
Abstract
Globally anthropogenic pressures on natural environments have caused species decline. Ground dwelling bird species declined in numbers because of loss of habitat and nesting sites. Protected areas (PAs) are generally perceived as areas with little disturbance; however, many have/are also subjected to anthropogenic land use change, such as agroforestry. Consequently, we investigated the presence and habitat use of a terrestrial avian species, the Crested Guinea-fowl (Guttera edouardi), in iSimangaliso Wetland Park, South Africa, a PA with varying land use change. We used single-season camera trap data from 99 trap stations across 24 days, and single season occupancy models to quantify the influence of habitat conversion and habitat characteristics on occupancy and detection probability of Crested Guinea-fowl. The average site occupancy (±standard error) and detection probability (±standard error) under the variable influence was 0.40 ± 0.07 and 0.43 ± 0.06 respectively from the top models. The most influential variables were natural forests, shrub cover, leaf litter, weed dominance, distance to water and narrow path. The natural forests, weed dominance and shrub cover positively influenced the occupancy of Crested Guinea-fowl. Leaf litter and distance to water had a positive influence on detection probability while narrow path had a negative influence. Our findings highlight the importance of natural forests to the survival of Crested Guinea-fowl in landscapes modified for agroforestry. Management initiatives are mandatory to ensure protection of adequate natural habitat for the conservation of forest specialists.
Zusammenfassung
Reaktion des Kräuselhauben-Perlhuhn ( Guttera edouardi ), eines Waldspezialisten, auf die räumliche Variation in der Landnutzung im iSimangaliso Feuchtgebiet in Südafrika Anthropogene Belastungen natürlicher Lebensräume haben weltweit zu einem Artenrückgang geführt. Bodenbewohnende Vogelarten sind aufgrund von Habitat- und Neststandortrückgang unter diesen zahlenmäßig rückläufigen Arten. Schutzgebiete gelten generell als Gebiete mit geringer Störung, aber viele von ihnen sind anthropogenen Veränderungen in der Landnutzung, wie etwa dem Waldfeldbau, ausgesetzt. Wir untersuchten deshalb die Präsenz und Habitatnutzung einer bodenbewohnenden Vogelart, dem Kräuselhaubenperlhuhn (Guttera edouardi), im iSimangaliso Feuchtgebiet in Südafrika, einem Schutzgebiet mit unterschiedlicher Veränderung in der Landnutzung. Um den Einfluss von Habitatumwandlung und Habitatcharakteristika auf die Besetzung und Nachweiswahrscheinlichkeit des Kräuselhauben-Perlhuhns zu bestimmen, verwendeten wir Daten von 99 Kamerafallen über 24 Tage einer Saison und Modelle zur Besetzung in derselben Saison. Die mittlere Besetzungsrate (±Standardfehler) und Nachweiswahrscheinlichkeit (± Standardfehler) lag unter dem Einfluss der Variablen bei 0,40 ± 0,07 und 0,43 ± 0,06 des jeweils besten Modells. Die Variablen mit dem größten Einfluss waren natürlicher Wald, Buschbedeckung, Laubstreu, Dominanz an Kräutern, Entfernung zum Wasser und die Enge des Pfades. Natürlicher Wald, Dominanz an Kräutern und Buschbedeckung hatte einen positiven Einfluss auf die Besetzungsrate mit Kräuselhauben-Perlhühnern. Laubstreu und Entfernung zum Wasser beeinflusste die Nachweiswahrscheinlichkeit positiv, während die Enge des Pfades einen negativen Einfluss hatte. Unsere Ergebnisse unterstreichen die Bedeutung von natürlichem Wald für das Überleben des Kräuselhauben-Perlhuhns in einer von Waldfeldbau modifizierten Landschaft. Managementinitiativen, die den Schutz angemessener Flächen natürlichen Habitats gewährleisten, sind für die Erhaltung von Waldspezialisten unerlässlich.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Habitat loss is one of the most significant threats to the survival of many species, especially terrestrial species (Pimm and Raven 2000; Munday 2004). Many natural landscapes across the world have been converted for various practices such as agriculture and livestock farming (Carrara et al. 2015; Millan et al. 2015). Land use changes are one of the main causes of population declines and changes in distribution of many bird species (Chamberlain and Fuller 2001; Watson et al. 2004; Ramesh and Downs 2014). With increasing changes in natural landscapes, many forest dependent bird species appear more vulnerable to extinction (Bennun et al. 1996) and habitat conversion due to the diminishing areas they use for roosting and nesting (Malan 1998; Barnes 2000; Maphisa et al. 2009). It is, therefore, necessary for effective management of natural habitats to have an improved understanding of the anthropogenic effects on natural habitats and their biodiversity, particularly on bird species (Brown et al. 2014).
In recent decades, one major change in South Africa and other parts of the world has been the replacement of natural landscapes with agroforestry or plantations of Eucalyptus spp. and Pinus spp. for commercial purposes (Allan et al. 1997; Styring et al. 2011; Lee and Carroll 2014). Globally, many studies have focused on the impacts of habitat loss on birds, but few studies have focused on the impacts of agroforestry on bird species (Chamberlain and Fuller 2001; Aratrakorn et al. 2006; Ramesh and Downs 2014). Many terrestrial bird species have become locally extinct because of the conversion of indigenous to plantations/agroforestry (Armstrong and Van Hensbergen 1996; Jansen et al. 1999; Naoe et al. 2012). Generally commercial plantations have a negative effect on bird species occurrence as the carrying capacity of the landscape decreases with range fragmentation and contraction (Neuschulz et al. 2011; Watson et al. 2004).
The biodiversity rich iSimangaliso Wetland Park (IWP), South Africa, is an area that, before a large portion was converted into plantations, was dominated by indigenous dry forest thickets, dune forest, lowland forests, and grasslands (Nustad 2011; Porter 2013). In recent years, most plantations were removed and restored to natural habitats (Porter 2013). There was agroforestry before the plantations were removed. However, some areas within IWP are still used for commercial plantations, and the impacts of these plantations on wildlife are not well documented. The Crested Guinea-fowl (Guttera edouardi) is a terrestrial bird that has declined nationally in recent years because of habitat fragmentation and commercial afforestation (Lawes et al. 2006). The Crested Guinea-fowls are terrestrial, forest specialists; however, they are sometimes found on forest edges (Barnes 2000). There is a paucity of literature on this bird species relative to other game birds worldwide (Van Niekerk 2015), and even less data on their abundance and distribution patterns with habitat availability. The conservation status of Crested Guinea-fowl in the International Union for Conservation of Nature (IUCN) is listed as least concern (BirdLife International 2015). Often, least concern species such as the Crested and Helmeted Guinea-fowl (Numida meleagris) receive little attention from ecologists (Malan 1998). However, the importance of conserving and monitoring even the most common species is necessary, as they play a crucial role in structuring the ecosystems (Gaston and Fuller 2008).
To ensure a reduction in the current biodiversity loss, it is important that research is conducted to better understand the structural and compositional changes in the different land forms, and the effects these changes have on the abundance and distribution of indigenous forest species (Sutherland et al. 2006; Radford and Bennett 2007). When conducting population monitoring studies, the presence/absence of species aids in estimating the proportion of area occupied (Ramesh and Downs 2014). Ignoring imperfect detection yields an inappropriate image of the species’ actual distribution because false absences (when the species is present, but escapes detection) are confounded with true absence. This can lead to inappropriate management decisions, making it necessary to accommodate imperfect detection in the analysis of presence/absence data. In addition, detection probability can be used as a proxy for the local relative abundance of the species (Royle and Nichols 2003; Péron and Altwegg 2015), making it possible to extract more than just presence information from the data. For this study, we used the occupancy modelling framework of Mackenzie et al. (2002) to estimate the site occupancy and detection probabilities of Crested Guinea-fowl. In cases where detection probabilities are ignored false estimates of the species could in turn affect management decisions (Mackenzie 2005; Ramesh and Downs 2014). Our main objective was to investigate the influence of habitat conversion and habitat characteristics on the occupancy and distribution of Crested Guinea-fowl. We hypothesized that plantations have a negative effect on the occupancy of Crested Guinea-fowl. We predicted that the Crested Guinea-fowls are associated primarily with natural forests and forest edges in iSimangaliso Wetland Park (IWP).
Materials and methods
Study site
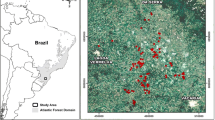
The study was conducted in the Western Shores (ca. 38,000 ha) and Eastern Shores (ca. 30,000 ha) of iSimangaliso Wetland Park (Latitude: −27.364387 to −28.445570; Longitude: 32.202902 to 32.756160) (Fig. 1), KwaZulu-Natal (KZN), South Africa. The study was conducted during a dry season (i.e., winter; July–August). The study site is situated in the northern part of KZN and has a broad habitat range, consequently supporting wildlife (Ramesh et al. 2016). Largely the Western Shores was converted to commercial plantations, Eucalyptus spp. and slash pine (Pinus elliottii) (Porter 2013) which is gradually being removed. The Western Shores also has open grasslands, dry forest thickets, sand forest and coastal lowland forest. In contrast, the Eastern Shores had less plantation cover and a few areas are recovering back to grasslands and native forest. It comprises open grasslands, coastal lowland forest and native dune forests especially along the eastern coast (Ramesh et al. 2016). The area has a subtropical climate with an average rainfall of 700 mm for the Western Shores and 1500 mm in the Eastern Shores (Leslie and Spotila 2001).

(from Ramesh et al. 2016)
Map of iSimangaliso Wetland Park, KwaZulu-Natal, South Africa with 99 camera trap station locations
Data collection and analysis
We conducted the study using passive infrared camera traps (Moultre M880, Trailcampro LLC, Springfield, MO, USA) to photograph various animal species (Ramesh et al. 2016). Crested guinea-fowl pictures were a by-catch of a study geared towards mammals by Ramesh et al. (2016). Systematic grids of 2 km2 covering 99 camera sites were used for photographing wildlife, and in every 2 km2 grid we had one camera station located at random in the grid cell. The average distance between cameras was 1.7 km, and these covered diverse habitat types including disturbed areas such as plantations. We wired the cameras on shrubs/trees at a height of 30 cm above the ground. To reduce the chances of having blank shots we removed a portion of vegetation in front of the camera to cover a wide range when taking a picture, and we ensured that the cameras were working. Multiple photographic detections of Crested Guinea-fowls in 24 h per camera site were considered as an independent photo. Since Crested Guinea-fowls were not common in the grasslands, we used data from camera-traps stationed in the non-open grassland habitat from Ramesh et al. (2016).
We considered the general maximum distance moved by ground-dwelling birds (e.g., red-necked spur-fowl Pternistis afer) to be about 1 km or less (Van Niekerk et al. 2009). We used ArcGIS 9.3.1 (ESRI, Redlands, CA, USA) to calculate the broad scale variables: percent area of natural forests, cleared plantation, and plantations by laying a 1 km buffer around each camera site using the 2008 KwaZulu-Natal land cover map (Ezemvelo KZN Wildlife 2011). Additionally, we extracted the broad scale variables: percent area of natural forests, cleared plantation, and plantations around 500 and 100 m buffer to determine if changing the radius scale around a camera site yield different results compared to the 1 km buffer extraction. Local scale variables: distance to water from the camera site (log transformed), percentage of leaf litter cover (within 5 m radius) were recorded in the field at every camera site, and plantation tree dominance, weed dominance (if the area is dominated/occupied by higher percentage of weeds than the native vegetation) were recorded as categorical variables and number of trees within 15 m of each camera trap. We also measured other local scale variables such as the percentage of canopy cover and percentage of shrub cover within 15 m radius of each camera trap. Leopards (Panthera pardus) were considered a survey covariate to assess the risk of predation on guinea-fowls in the analyses. Camera traps placed on park management roads were considered as a wider path, whereas those on animal paths/trails were considered as a narrow path. To increase the detection probability the data from 4 days were summed up into a single sampling occasion giving us six sampling occasions. For each occasion the target species could score a 1 or a 0, where 1 indicated that the animal was captured at the sampling station during that trapping occasion (at least one of the 4 days) and 0 if it was not captured. This matrix was developed for Crested Guinea-fowls and leopard. Eventually, presence/absence of leopard and wider/narrow path were covariates to model the detection probability of guinea-fowls. We modelled the occupancy and detection probability of Crested Guinea-fowl using single season occupancy models in program PRESENCE 7.9 as a function of site and survey specific covariates (Hines 2006; Mackenzie et al. 2006).
We constructed a global model with all covariates first. This was then followed by models with each covariate run separately, and later with various combinations based on their model weights and the strength of the untransformed coefficient estimates. The two-step procedure started with modelling occupancy first, then detection probability under the influence of each covariate, and later with various combinations of covariates. Lastly, we constructed models for occupancy and detection probability together. To assess the fit of the models to our data, we ran 10,000 bootstrap samples (ĉ > 1.0 indicated over-dispersion of data), and performed Pearson goodness-of fit tests (White and Burnham 1999). The models were compared using the Akaike information criterion (AIC) and all the models with delta AIC values less than two were declared to be the best models (Burnham and Anderson 2002). To check the strength of these variables on detection probability and occupancy, we calculated the Akaike weights (ω i).
Results
A total of 134 photographs of Crested Guinea-fowl were recorded from 2832 camera trap nights in IWP. There were 116 independent photographs of Crested Guinea-fowl from the Western Shores and 18 on the Eastern Shores. The Crested Guinea-fowl group size ranged from 1 to 18 and the mean group size was four individuals in both the Western and Eastern Shores.
The proportion of camera traps that yielded at least one picture of guinea-fowl (naïve occupancy) was 0.37. We observed a slight difference between the estimated and naïve occupancy. Models under the influence of variables, estimated an average site occupancy and detection probability of 0.40 ± 0.07 and 0.43 ± 0.06, respectively, from the top models. Out of all the variables measured, we found three variables each consistently associated with the occupancy (natural forest, shrub cover, weed dominance) and detection probability (leaf litter, narrow path, and distance to water) of Crested Guinea-fowl across land cover scales 1 km, 500 and 100 m from the top models (Tables 1, 2). Among all the models, only two top models (∆AIC ≤ 2) substantially contributed to species occurrence consistently at each land cover scale analysis (Tables 1, 2). The top models [psi (nf + sc), p (leafl + npath + dt2w; psi (nf + w), p (npath + dt2w)] had the maximum model AIC weight, suggesting that natural forest, shrub cover and weed dominance influenced the occupancy of Crested Guinea-fowl positively. Leaf litter and distance to water influenced the detection probability positively, while narrow path had a negative influence on the detection probability. The variable importance of the overall summed model AIC weights for each variable with respect to occupancy was high for natural forest (ω i = 1), followed by shrub cover (ω i = 0.48–0.66), weed dominance (ω i = 0.25–0.36), leaf litter (ω i = 0.27–0.66), and other variables (≤0.09). The overall summed AIC model weights for each variable with respect to detection was high for narrow path (ω i = 1) and distance to water (ω i = 0.98–1.0), followed by leaf litter (ω i = 0.47–0.53) and other variables (ω i = ≤0.02). Predicted occupancy of Crested Guinea-fowl and natural forest had a strong positive correlation (R 2 = ≥0.92; P < 0.001; Fig. 2), whereas occupancy and shrub cover had a weak correlation (R 2 = 0.24–0.31; P < 0.01; Fig. 3). Detection probability and leaf litter had a weak correlation (R 2 = 0.21; P = 0.036), whereas detection probability and distance to water had a strong positive correlation (R 2 = 0.70; P < 0.001). Furthermore, weed dominance had a positive effect on the occupancy of Crested Guinea-fowl (Fig. 4). However, the combined effect of variables was strong in the top models determining occupancy and detection probability of Crested Guinea-fowl.

Based on the covariates in the top models (ΔAIC ≤ 2); site occupancy of Crested Guinea-fowl modelled in relation to percent area of natural forest extracted at a 1 km, b 500 m and c 100 m radius around every camera site

Based on the covariates in the top models (ΔAIC ≤ 2); site occupancy of Crested Guinea-fowl modelled in relation to percent of shrub cover when broad scale variable such as the percent area of natural forest were extracted at a 1 km, b 500 m and c 100 m radius around every camera site

Based on the covariates in the top models (ΔAIC ≤ 2); site occupancy of Crested Guinea-fowl modelled in relation to weed dominance when broad scale variable such as the percent area of natural forest were extracted at 1 km, 500 and 100 m radius around every camera site
Discussion
Our study investigated the effect of land use on the occupancy and distribution of the Crested Guinea-fowl in IWP during the winter season. Variables at broad scale (percent area of natural forest) and local scale (shrub cover, weed dominance, leaf litter, narrow path, and distance to water) influenced occurrence of Guinea-fowl. The site occupancy modelling clearly showed that indigenous forests play an important role in predicting Crested Guinea-fowl occupancy. Natural forests had the highest summed model weight, showing that the occupancy of Crested Guinea-fowl is mostly dependent on this variable. Therefore, it is very important that appropriate conservation measures are taken in order to ensure that this species does not lose its natural forest habitat. Also. no direct effect of plantations on the occupancy of Crested Guinea-fowl was recorded; there is a likely indirect effect via the decrease in indigenous forest cover where there are plantations. Native habitats are important for the survival of many terrestrial species, particularly for specialist species (Bennun et al. 1996; Ramesh et al. 2016). Ramesh et al. (2016) highlighted the need to reduce the rate of commercial afforestation, particularly in protected areas, since these areas play a significant role in preserving biodiversity and species richness. The plantations in IWP are assumed to be the main reason for the low estimated site occupancy. In comparison to other similar studies, Allan et al. (1997) discussed the negative impacts that commercial plantations have on the biodiversity as a whole, thus showing the importance of native plants especially on the survival of ground-dwelling birds that are forest specialists. On the contrary, some studies have found that commercial plantations favour and increase the distribution ranges of some bird species (Brockerhoff et al. 2008; Loyn et al. 2007). Ramesh and Downs (2014) found that in the absence of indigenous forest, commercial plantations positively influenced the site occupancy of red-necked spur-fowl, but also discussed their susceptibility to detrimental effects of changing land-use.
Shrub cover has been observed as an important factor in the survival of many avian populations. This factor is mostly important in providing nesting sites (Donnelly and Marzluff 2004). In this study, shrub cover was an important variable and positively influenced the site occupancy of Crested Guinea-fowl. It was assumed that the Crested Guinea-fowl also uses shrub areas for nesting and roosting. Several studies have discussed that areas with native forests have more insect groups than areas with commercial plantations (Helle and Muona 1985; Vetter et al. 2011). For insectivorous bird species, the presence of invertebrates in forests is essential for their survival since it is their primary source of food (Helle and Muona 1985; Vetter et al. 2011). Invertebrates use the leaf litter from the indigenous trees to hide from predators (Coolen et al. 2005), which supports our findings that leaf litter positively influenced the detection of Crested Guinea-fowl. Another important factor in driving the distribution and detection of many species is predation risk; higher predation risks limit distribution and vice versa (Watts 1991). The detection probability of Crested Guinea-fowl was found to increase as the distance from the water-points increased. Animals using water-points are generally susceptible to high predation risk (Valeix et al. 2008). Therefore, it was assumed that the Crested Guinea-fowl avoided the areas near water points in order to reduce the risk of predation. Also, competition with other game-birds near water points might have influenced the negative relationship between distance to water and detection probability. Moreover, another possible reason for the observed relationship between distance to water and detection probability is that the Crested Guinea-fowls are usually abundant in dry habitats (BirdLife International 2015). Furthermore, narrow path negatively influenced the detection of Crested Guinea-fowl in the study sites. Usually, narrow paths especially in forests, are used by small sized species in an effort to avoid larger predators such as leopards. However, contrary to our prediction, our results showed that Crested Guinea-fowls avoided narrow paths and used wider paths, possibly indicating that there may be less predation pressure on the wider paths by larger predators. The effect of narrow path possible these paths are preferred by mesopredators in areas with abundant large predators (Ramesh et al. 2016). In addition, we presumed that the Crested Guinea-fowl used the wider paths because of the increased probability of finding food (i.e., seeds) in wider paths compared to narrow paths.
In conclusion, habitat conversion had significant effects on the distribution of Crested Guinea-fowl. Appropriate conservation measures are needed in order to ensure that the current native dune forests in IWP are protected from human induced modifications. This study highlighted the most important variables for the distribution and occupancy of Crested Guinea-fowl and found that natural forests were the most important variable for occupancy. Although the study focused on the impacts of habitat conversion and habitat characteristics on occupancy and distribution of a single species, the findings can be helpful on the conservation of other terrestrial bird species, particularly those who solely depend on indigenous forests to survive. Moreover, the sampling method used in this study can be applied when investigating the occupancy and distribution of many other species with specific niches (Ramesh and Downs 2014). For our study we ran a single season model which gave us the results of occupancy and detection probability for only one season (i.e., winter). We recommend that future studies run a multi-season model in order to determine if the occupancy and distribution changes between the four seasons (i.e., spring, summer, autumn, winter), or during the breeding and non-breeding seasons. Moreover, it is recommended that the impacts of land-use change on ground-dwelling birds be thoroughly investigated, as well as the degree to which factors like predation have on the distribution of species. Finally, future studies must also investigate the patch size of indigenous forests required by ground-dwelling birds, particularly the forest dependent species.
References
Allan DG, Harrison JA, Navarro R, Van Wilgen BW, Thompson MW (1997) The impact of commercial afforestation on bird populations in Mpumalanga Province, South Africa—insights from bird-atlas data. Biol Conserv 79:173–185
Aratrakorn S, Thunhikorn S, Donald PF (2006) Changes in bird communities following conversion of lowland forest to oil palm and rubber plantations in southern Thailand. Bird Conserv Int 16:71–82
Armstrong AJ, Van Hensbergen HJ (1996) Impacts of afforestation with pines on assemblages of native biota in South Africa. S Afr J For 175:35–42
Barnes KN (2000) The Eskom red data book of birds of South Africa, Lesotho and Swaziland. Bird Life South Africa, Johannesburg
Bennun L, Dranzoa C, Pomeroy D (1996) The forest birds of Kenya and Uganda. J East Afr Nat yHist 85:23–48
BirdLife International (2015) Guttera edouardi. http://www.birdlife.org. Accessed 10 Oct 2015
Brockerhoff EG, Jactel H, Parrotta JA, Quine CP, Sayer J (2008) Plantation forests and biodiversity: oxymoron or opportunity? Biodivers Conserv 17:925–951
Brown ML, Donovan TM, Schwenk WS, Theobald DM (2014) Predicting impacts of future human population growth and development on occupancy rates of forest-dependent birds. Biodivers Conserv 170:311–320
Burnham KP, Anderson DR (2002) Model selection and multi model inference: a practical information-theoretic approach. Springer, New York
Carrara E, Arroyo-Rodríguez V, Vega-Rivera JH, Schondube JE, de Freitas SM, Fahrig L (2015) Impact of landscape composition and configuration on forest specialist and generalist bird species in the fragmented Lacandona rainforest, Mexico. Biodivers Conserv 184:117–126
Chamberlain DE, Fuller RJ (2001) Contrasting patterns of change in the distribution and abundance of farmland birds in relation to farming system in lowland Britain. Glob Ecol Biogeogr 10:399–409
Coolen I, Dangles O, Casas J (2005) Social learning in non-colonial insects? Curr Biol 15:1931–1935
Donnelly R, Marzluff JM (2004) Importance of reserve size and landscape context to urban bird conservation. Conserv Biol 18:733–745
Ezemvelo KZN Wildlife (2011) 2008 KZN Province Land-Cover Mapping (from SPOT5 Satellite imagery circa 2008). Prepared for Ezemvelo KZN Wildlife (Biodiversity Research), South Africa
Gaston KJ, Fuller RA (2008) Commonness, population depletion and conservation biology. Trends Ecol Evol 23:14–19
Helle P, Muona J (1985) Invertebrate numbers in edges between clear-fellings and mature forests in northern Finland. Silva Fenn 19:281–294
Hines JE (2006) PRESENCE-Software to estimate patch occupancy and related parameters: USGS-PWRC. http://www.mbr-pwrc.usgs.gov/software/presence.html. Accessed 3 April 2015
Jansen R, Little RM, Crowe TM (1999) Implications of grazing and burning of grasslands on the sustainable use of francolins (Francolinus spp.) and on overall bird conservation in the highlands of Mpumalanga province, South Africa. Biodivers Conserv 8:587–602
Lawes MJ, Fly S, Piper SE (2006) Gamebird vulnerability to forest fragmentation: patch occupancy of the Crested Guinea-fowl (Guttera edouardi) in Afromontane forests. Anim Conserv 9:67–74
Lee MB, Carroll JP (2014) Relative importance of local and landscape variables on site occupancy by avian species in a pine forest, urban, and agriculture matrix. For Ecol Manag 320:161–170
Leslie AJ, Spotila JR (2001) Alien plant threatens Nile crocodile (Crocodylus niloticus) breeding in Lake St. Lucia, South Africa. Biol Conserv 98:347–355
Loyn RH, McNabb EG, Macak P, Noble P (2007) Eucalypt plantations as habitat for birds on previously cleared farmland in south-eastern Australia. Biol Conserv 137:533–548
Mackenzie DI (2005) What are the issues with presence-absence data for wildlife managers? J Wildl Manag 69:849–860
MacKenzie DI, Nichols JD, Lachman GB, Droege S, Royle JA, Langtimm CA (2002) Estimating site occupancy rates when detection probabilities are less than one. Ecology 83:2248–2255
MacKenzie DI, Nichols JD, Royle JA, Pollock KP, Bailey LL, Hines JE (2006) Occupancy estimation and modelling: inferring patterns and dynamics of species occurrence. Academic Press, New York
Malan G (1998) Summer grassland cover on cattle farms in KwaZulu-Natal: does it limit nesting habitat for helmeted guineafowl? S Afr J Wildl Res 28:105–109
Maphisa DH, Donald PF, Buchanan GM, Ryan PG (2009) Habitat use, distribution and breeding ecology of the globally threatened Rudd’s lark and Botha’s lark in eastern South Africa. Ostrich 80:19–28
Millan CH, Develey PF, Verdade LM (2015) Stand-level management practices increase occupancy by birds in exotic Eucalyptus plantations. For Ecol Manag 336:174–182
Munday PL (2004) Habitat loss, resource specialization, and extinction on coral reefs. Glob Chang Biol 10:1642–1647
Naoe S, Sakai S, Masaki T (2012) Effect of forest shape on habitat selection of birds in a plantation-dominant landscape across seasons: comparison between continuous and strip forests. J For Res 17:219–223
Neuschulz EL, Botzat A, Farwig N (2011) Effects of forest modification on bird community composition and seed removal in a heterogeneous landscape in South Africa. Oikos 120:1371–1379
Nustad KG (2011) Performing natures and land in the iSimangaliso Wetland Park, South Africa. Ethnos 76:88–108
Péron G, Altwegg R (2015) Twenty-five years of change in southern African passerine diversity: nonclimatic factors of change. Glob Chang Biol 21:3347–3355
Pimm SL, Raven P (2000) Biodiversity: extinction by numbers. Nature 403:843–845
Porter RN (2013) South Africa’s first World Heritage Site. In: Perissinotto R, Stretch DD, Taylor RH (eds) Ecology and conservation of estuarine ecosystems: Lake St Lucia as a global model. Cambridge University Press, Cambridge, pp 1–19
Radford JQ, Bennett AF (2007) The relative importance of landscape properties for woodland birds in agricultural environments. J Appl Ecol 44:737–747
Ramesh T, Downs CT (2014) Land use factors determining occurrence of Red-necked Spurfowl (Pternistis afer) in the Drakensberg Midlands, South Africa. J Ornithol 155:471–480
Ramesh T, Kalle R, Rosenlund H, Downs CT (2016) Native habitat and protected area size matters: Preserving mammalian assemblages in the Maputaland Conservation Unit of South Africa. For Ecol Manag 360:20–29
Royle JA, Nichols JD (2003) Estimating abundance from repeated presence-absence data or point counts. Ecology 84:777–790
Styring AR, Ragai R, Unggang J, Stuebing R, Hosner PA, Sheldon FH (2011) Bird community assembly in Bornean industrial tree plantations: effects of forest age and structure. For Ecol Manag 261:531–544
Sutherland WJ, Armstrong-Brown S, Armsworth PR, Tom B, Brickland J, Campbell CD, Watkinson AR (2006) The identification of 100 ecological questions of high policy relevance in the UK. J Appl Ecol 43:617–627
Valeix M, Fritz H, Matsika R, Matsvimbo F, Madzikanda H (2008) The role of water abundance, thermoregulation, perceived predation risk and interference competition in water access by African herbivores. Afr J Ecol 46:402–410
Van Niekerk JH (2015) Vocal behaviour of crested guinea-fowl (Guttera edouardi) based on visual and sound playback surveys in the Umhlanga Lagoon Nature Reserve, KwaZulu-Natal Province, South Africa. Avian Res 6:1–8
Van Niekerk JH, Barendse M, Mare F (2009) Behaviour of Red necked spurfowl Pternistis afer in the Boknes and Cannon Rock coastal resorts, Alexandria district, Eastern Cape Province, South Africa. Ostrich 80:43–45
Vetter D, Hansbauer MM, Végvári Z, Storch I (2011) Predictors of forest fragmentation sensitivity in Neotropical vertebrates: a quantitative review. Ecography 34:1–8
Watson JE, Whittaker RJ, Dawson TP (2004) Habitat structure and proximity to forest edge affect the abundance and distribution of forest-dependent birds in tropical coastal forests of south eastern Madagascar. Biol Conserv 120:311–327
Watts BD (1991) Effects of predation risk on distribution within and between habitats in savannah sparrows. Ecology 72:1515–1519
White GC, Burnham KP (1999) Program MARK: survival estimation from populations of marked animals. Bird Study 46:120–139
Acknowledgments
We would like to thank the University of KwaZulu-Natal and the Hans Hoheisen Charitable Trust for funding the project. We are grateful to iSimangaliso Wetland Park Authority and Ezemvelo KZN Wildlife for permission to conduct research in the park. We would like to thank E. Kheswa for her assistance during the project work.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by T. Gottschalk.
Rights and permissions
About this article

Cite this article
Maseko, M.S.T., Ramesh, T., Kalle, R. et al. Response of Crested Guinea-fowl (Guttera edouardi), a forest specialist, to spatial variation in land use in iSimangaliso Wetland Park, South Africa. J Ornithol 158, 469–477 (2017). https://doi.org/10.1007/s10336-016-1406-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-016-1406-7