (2004) Masked prime stimuli can bias ‘free’ choices between response alternatives. Psychon Bull Rev 11:463–468
Schlaghecken F, Bowman H, Eimer M (2006) Dissociating local and global levels of perceptuo-motor control in masked priming. J Exp Psychol Hum Percept Perform 32:618
Schlaghecken F, Blagrove E, Maylor EA (2008) No difference between conscious and nonconscious visuomotor control: evidence from perceptual learning in the masked prime task. Conscious Cogn 17:84–93
Schlaghecken F, Klapp ST, Maylor EA (2009) Either or neither, but not both: locating the effects of masked primes. Proc R Soc B 276:515–521
Schmidt F, Haberkamp A, Schmidt T (2011a) Dos and dont’s in response priming research. Adv Cogn Psychol 7:120–131
Schmidt T, Haberkamp A, Veltkamp GM, Weber A, Seydell-Greenwald A, Schmidt F (2011b) Visual processing in rapid-chase systems: image processing, attention, and awareness. Front Psychol. https://doi.org/10.3389/fpsyg.2011.00169
Schmidt T, Hauch V, Schmidt F (2015) Mask-triggered thrust reversal in the negative compatibility effect. Atten Percept Psychophys 77:2377–2398. https://doi.org/10.3758/s13414-015-0923-4
Simion F, Regolin L, Bulf H (2008) A predisposition for biological motion in the newborn baby. Proc Natl Acad Sci 105:809–813
Treue S, Maunsell JHR (1996) Attentional modulation of visual motion processing in cortical areas MT and MST. Nature 382:539–541
Tukey JW (1977) Exploratory data analysis. Addison-Wesley, Reading
Verleger R, Jaskowski P, Aydemir A, van der Lubbe RHJ, Groen M (2004) Qualitative differences between conscious and nonconscious processing? On inverse priming induced by masked arrows. J Exp Psychol Gen 133:494–515
Wyatt N, Machado L (2013) Evidence inhibition responds reactively to the salience of distracting information during focused attention. PLoS ONE 8:e62809
Xu Y, Suzuki S, Franconeri SL (2013) Shifting selection may control apparent motion. Psychol Sci 24:1368–1370
Acknowledgements
We thank Leonard Brauch for his help in data collection.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
All procedures performed in our experiments involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Declaration of Helsinki and its later amendments or comparable ethical standards. The experiment reported here was approved by the Local Ethical Committee of the University of Hildesheim.
Additional information
Handling editor: Anna Belardinelli (University of Tübingen); Reviewers: Brenda Ocampo (University of Queensland), Markus Janczyk (University of Tübingen).
Rights and permissions
About this article

Cite this article
Bermeitinger, C., Hackländer, R.P. Response priming with motion primes: negative compatibility or congruency effects, even in free-choice trials. Cogn Process 19, 351–361 (2018). https://doi.org/10.1007/s10339-018-0858-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10339-018-0858-5