
Abstract
Dispersal to efficiently locate future breeding areas in young animals has important consequences for survival, life history and future breeding success, and therefore population dynamics and evolution. Long-distance migrant birds that have little time immediately pre-breeding may use the post-fledging period to locate their future breeding territory. We radio-tracked 37 juvenile Cyprus Wheatears, Oenanthe cypriaca, a long-distance migrant passerine, to investigate movements for 10 weeks post-fledging. We measured how distance from the nest and distance of consecutive movements changed with age. We tested whether distance from the nest stopped increasing and consecutive movement distance became similar to pre-dispersal distances, consistent with a fledgling adopting a post-fledgling home range that could reflect scoping out a future local breeding territory. Fledglings had a very high survival rate. Directed movements away from the natal territory started at about 18 days until about 45 days when individuals adopted a more fixed location on average about 600 m (range 0–1500) from their natal site, and further movements were at a scale equivalent to movements within an adult-sized breeding territory. Our results suggest dispersal followed by settlement in a fixed home range prior to first migration that could function to identify the breeding site location for the following year.
Zusammenfassung
Ortsveränderungen flügger Zypernsteinschmätzer Oenanthe cypriaca: Belege für Streifgebiete abseits des Geburtsortes zwischen dem Ausfliegen und dem ersten Wegzug
Dismigration bei Jungtieren zur effizienten Suche nach zukünftigen Brutgebieten hat bedeutende Auswirkungen auf Überleben, Biologie sowie späteren Bruterfolg und damit auf die Populationsdynamik und die Evolution. Langstreckenzieher, die unmittelbar vor der Brutsaison nur wenig Zeit zur Verfügung haben, können die Phase nach dem Ausfliegen nutzen, um ihr zukünftiges Brutrevier zu finden. Wir telemetrierten 37 juvenile Zypernsteinschmätzer, Oenanthe cypriaca, eine langstreckenziehende Singvogelart, um Ortsveränderungen während zehn Wochen nach dem Ausfliegen zu untersuchen. Wir quantifizierten, wie sich die Entfernung vom Nest und die Strecke der aufeinanderfolgenden Ortsveränderungen mit dem Alter wandelten. Wir prüften, ob die Entfernung vom Nest nicht weiter zunahm und ob sich das Ausmaß anschließender Ortsveränderungen dem Niveau vor Einsetzen der Dismigration annäherte, wie dies für einen flüggen Jungvogel zu erwarten wäre, der nach dem Ausfliegen ein Streifgebiet nutzt, welches ein Zeichen für die Suche nach einem zukünftigen Brutrevier im Umkreis sein könnte. Wir untersuchten, in wie fern sich der Abstand zum Nest sowie die nachfolgenden Ortsveränderungen mit dem Alter ändern. Flügge Jungvögel hatten eine sehr hohe Überlebensrate. Zielgerichtete Ortsveränderungen fort vom Geburtsort begannen nach zirka 18 Tagen bis nach etwa 45 Tagen, wonach sich die Vögel an einem konstanteren, im Schnitt etwa 600 m (zwischen 0–1500 m) vom Geburtsort entfernten Ort niederließen und sich die weiteren Ortsveränderungen in einem mit dem Brutrevier eines Altvogels vergleichbaren Rahmen abspielten. Unsere Ergebnisse deuten auf eine Dismigration, gefolgt von einem Niederlassen in einem festen Streifgebiet vor dem ersten Wegzug hin, das dazu dienen könnte, einen Brutplatz für das folgende Jahr zu finden.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The post-fledging period represents a time of intense selective pressure for birds with juveniles typically experiencing high mortality rates (Morton 1992; Grüebler et al. 2014; Naef-Daenzer and Gruebler 2016) as they develop and disperse into novel areas (Anders et al. 1997; Cox et al. 2014). Despite the fundamental importance of the post-fledging dispersal period, it remains understudied in comparison to the other stages of avian development (Cox et al. 2014). The main reason for this is the difficulty of tracking and observing young after they leave the natal area (Kershner et al. 2004b) and the large spatial scale they can range over (Sutherland et al. 2000).
The post-fledging period has been shown to be dynamic rather than a singular unified stage of life: as fledglings develop, the selective pressures they experience change due to increased mobility and knowledge of the foraging environment and predators (Raybuck et al. 2020). Fledglings remain within the natal area within the first few weeks after fledging and are at their most vulnerable to predation (Moore et al. 2010; Ausprey and Rodewald 2011; Eng et al. 2011). However, as fledglings develop and their mobility increases, they are able to utilise different habitats that are potentially distant, which could represent a balance between predator protection (Morton 1991; Anders et al. 1998; King et al. 2006) and optimal food availability (Vega Rivera et al. 1998; Vitz and Rodewald 2007). Reduction in the chance of predation as age increase may cause fledglings to shift their post-fledging strategy from mainly predator avoidance towards foraging (Streby et al. 2011) or locating future breeding territories (Mitchell et al. 2010).
Mechanisms that allow the location of suitable breeding areas are likely to be under strong selection (Kokko 1999). This may be particularly intense in long-distance migrants which spend the non-breeding season far from their breeding areas (Lombardo 1987, e.g. Arlt and Part 2008). They then lack the opportunity to locate breeding areas as adults except during the short period after arrival back on the breeding grounds in the spring, when early location and occupation of a territory increases fitness (Morrison et al. 2019, e.g. Aebischer et al. 1996, Currie et al. 2000, Velmala et al. 2015). Therefore, there is likely to be strong selection for potential breeding territories to be identified by juveniles in the post-fledging period before first migration, and this may be one of the reasons why juveniles also tend to migrate later than adults (Newton 2008). Location of future breeding territories during this period may also allow the quality of potential future territories to be assessed through the breeding density, nesting site availability or success of conspecifics (Lombardo 1987; Nocera et al. 2006; Betts et al. 2008), or through food availability. If the post-fledging period of long-distance migrants functions as a period for potentially locating a breeding site for the following year, we might then expect fledglings to disperse but then settle in fixed locations away from the natal site, but suitable for breeding.
Processes other than locating future breeding sites may also lead to patterns of post-juvenile dispersal in migrant birds. The migration commencement hypothesis suggests that movements by fledglings are orientated in the direction that facilitates eventual autumn migration (Rappole and Ballard 1987). The habitat optimisation hypothesis suggests that fledglings select areas to maximise access to food or to avoid predators (Streby et al. 2011). The homing target formation hypothesis suggests that fledglings seek out large landscape features that will help them recognise either their natal area or dispersal area during their spring migration (Wiltschko and Wiltschko 1978). Patterns could even be generated by a random process of dispersal followed by home range formation, where individual fledglings have a random direction, distance and duration of dispersal, followed by feeding in a relatively small area up until migration. However even if this process is random, then selection should favour those individuals that return to good areas to breed. It is likely that several or all hypotheses may account for the movements recorded during the post-fledging period, acting at different spatial and temporal scales.
Here, we measure the post-fledging movements of Cyprus Wheatear, Oenanthe cypriaca, to determine whether they are consistent with the prospecting for future breeding locations hypothesis. Cyprus Wheatears are widespread and common migrants (Randler et al. 2010) endemic to Cyprus (Randler et al. 2012) that winter in sub-Saharan East Africa from southern Sudan to Ethiopia (Xenophontos et al. 2017). Measuring space use during the post-fledging period and territory adoption the following year is much more tractable than in many systems. Most juvenile Cyprus Wheatears (92%) have been found to breed in the year after fledging less than 1 km away from their natal site and to have annual survival rates that may exceed 35% (Xenophontos and Cresswell 2016b), although recent work has suggested that both of these measures are likely to vary annually (Patchett unpublished data). Survival in the first 4 weeks post-fledging is particularly high (> 95%, Xenophontos and Cresswell 2016a). Previous observations suggest that juveniles move short distances before settling in potentially suitable breeding habitat before their first migration.
We measured how direction and distance from the nest site varied with age over a 70-day period post-fledging to determine if there was any evidence for adoption of a post-fledging home range after initial dispersal during this period. We assumed that when juveniles begin moving distances that exceed the average diameter of a breeding territory that they are dispersing. We then examined the variation in the direction and distance from their natal site, and distance between consecutive radio-tracked locations. This allowed us to determine whether distance from the natal site became fixed, and daily movements became smaller as would be expected with adoption of a post-fledging home range. If individual juveniles disperse (leave their natal territory) completely randomly in terms of direction and distance, then distances from the natal site will inevitably decrease (because all movements initially must be away from the natal site, whereas as an animal moves further away an increasing proportion of movements will be movements back to the natal territory). Consecutive distances, however, will not change. If individual juveniles disperse randomly with respect to distance but not direction, they will continue to increase in distance from their natal site, and consecutive distances will remain similar. However, if juveniles adopt a post-fledging home range, then distance from the natal territory will level off, and then show smaller consecutive movement distances. Post-dispersal movement distances are then likely to be similar to those at the natal site if fledglings are restricting their movements to an area equivalent to an adult breeding territory. In practical terms, dispersal away from the natal territory, followed by adoption of a home range equivalent in size to the natal site, will lead to a sigmoidal relationship between distance away from the nest and age, and a quadratic relationship (with a maximum during the dispersal phase) between consecutive distances between locations (step lengths) and age. Furthermore, if post-fledging dispersal leads to home ranges corresponding to future breeding locations, we would expect birds to spread out rather than aggregate, and to settle in suitable breeding habitat.
Here, we first test whether individual juvenile Cyprus Wheatears disperse in a consistent direction and whether they ended spatially separated at a scale larger than an individual adult territory, within potentially suitable breeding areas. Having established this, we then show that after larger-scale movements away from the natal site, individuals are likely to stop moving away from the natal area, and restrict their movements to a smaller scale, equivalent to them using adopting a home range similar in size to an adult breeding territory. Finally, we determine the average duration of the dispersal phase and how long after leaving the natal territory juveniles settle into a more fixed home range.
Methods
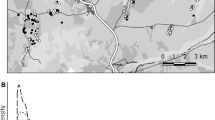
The study was conducted during the breeding season of 2019 in a 130-ha area at Troodos National Forest Park (34° 56011 N 32° 51048 E: Fig. S1), at about 1800 m a.s.l. on Cyprus. The study area was within an old, low-density coniferous forest, supporting one of the densest breeding Cyprus Wheatear populations on the island (Flint and Stewart 1992). The park covers an area of 9029 ha with its highest peak at 1952 m. Habitats in the study area include Pinus nigra ssp. pallasiana forest, Juniperus foetidissima woodland and Serpentinophilous grasslands. Annual precipitation in the area is high but restricted to the winter period; summers are very dry and warm with afternoon temperatures reaching 35 °C.
From May through June 2019, we opportunistically found Cyprus Wheatear nests through close observation of adult behaviour such as nest building and feeding chicks. We monitored nests every 3–4 days and every 1–2 days as fledging approached (fledging occurs at approximately 14 days after hatching). Cyprus Wheatear fledglings were captured in mist nests or spring-traps within 25 days of fledging in their natal territory: the average age at tagging was 13.5 days (± 1.0 SE; range 1–25 days, where day 0 is the day of fledging). Only broods which were certain to have been produced within a territory, with attendant colour-ringed parents, were targeted. Colour ringed parent birds were seen providing parental care to the fledglings in almost all cases so that their parentage was known at the time of capture, and where broods were targeted after the second week after fledging these were from the earliest few nests on the study site, with the broods and their colour-ringed parents being confirmed as remaining within the natal territory before capture was attempted.
Each chick was ringed (with permission from the Game Fund, Ministry of Interior and BirdLife Cyprus) with a unique combination of a single metal ring and three colour rings. It was not possible to ring Cyprus Wheatears in the nest, because access to the nest cavity would have destroyed the nest site. Additionally, we attached a 0.42 g Ag379 radio-transmitter (Biotrack, Wareham, UK) with a 13 cm whip antenna and a battery life of c. 50 days. Tags were attached using a modified leg-loop harness design (Rappole and Tipton 1991; Streby et al. 2015). The harnesses were constructed of 0.5 mm elastic thread that would naturally degrade over 3 months allowing the radio tag to fall off prior to migration. Birds weighed on average 15.82 g when ringed and the tags with harness on average weighed 0.49 g, which equals 3.1% of body mass.
We aimed to catch every chick in a brood (typical brood size is 5, Xenophontos and Cresswell 2016b) with a ratio of three with radio-transmitter tags to two with colour rings only. In many cases, only some chicks from the brood could be caught because juveniles avoided nets or spring traps. In instances, where only four birds were caught, three fledglings were fitted with radio-transmitters and one with rings only. A single fledgling was tagged in seven broods, two fledglings were tagged in six broods, and three fledglings were tagged in six broods. In total, there were 15 fledglings colour ringed only from nine broods. There were no obvious tag effects in the study—no obvious mortality was recorded (see below). Birds fitted with tags flew off in the same manner as birds that had only colour rings. Birds with tags were seen in subsequent days feeding and behaving identically to colour ringed only birds from the same brood. Colour ringed only chicks were also seen throughout the extended study area (see below) while locating the tagged chicks suggesting there was no obvious tag effect on dispersal distances. We had no evidence of chick mortality prior to the catching period. For almost all of the broods we have complete nest information. The number of chicks present in the natal territory receiving parental care at the time of catching was the same as the number of chicks recorded prior to fledging.
We defined an average natal territory size (for the purpose of classifying when juveniles started to disperse and move away from their natal territory) by calculating the mean distance between nests. We measured the distance between 12 adjacent nests in the central area of the study site where all territories were known and surrounded by other territories. We then used half the mean distance between the 12 nests to define an average natal territory size. This had a radius of 175 m (± 1 SE 45.4 m).
The study site was divided into sections based upon topography and local features. As juveniles began to disperse, a 300 m buffer was added to the central field site where fledglings were tagged. A further 300 m buffer was also searched for any birds that we thought had dispersed from the central field site and that were not located in the first buffer zone. This occurred 5 weeks after the first tag was fitted after it became apparent that at least four birds had moved out of the first buffer area.
Locations were collected daily but each section of the study site was visited only once every 3–8 days, resulting in 15 discrete sampling events during the study covering the entire study site. Individual birds could be recorded in different sections during a sampling event, or not located, leading to greater than or fewer than 15 locations being recorded, respectively. The mean sampling interval per individual was 5.8 (± 0.4 SE) days: there was no relationship between age of an individual and sampling interval (0.0074 ± 0.026 SE, t181 = 0.3, P = 0.78; sampling interval ~ age with individual identity as a random effect). Because of this even pattern of sampling with age, sampling interval was not considered as a confounding effect when examining distance from the nest or distance between consecutive locations.
In each section, five or ten (to reflect the area of the section) random sampling points were calculated using the QGIS v3.4 (QGIS Development Team 2019. QGIS Geographic Information System. Open Source Geospatial Foundation Project, http://qgis.osgeo.org) random location generator. Random points, rather than a regular grid, were used to potentially sample all locations within the unevenly shaped areas. For each observation day, new random points were created, if the random points were adjacent and there was < 5 m distance between them, the additional random point was removed from that day’s observations. At each sampling point, we scanned for the radio frequencies of each radio tag for a period of 10 s each. A programmable SIKA receiver attached to a three-element Yagi antenna was used to locate individuals and then their location was visually confirmed and recorded using GPS. Where multiple signals were detected, we located the first detected individual before returning to the sampling point to find the next detected bird. This was carried out until all detected frequencies were located.
The average tracking time was 37.4 (± 12.3 SE) days, with a range of 1–70 days post-fledging tracked. An average of 6.9 (± 1.1 SE) locations were taken per individual. All tracked locations are mapped in Fig. S1. Of the 37 birds fitted with tags (Fig. S2), 21 dispersed from their natal territory to settle within the study area but away from their natal territory, five moved from their natal territory and then returned to it and two birds did not disperse from their natal territories. Five birds had their tags fail a few days after fitting giving limited data. In four out of five of these cases which we classified as tag failure, tags transmitted a distorted signal in the days before the tag stopped transmitting, indicating an impending tag failure and the four birds were then re-sighted carrying non-functional tags. The final four birds were tracked moving away from their natal area and then were lost: these were assumed to have dispersed completely outside the study area. One bird that was thought to have dispersed in this way was then re-found outside the extended field site boundary 1.1 km from its natal territory, confirming this assumption. But we do not know for certain what happened with four individuals (one assumed tag failure and three assumed dispersed out of the study area). For the assumed tag failure, the bird was not seen again after the day of tagging and may have died. For the three assumed dispersed out of the study area, we do not think the tags failed because there was no indication in the signal noise received on last detection that the tag was failing. Some tags stopped functioning near the end of the study as their batteries were exhausted. We were able to discriminate between end of battery life and mortality because even after the battery of the tag had died, fledglings remained in the same approximate area of the last transmission. For 13 birds where the tag functioned as expected but the battery ran out before the end of the study, 12 birds were resighted in the same approximate location after their tag stopped transmitting and one that was not initially found in the same location after tag transmission ceased was resighted 4 weeks later alive with the tag present.
Statistical analysis
First, we tested whether individuals dispersed in similar or random directions from their natal territory. Uniformity of the direction of each location of a juvenile from its nest site, after it left its natal area, was tested with a Rayleigh test to determine if individuals dispersed in a consistent direction. We then tested whether the mean distance between birds increased with age to determine whether individuals dispersed to spatially separated sites. To do this without the confounding effects of chicks within and between broods, we calculated the mean location of each bird in a brood, and then the mean of these within brood locations for each brood, for different ages of fledglings pooled into weeks. The mean of all possible distances between pairs of brood locations (i.e. their average spacing) was then calculated for each week.
Second, we calculated distance moved with age. For every sampling event, the distance from the nest, and the distance between it and the last location was calculated for every individual tracked. All distances were calculated as great circle distances using the distHaversine function from the geosphere package in R. We then plotted these two distances with age and modelled the best fit function, expecting that distance from the nest would take a sigmoidal function, and distance travelled between consecutive locations a quadratic function, if fledglings dispersed to a new home range within the study area as if scoping out future breeding territories. We modelled distance with age on either raw data, with individual identity as a random factor in a mixed model framework to control for uneven sampling across individuals, or, for visualisation, aggregated into one mean for each individual within a weekly period.
Having established, with the analysis above, that there was a pattern of dispersal where individuals moved a lot (dispersal) preceded by a period in the natal territory (pre-dispersal) and followed by a period in a home range when they moved much less (post-dispersal), we then determined the timing of these periods. We varied the relative duration of the three dispersal periods to determine which combination of durations best predicted variation in distance in a simple factorial model. The data were effectively subset into the three different periods (pre-dispersal, dispersal and post-dispersal). The end of the pre-dispersal period was varied from 10 to 25 days, and the end of the dispersal period from 35 to 65 days. A model was then run for distance predicted by each range of period durations, and the model with the lowest AIC value was chosen. The process was iterative, varying relative duration of the periods until the lowest AIC value was obtained. The model tested in each case was then: distance measure ~ period, with period as a three-way factor; sample size was the number of individuals tracked, with each individual contributing one mean value per period; bird identity was added as a random factor because some individuals contributed a mean in all periods, and other only in some. Birds with only one record in a period were removed from the analysis. Within each period, the mean distance between locations, and its standard error, were calculated from all possible pairwise distances between locations in a period, for each individual separately. Maximum distance was calculated as the maximum possible distance between any pair of locations recorded in a period, for each individual separately.
Analysis was carried out using generalised linear mixed models (glmm), general additive models (gam), and nonlinear models (nls) using the libraries nlme, lmertest, MuMIn and mgcv in R. Model fit was assessed by visual inspection of residuals plotted against fitted values and quantile plots and were reasonable in all cases, apart from models with distance moved between consecutive locations as a dependent variable: these models had reasonable assumptions after log-transforming distance. Predicted values were plotted using the lme4, AICcmodavg and ggplot2 packages in R; variance was partitioned into main and random effects in mixed models using MuMIn.
Results
Survival rate was high. Survival rate of the 37 fledglings tagged varied from a minimum of 89.2% (assuming the four individuals that could not be accounted for died) to a maximum of 100% (assuming the four individuals that could not be accounted for dispersed completely out of the study area). Individuals were tracked for approximately 35 days on average, over a 2-month period after fledging, giving a minimum daily survival rate during this period of 0.997, and a maximum survival rate of 1.
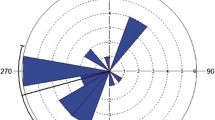
Fledglings showed consistency in direction from their nest site, but they dispersed in different directions. The directions moved by individual fledglings was highly directional (Rayleigh’s test P < 0.001 in all cases). Fledglings (N = 19 broods) became more separated with age, rather than more clustered, more than doubling their average distance apart over the 10-week study period (the increase in the mean value of all possible paired distances between average locations for a brood was 51.4 ± 12.2 SE meters per week t1,8 = 4.2, P = 0.003: Fig. 1, Fig. S1). At least 16% of fledglings occupied territories where adults were still breeding after dispersal within the main field site, and 20% of fledglings returned to their natal territory after initially dispersing several hundred meters away (Figs. S1, S2). All tagged individuals moved to potentially suitable breeding areas in the sense that all areas were typical breeding habitat and were part of breeding adult territories at least once during the longer term, ten year study (Fig. S1, Xenophontos and Cresswell 2016a,b; Xenophontos et al. 2017).

Average distance between all locations for individual brood means with week since fledging to show that that tracked fledglings became increasingly spaced apart with age
Distance from the nest increased with age before stabilising. A GAM with a single term of days since fledging, and a random effect of bird identity showed a sigmoidal pattern and explained 43% of the deviance (F4.5 = 40.6, P < 0.001, n = 255: Fig. 2a). When aggregated into 11 week-long periods, a sigmoidal function with an asymptote of 594 ± 51 SE meters (t = 11.7, P < 0.001), and the highest rate of change at 28.9 ± 2.9 SE days (t = 9.8, P < 0.001, with an s value of 0.12 ± 0.04 SE, t = 3.2, P = 0.014) significantly explained distance from the nest with age (Fig. 2b). A sigmoidal function was a better fit than a straight line fit (change in AIC with a linear fit + 2.51).

Distance (m) moved by Cyprus Wheatear fledglings from their nest site with age. a Raw data, each location is a point with individuals linked by lines; fitted curve is a GAM of distance with age, with individual as a random effect. b Data aggregated into weeks, the mean distance from the nest for an individual in a week was calculated and then the mean of all individual weekly means are plotted: fitted curve is the best fit sigmoidal function. The horizontal dashed line is the mean radius of the natal territory, 175 m
Distances moved between consecutive locations increased with age before decreasing again. A quadratic function of age explained some of the variation in distance moved by fledglings between successive locations, with individual identity as a random effect [log distance ~ (0.052 ± 0.024 SE × age, t181 = 2.1, P = 0.033) + (− 0.00098 ± 0.0003 SE × age2, t181 = − 2.7, P = 0.007) + (4.1 ± 0.4 SE, t181 = 11.2, P < 0.001): Fig. 3a] accounting for 13% of variance. When data were aggregated into 11 week-long periods the evidence for a quadratic effect was stronger [distance ~ (12.9 ± 4.2 SE × age, t = 3.0, P = 0.018) + (− 0.19 ± 0.06 SE × age2, t = − 3.1, P = 0.016) + (20.0 ± 64.0 SE, t = 0.3, P = 0.76): Fig. 3b] accounting for 47% of the variance.

Distance (m) moved by Cyprus Wheatear fledglings between successive locations with age. a Raw data, each location is a point with individuals linked by lines; fitted curve is a GLM with individual as a random effect. b Data aggregated into weeks, the mean distance between consecutive locations for an individual in a week was calculated and then the mean of all individual weekly means are plotted: fitted curve is the best fit quadratic function. The horizontal dashed line is the mean radius of the natal territory, 175 m
The best-supported model for the timing of dispersal indicated that birds remained in their natal territory until age 18 days (period 1), those that left their natal territory dispersed until 40–45 days (period 2) and adopted a more fixed location post 40–45 days (period 3): 18, 40 day periods AIC = 721, or 18,45 = 722; other periods gave less good model fit, e.g. 16,40 days AIC = 725; 20,40 = 724; 18,35 = 725; 18,50 = 730; Figs. 1, 2. During Period 2 birds showed a significantly greater distance between consecutive locations (Fig. 4a; Table 1), maximum distance between locations (Fig. 4b; Table 1), and variation in the mean distance travelled (Fig. 4c; Table 1) compared to during period 1. During period 3, distances moved between consecutive locations (Fig. 4a; Table 1), the maximum distance between locations (Fig. 4b; Table 1) and the variation in mean distance travelled (Fig. 4c; Table 1) were similar to those in period 1, before they started dispersing.

Boxplots of a mean distance travelled, b maximum distance and c standard error of the mean distance travelled between consecutive points in a period. Pre-dispersal, period 1, is up to 18 days post-fledging, period 2 was 19–40 days post-fledging, period 3 was 41 days post-fledging till end of tracking. N = 69 observations from 34 birds; average measures calculated for each bird within each period
Discussion
Post-fledging survival
Overall survival rates in the study were very high and there was little or even no mortality recorded throughout the tracking period: high post-fledging mortality is more typical in the first 3 weeks after fledging (Cox et al. 2014; Naef-Daenzer and Gruebler 2016). Given the lack of predators in the Troodos area and previous work where juvenile survival rates even in the first 4 weeks post-fledging were found to be at least 95% (Xenophontos and Cresswell 2016b), dispersal out of the field site for three of the four birds we could not account for seems reasonable. Similarly, considering we had 14% of tags that failed early, it seems reasonable that the remaining bird we could not account for suffered tag failure. Regardless, an absolute minimum approximate survival rate for fledglings in the first two months of 89% is exceptional, and real survival rate is likely to be higher than this. Post-fledging mortality is mainly attributed to predation (Anders et al. 1997), poor weather conditions (Anders et al. 1997) or poor competitive foraging skills (Maness and Anderson 2013). A likely explanation for the lack of mortality recorded in this study is the scarcity of rain (none recorded during the fledging period) or predators at the study site on Troodos. During the study, only one predation attack was witnessed (an unsuccessful hunt by a juvenile Goshawk Accipiter gentilis), and no Sparrowhawks Accipiter nisus or other likely avian predators were seen hunting birds on the study area (as in previous years at the the study site, Xenophontos and Cresswell 2016b).
Age at dispersal
Juvenile Cyprus Wheatears moved away from their natal sites as they got older consistent with patterns shown in other passerines (Kershner et al. 2004a; Berkeley et al. 2007; Vitz and Rodewald 2010). On average, birds did not start moving away from their natal territory until 18 days. This period probably represents the post-fledging dependence period. Whilst the duration of parental care was not empirically measured, it is likely that the period lasted close to 14 days as adult birds were not observed regularly feeding fledglings after this period (pers. obs.). Dispersal of fledglings in many temperate bird species may often be when their developing foraging skills allow independence (Yoerg 1998; Pedersen et al. 2013), but there may be many other factors involved (Bombieri et al. 2018). For example, in some high temperature years when only single broods of Cyprus Wheatear are produced, the post-fledging feeding period may be double this (Xenophontos and Cresswell 2016a). Juveniles started to feed for themselves after 7 days and appeared to be mostly foraging for themselves after 14 days (pers. obs.). Adult males were observed chasing any of their first brood remaining in their territory if a second breeding attempt was initiated (pers. obs.). This aggressive behaviour occurred approximately 3 weeks after fledging occurred, though it was only observed in two territories.
Post-fledging movement hypotheses
The results of this study are consistent with the breeding territory prospecting hypothesis (Lombardo 1987). Although the number of locations obtained from this study is too small to obtain robust final home range estimates, after about 40–45 days post-fledging most fledglings had stopped moving away from the nest and the distance moved between consecutive points became relatively small, consistent with movement within a fixed home range rather than continuing dispersal. Other studies that tracked juveniles during the independence phase have noted the formation of a post-fledging home range (Anders et al. 1998; Streby and Andersen 2012).The breeding territory prospecting hypothesis also suggests that some juveniles, at least, will select, in their year of fledging, a breeding territory that is already being used by adults on the basis of positive social information (Nocera et al. 2006) and we found at least 22% of juveniles to have their final locations in territories where breeding was still occurring. All juveniles ended up in areas where we have observed breeding pairs in at least 1 year of the long-term study. Some individuals, however, remained in their natal territories. This may reflect a choice by the fledglings to attempt to breed in their natal area the following year, or a lack of complete dispersal during the time period of the study. In the former case, these individuals (including those that dispersed but then returned to their natal territory) add much to the noise in the data set shown in Fig. 2 by reducing the gradient of the function of distance from the nest with age and its asymptote. In the latter case, any individuals that have not started or completed dispersal will reduce the gradients of the function of consecutive distance travelled with age shown in Fig. 3, and add to the larger variation shown in the third period in all panels in Fig. 4. Despite this variation reducing the statistical significance of any relationship, we still show a pattern on average that suggests most fledglings move away from their natal territory and settle in an equivalently sized area, in suitable breeding habitat over a 2-month period.
There was less evidence for the other hypotheses to account for post-fledging dispersal pattern. We observed no obvious clustering from tracked individuals as would be expected for the homing target hypothesis (Wiltschko and Wiltschko 1978), and no groups of fledglings as might be likely if they were seeking and ending up in the best foraging areas or the safest areas, as would be expected for the habitat optimisation hypothesis (see Streby et al. 2011). Given the short distances involved, the migration commencement hypothesis also does not seem relevant for our observations (see Rappole and Ballard 1987). There was also no strong suggestion during the tracking period that fledgling locations were associated with large landscape features that would assist them with return navigation to their natal area (see Mitchell et al. 2010): locations looked and in many cases were identical to adult breeding territories scattered throughout the study site. Cyprus Wheatears fledge from June onwards, but do not leave Cyprus on migration until late October (Xenophontos et al. 2017; and this also applies to juveniles, unpublished data). Therefore, it is possible that Cyprus Wheatear fledglings moved to identify migration homing targets at a larger scale after the tracking period finished.
Our results suggest that movement behaviour immediately post-fledging in a long-distance migrant passerine could function to facilitate location of breeding territories so leading to potentially earlier and more successful breeding the following year. But the pattern may have arisen by other processes. Our results could be, for example, entirely generated by random process of dispersal where individual fledglings have a random direction, distance and duration of dispersal, followed by feeding in a relatively small area up until migration. However, a key part of the future breeding territory location hypothesis is dispersal to potential future suitable breeding sites, and we show this was occurring within the first 2 months after fledging and before first long distance migration. Further research is now required to investigate if first-year birds return to breed in the post-fledging home range they established the previous summer, and then whether this leads to fitness benefits relative to any individuals that do not.
References
Aebischer A, Perrin N, Krieg M, Studer J, Meyer DR (1996) The role of territory choice, mate choice and arrival date on breeding success in the Savi’s warbler Locustella luscinioides. J Avian Biol 27:143–152
Anders AD, Dearborn DC, Faaborg J, Thompson FR (1997) Juvenile survival in a population of neotropical migrant birds. Conserv Biol 11:698–707
Anders AD, Faaborg J, Thompson FR III (1998) Postfledging dispersal, habitat use, and home-range size of juvenile Wood Thrushes. Auk 115:349–358
Arlt D, Part T (2008) Post-breeding information gathering and breeding territory shifts in northern wheatears. J Anim Ecol 77:211–219
Ausprey IJ, Rodewald AD (2011) Postfledging survivorship and habitat selection across a rural-to-urban landscape gradient. Auk 128:293–302
Berkeley LI, McCarty JP, Wolfenbarger LL, Berkeley LI, McCarty JP, Wolfenbarger LL (2007) Postfledging survival and movement in Dickcissels (Spiza americana): implications for habitat management and conservation. Auk 124:396–409
Betts MG, Hadley AS, Rodenhouse N, Nocera JJ (2008) Social information trumps vegetation structure in breeding-site selection by a migrant songbird. Proc R Soc Lond B Biol Sci 275:2257–2263
Bombieri G, Fasciolo A, Penteriani V, Illera JC, Chamberlain D, Delgado MD (2018) Disentangling the effects of genetic and environmental factors on movement behaviour. Ethology 124:139–148
Cox WA, Thompson FR, Cox AS, Faaborg J (2014) Post-fledging survival in passerine birds and the value of post-fledging studies to conservation. J Wildl Manag 78:183–193
Currie D, Thompson DBA, Burke T (2000) Patterns of territory settlement and consequences for breeding success in the northern Wheatear Oenanthe oenanthe. Ibis 142:389–398
Eng ML, Stutchbury BJM, Burke DM, Elliott KA (2011) Influence of forest management on pre- and post-fledging productivity of a neotropical migratory songbird in a highly fragmented landscape. Can J For Res Rev Can Rech For 41:2009–2019
Flint PR, Stewart PF (1992) The birds of Cyprus. British Ornithologists Union, Tring
Grüebler MU, Korner-Nievergelt F, Naef-Daenzer B (2014) Equal nonbreeding period survival in adults and juveniles of a long-distant migrant bird. Ecol Evol 4:756–765
Kershner EL, Walk JW, Warner RE (2004a) Postfledging movements and survival of juvenile eastern meadowlarks (Sturnella magna) in Illinois. Auk 121:1146–1154
Kershner EL, Walk JW, Warner RE (2004b) Postfledging movements and survival of juvenile eastern meadowlarks (Sturnella magna) in Illinois. Auk 121:1146–1154
King DI, Degraaf RM, Smith ML, Buonaccorsi JP (2006) Habitat selection and habitat-specific survival of fledgling ovenbirds (Seiurus aurocapilla). J Zool 269:414–421
Kokko H (1999) Competition for early arrival in migratory birds. J Anim Ecol 68:940–950
Lombardo MP (1987) Attendants at tree swallow nests. II. The exploratory-dispersal hypothesis. Condor 89:138–149
Maness TJ, Anderson DJ (2013) Predictors of juvenile survival in birds. Ornithol Monogr 78:1–55
Mitchell GW, Taylor PD, Warkentin IG (2010) Assessing the function of broad-scale movements made by juvenile songbirds prior to migration. Condor 112:644–654
Moore LC, Stutchbury BJM, Burke DM, Elliott KA (2010) Effects of forest management on postfledging survival of rose-breasted grosbeaks (Pheucticus ludovicianus). Auk 127:185–194
Morrison CA, Alves JA, Gunnarsson TG, Prisson B, Gill JA (2019) Why do earlier-arriving migratory birds have better breeding success? Ecol Evol 9:8856–8864
Morton ML (1991) Postfledging dispersal of green-tailed towhees to a subalpine meadow. Condor 93:466–468
Morton ML (1992) Effects of sex and birth date on premigration biology, migration schedules, return rates and natal dispersal in the mountain white-crowned sparrow. Condor 94:117–133
Naef-Daenzer B, Gruebler MU (2016) Post-fledging survival of altricial birds: ecological determinants and adaptation. J Field Ornithol 87:227–250
Newton I (2008) The migration ecology of birds. Academic Press, Oxford
Nocera JJ, Forbes GJ, Giraldeau LA (2006) Inadvertent social information in breeding site selection of natal dispersing birds. Proc R Soc Lond B Biol Sci 273:349–355
Pedersen D, Thorup K, Sunde P, Jacobsen LB, Rahbek C (2013) Post-fledging behaviour of juveniles in the little owl (Athene noctua). Ornis Fenn 90:117–128
Randler C, Teichmann C, Pentzold S (2010) Breeding habitat preference and foraging of the Cyprus Wheatear Oenanthe cypriaca and niche partitioning in comparison with migrant Oenanthe species on Cyprus. J Ornithol 151:113–121
Randler C, Foerschler MI, Gonzalez J, Aliabadian M, Bairlein F, Wink M (2012) Phylogeography, pre-zygotic isolation and taxonomic status in the endemic Cyprus Wheatear Oenanthe cypriaca. J Ornithol 153:303–312
Rappole JH, Ballard K (1987) Postbreeding movements of selected species of birds in Athens, Georgia. Wilson Bull 99:475–480
Rappole JH, Tipton AR (1991) New harness design for attachment of radio transmitters to small passerines. J Field Ornithol 62:335–337
Raybuck DW, Larkin JL, Stoleson SH, Boves TJ (2020) Radio-tracking reveals insight into survival and dynamic habitat selection of fledgling Cerulean Warblers. Condor 122:1–15
Streby HM, Andersen DE (2012) Movement and cover-type selection by fledgling ovenbirds (Seiurus aurocapilla) after independence from adult care. Wilson J Ornithol 124:620–625
Streby HM, Peterson SM, Andersen DE (2011) Invertebrate availability and vegetation characteristics explain use of nonnesting cover types by mature-forest songbirds during the postfledging period. J Field Ornithol 82:406–414
Streby HM, McAllister TL, Kramer GR, Peterson SM, Lehman JA, Andersen DE (2015) Minimizing marker mass and handling time when attaching radio-transmitters and geolocators to small songbirds. Condor 117:249–255
Sutherland GD, Harestad AS, Price K, Lertzman KP (2000) Scaling of natal dispersal distances in terrestrial birds and mammals. Conserv Ecol 4(1):16
Vega Rivera JH, McShea WJ, Rappole JH, Hass CA (1998) Postbreeding movements and habitat use of adult wood thrushes in northern Virginia. Auk 116:458–466
Velmala W, Helle S, Ahola MP, Klaassen M, Lehikoinen E, Rainio K, Sirkia PM, Laaksonen T (2015) Natural selection for earlier male arrival to breeding grounds through direct and indirect effects in a migratory songbird. Ecol Evol 5:1205–1213
Vitz AC, Rodewald AD (2007) Vegetative and fruit resources as determinants of habitat use by mature-forest birds during the postbreeding period. Auk 124:494–494
Vitz AC, Rodewald AD (2010) Movements of fledgling ovenbirds (Seiurus aurocapilla) and worm-eating warblers (Helmitheros vermivorum) within and beyond the natal home range. Auk 127:364–371
Wiltschko W, Wiltschko R (1978) Theoretical model for migratory orientation and homing in birds. Oikos 30:177–187
Xenophontos M, Cresswell W (2016a) Reproductive success and productivity of the Cyprus Wheatear Oenanthe cypriaca, a migratory, island endemic. J Ornith 157:721–731
Xenophontos M, Cresswell W (2016b) Survival and dispersal of the Cyprus wheatear Oenanthe cypriaca, an endemic migrant. J Ornithol 157:707–719
Xenophontos M, Blackburn E, Cresswell W (2017) Cyprus wheatears Oenanthe cypriaca likely reach sub-Saharan African wintering grounds in a single migratory flight. J Avian Biol 48:529–535
Yoerg SI (1998) Foraging behavior predicts age at independence in juvenile Eurasian dippers (Cinclus cinclus). Behav Ecol 9:471–477
Acknowledgements
We would like to thank Tasso Leventis who funded the tags used in this study and who provided logistical support, and Alex Kirschel for logistical support. We thank three anonymous referees for their comments that substantially improved the paper.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by F. Bairlein.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Styles, P., Patchett, R., King, J.R. et al. Movements of Cyprus Wheatear Oenanthe cypriaca fledglings: evidence of a post-fledging home range away from the natal site prior to first migration. J Ornithol 162, 339–348 (2021). https://doi.org/10.1007/s10336-020-01843-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-020-01843-z