
Abstract
Young night-migratory birds establish a functional star compass during ontogeny by searching the starry night sky for its centre of rotation and interpreting this as “north”. The birds then use the learned location of the rotational centre to calibrate their magnetic compass. Here, we examine the required duration of this learning process. We exposed three groups of hand-raised inexperienced European Robins Erithacus rubecula to a rotating artificial star pattern for 3 weeks, 1 week, or 1 night, respectively, during ontogeny. A control group was exposed to the same, but stationary, artificial star pattern for 3 weeks. During their first autumn migration, we tested the birds’ orientation under the stationary star pattern as well as in the local geomagnetic field with no stars visible. Only the group that had the longest exposure time to celestial rotation during ontogeny was able to show orientation in the appropriate direction during autumn migration in the local geomagnetic field in relation to the memorized former centre of celestial rotation. This suggests that these birds had calibrated their magnetic compass relative to the star pattern seen during ontogeny. All other groups showed inappropriate or random orientation both under the stationary star sky and in the local geomagnetic field, suggesting that 7 nights of observing celestial rotation are not sufficient for young European Robins to establish a star compass and to calibrate their magnetic compass accordingly.
Zusammenfassung
Das Erlernen des Sternenkompasses: Wie lange brauchen Zugvögel dafür?
Junge, nachts ziehende Singvögel entwickeln einen Sternenkompass während ihrer Ontogenie, indem sie den nächtlichen Sternenhimmel beobachten, das Rotationszentrum ermitteln und dieses für sich als “Norden” interpretieren. Anschließend wird mit Hilfe dieses Wissens über die Lage des Sternen- Nordens ihr magnetischer Kompass kalibriert. In der vorliegenden Studie untersuchen wir, wie lange Vögel für diesen Lernprozess benötigen. Wir bildeten drei Gruppen von jungen, unerfahrenen, handaufgezogenen Rotkehlchen Erithacus rubecula und präsentierten ihnen für jeweils drei Wochen, eine Woche bzw. nur eine Nacht ein künstliches, rotierendes Sternenmuster. Während ihres ersten Herbstzuges testeten wir die Vögel sowohl unter dem stationären Sternenmuster als auch im lokalen Erdmagnetfeld ohne sichtbares Sternenmuster. Lediglich die Gruppe, die während der Ontogenie am längsten den rotierenden Sternenhimmel beobachten durfte, konnte eine entsprechende Orientierung im lokalen Erdmagnetfeld bezüglich des erinnerten Rotationsfixpunktes zeigen. Dies legt nahe, dass diese Vögel während ihrer Ontogenie ihren Magnetkompass bezüglich des Sternenmusters kalibriert haben. Alle anderen Gruppen zeigten weder unter dem Sternenmuster noch im lokalen Erdmagnetfeld eine angemessene Zugrichtung. Das deutet darauf hin, dass sieben Nächte Beobachtung des rotierenden Sternenhimmels während der Ontogenie nicht ausreichen den Sternenkompass zu lernen und den Magnetkompass entsprechend zu kalibrieren.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Every autumn, young birds of various passerine species migrate south towards their wintering quarters without being guided by experienced birds. To cope with this extraordinary journey, these young birds can rely on a circannual clock and different compass systems (Gwinner 1996; Berthold and Helbig 1992; Mouritsen and Mouritsen 2000; Mouritsen 2003; Liu and Chernetsov 2012). They can use the sun and associated cues (Kramer 1950; Moore 1978; Cochran et al. 2004; Muheim et al. 2006a), the magnetic field (Merkel and Wiltschko 1965; Wiltschko and Wiltschko 1972; Cochran et al. 2004) and the stars (Sauer 1957; Emlen 1967a, b) for orientation. The star compass was one of the first orientation mechanisms that was scientifically described (Sauer 1956). Although juvenile birds can use a star compass during their first migration, it is not the star patterns that are inherited. Emlen (1967a, b, 1970) showed that juvenile birds have to learn the star compass before their first migration. During their first summer, the birds observe the starry night sky and derive from it the centre of celestial rotation (Emlen 1967a, b, 1970). Due to the rotation of the Earth, the centre of the apparent celestial rotation coincides with the geographic north and south poles. In the northern hemisphere, the Polar Star is located nearly exactly at this rotational centre and, thus, indicates north.
Emlen (1967a, b, 1970) also discovered that Indigo Buntings Passerina cyanea, a North American night-migratory songbird, did not have any prior expectation about how the star patterns should look like and where the rotational centre is located. He rotated the planetarium sky around another star and the birds considered that star as their “North” Star (Emlen 1967a, b, 1970). Wiltschko et al. (1987) took this concept one step further by showing that Garden Warblers Sylvia borin, a European night-migratory songbird, can learn and use a completely artificial pattern for orientation, even when it consists of just 16 light dots, and even though there was no light dot in the centre of the “sky”. Thus, fledgling birds seem to respond to the apparent motion of the night sky and interpret the centre of rotation as poleward (north in the northern hemisphere and south in the southern hemisphere).
Once their stellar compass is fully established, birds no longer need the starry sky to rotate (e.g. Emlen 1967a, b; Wiltschko et al. 1987; Mouritsen and Larsen 2001; Liu and Chernetsov 2012), and they can orient by the stars even if only some of the stars or star patterns are visible (Emlen 1970). This means that birds subsequently learned the star constellations around the celestial centre of rotation (Emlen 1970). Furthermore, the rotational phase of the stars seems to be irrelevant (Emlen 1970; Mouritsen and Larsen 2001).
Cue conflict experiments revealed that, during ontogeny, celestial cues seem to be superior to magnetic cues (Emlen 1970; Bingman 1983, 1984; Wiltschko et al. 1987; Liu and Chernetsov 2012). But this relationship seems to change during migration, at least under some circumstances (Wiltschko and Wiltschko 1976; Beason 1987; Åkesson et al. 2002; Cochran et al. 2004; Muheim et al. 2006a, b; Chernetsov et al. 2011; Liu and Chernetsov 2012).
Able and Able (1990b) placed young Savannah Sparrows Passerculus sandwichensis during the pre-migratory phase in an experimental set-up where the birds could detect the rotational centre of an artificial star pattern in the local undisturbed geomagnetic field. These birds oriented in the appropriate direction during their first migration in autumn, even in the absence of the star patterns, when the magnetic field remained the only available orientation cue. The direction chosen was on the axis that included the former celestial centre of rotation (Able and Able 1990b). This and other studies on juvenile birds suggest that the star compass is the primary compass system during ontogeny and that the magnetic compass is calibrated accordingly (e.g. Emlen 1970; Bingman 1983; Able and Able 1990a; Prinz and Wiltschko 1992; Weindler et al. 1995; but see Alerstam and Högstedt 1983). Several studies on other passerine migrants, however, suggest that the magnetic compass system can carry a calibration on its own (Wiltschko and Gwinner 1974; Wiltschko et al. 1980; Bletz et al. 1996; Beck and Wiltschko 1982; Bingman 1981, 1983; Able and Able 1990a). Muheim et al. (2006b), and Liu and Chernetsov (2012) provide detailed reviews of the extensive literature on the interrelation of celestial and magnetic compass cues.
Despite more than five decades of research on the star compass mechanism, little has been done on the question of how birds can learn the star compass. For example, the basic question: “How long do birds have to observe the stellar rotation in order to establish their star compass?” has not yet been addressed. In previous studies on hand-raised night-migratory birds, exposure times to celestial rotation were ca. 1 month in Emlen (1967b, 1970) and Wiltschko et al. (1987), 22 nights in Able and Able (1990b), 7 weeks in Prinz and Wiltschko (1992) and 14 nights in Weindler et al. (1995). Thus, it can be concluded that 2 weeks or more is sufficient to enable birds to detect the centre of celestial rotation, to learn the star patterns and to calibrate their magnetic compass accordingly. But it remains unknown whether birds can do so within a much shorter time period.
In the present study, we investigated how many nights during the pre-migratory period young inexperienced European Robins Erithacus rubecula needed to be exposed to stellar rotation in order to orient in their proper migratory direction during autumn.
We therefore exposed groups of birds for 1, 7 or 21 nights to a rotating star pattern during their first summer. We tested the birds’ orientation under the stationary starry sky and in wooden huts where they only had the magnetic field available during their first autumn migratory season.
Methods
Subjects
Thirty-seven European Robin Erithacus rubecula nestlings between the ages of 5 and 11 days were collected from eight nests in May 2010 in the vicinity of Oldenburg (Lower Saxony, Germany). The birds were hand-raised and kept in a windowless room and, thus, prevented from seeing any celestial cues such as the sun, the stars or the polarised light pattern of the sky. Automatically controlled daylight lamps mimicked the local photoperiod. 30 min before sunrise and 30 min after sunset, weak warm-light lamps simulated dusk and dawn. All experiments were performed in accordance with EU and national guidelines for the use of animals in research and were approved by local authorities (Niedersächsisches Landesamt für Verbraucherschutz und Lebensmittelsicherheit/LAVES, Oldenburg, Germany).
Star test chambers
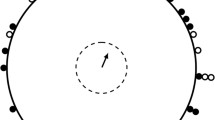
Three wooden 2 × 2 × 2 m chambers were built. The chambers were shielded outside with aluminium and were grounded in order to screen electromagnetic disturbances while maintaining the Earth’s stationary magnetic field. A black curtain covered the walls inside from the top to the floor. An artificial sky, which consisted of a 2-m-diameter wooden disk painted black, was mounted on the ceiling of each chamber. Sixteen warm-light diodes with a continuous light spectrum were embedded in each disk in eight fields of two diodes each at a distance of 55 cm from the centre. The geometric pattern of the diodes was a reconstruction from Wiltschko and Wiltschko (1976) and Wiltschko et al. (1987). The ceiling plate was rotated by a motor with a speed of 360° per 24 h. In each chamber, an additional motor was installed that could not move the disk but emitted a noise similar to the real motors. This sham motor was switched on in cases when a stationary sky was presented in order to guarantee comparable conditions. Ten Emlen funnels (Emlen and Emlen 1966) covered with nets were set up evenly spaced on the floor at a distance of 75 cm from the centre of the chamber (Fig. 1). The rotational centre of the sky disk (equivalent to “star” north) was in the middle of the chamber. Therefore, it deviated by 36° from geomagnetic north for each position. This leads to the effect that birds which are perfectly oriented relative to geomagnetic north would be absolutely randomly oriented relative to stellar north (Fig. 2a–c) and vice versa (Fig. 2d–f).

Schematic drawing of the star test chambers. European Robins Erithacus rubecula learned at ten individually fixed funnel positions. During testing, the birds’ view was limited by a black paper screen (demonstrated exemplarily). An additional light source was added in the centre. It was located in an opaque cylinder and covered with a Plexiglas plate with a black central part and thus emitted diffuse light onto the periphery of the ceiling. The star disk rotated counter clockwise (CCW) by 360° in 24 h and the rotational fix point of the disk is expected to be interpreted by the young birds as stellar north (sN). The true geographical direction of the centre of rotation of the artificial starry sky depends on the individual learning position of the bird, whereas magnetic north (mN) is in a consistent geographical direction for all funnel positions in the chamber

Theoretical predictions of bird orientation in the star test chambers depending on whether the birds would orient towards magnetic south ±5° (a–c) or towards stellar south ±5° (d–f). Left panel schematic drawing of funnel positions in the star test chambers. Small circles depict the funnel positions. One individual bird was always placed at the same funnel position in the same star test chamber during learning and testing. Arrows in (a) and (d) indicate exemplarily the ideally expected preferred jumping direction of individual birds inside the star test chamber if the birds would orient relative to magnetic north (mN) (a) or artificial stellar north (sN) (d). Note that, due to the experimental design, if the birds use a magnetic compass that is not calibrated from celestial rotation, the birds should be significantly oriented relative to mN but should be randomly oriented relative to their learned sN (b, c). If birds use a magnetic compass that is calibrated by the learned sN, they should be randomly oriented relative to mN, but significant magnetic orientation should appear after the magnetic directions are corrected for each birds’ leaned sN direction (e. f). Middle panel bird orientation relative to magnetic north; black dots represent the mean orientation of an individual bird. Right panel bird orientation relative to learned stellar north
Magnetic test huts
The magnetic orientation of birds was tested in four wooden huts. The huts were shielded with aluminium inside and were grounded to maintain an unchanged stationary geomagnetic field but to screen electromagnetic disturbances. The set-up was homogeneously illuminated by dim light bulbs with 2.1 mW/m2. Up to nine Emlen funnels covered with an opaque Plexiglas plate were set up on a wooden desk in the middle of each hut. In contrast to other tests by our group (e.g. Zapka et al. 2009; Hein et al. 2011; Engels et al. 2012), no artificially induced magnetic field was applied by Helmholtz coils around the desk so that only the natural local geomagnetic field was available for orientation.
Learning
Thirty-seven nestlings were split into four groups of ten or nine birds each with siblings distributed as evenly as possible. The birds were exposed to the artificial starry sky for learning well before migration. The first group of birds was exposed for 21 nights between 26 July and 30 August 2010 (long learning group), the second group for 7 nights between 7 August and 28 August 2010 (middle learning group) and the third group for only 1 night on 21 August 2010 (short learning group). The exact exposure dates for each group are summarised in Table 1. The fourth group served as a control group that was allowed to observe the same but stationary sky for 21 nights in parallel to the long learning group. We know that night-migratory songbirds have learned the location of the centre of celestial rotation at the start of their migratory period, but the exact sensitive period for star compass learning in European Robins within the pre-migratory period is not yet known. Generally, sensitive periods for learning and imprinting do not end abruptly but can be extended in time if animals do not have access to the critical stimulus during the normal sensitive period (Bischof 2007). We therefore chose to expose our birds to the rotating starry sky towards the end of the pre-migratory period.
For illustration purposes, the centre of the artificial starry sky is always defined as stellar north even though the birds of the control group are not expected to consider the centre of the star sky disk as stellar north. Each night, the birds were transported from their home cages to the test cages in boxes covered by a blanket to prevent the birds from seeing any celestial cues. During the whole learning phase, each individual bird kept its same funnel position in the same star test chamber. The first exposures started on 26 July 2010 at 2130 hours at local sunset and lasted until 0530 hours at local sunrise. We kept the starting and ending clock times constant throughout the learning phase to keep the duration of exposure constant between groups.
Testing
During testing, birds were brought from their home cages at local sunset in blanket-covered transport boxes to the star test chambers. The individual birds were placed at exactly the same funnel position where they had learned the stars. The birds could therefore use the artificial star configuration as well as the Earth’s magnetic field for orientation. After initial tests revealed a strong light artefact towards the centre of the hut, the star test chambers were slightly modified. We set up black paper screens of 30 cm in height around the funnels to reduce the birds’ view and to only allow them to see the starry sky disk above each funnel (Fig. 1). Also, an additional light source (LED warm white 3,100k, 1.8 W) was mounted in the centre of each chamber. The light was placed on the floor in a 20-cm-tall plastic cylinder. On top of the cylinder was an opaque Plexiglas plate on which we placed a 10-cm-diameter black cardboard circle, so that the LED primarily illuminated the periphery of the ceiling (Fig. 1). Although the light artefact remained, light conditions inside the star test chambers were more homogeneous and tests under these conditions were carried out between 16 September and 26 October 2010. The artificial star pattern was stationary for all groups during orientation testing and the Emlen funnels were equipped with scratch sensitive paper (Mouritsen et al. 2009) in order to record scratches resulting from the migratory restlessness behaviour of the birds. The overlap points of the papers were randomly chosen to be oriented towards geomagnetic north, south, east or west. Tests started half an hour after sunset and were run for 1.5 h.
After noticing a photo-tactic artefact towards the centre of the star test chambers, we also performed tests in the magnetic test huts where only the local geomagnetic field was available as an orientation cue between 17 September and 10 November 2010. For the magnetic orientation tests, the birds were brought from their home cages at sunset and placed in the Emlen funnels for 1.5 h. The test position of the individual birds inside the magnetic test huts changed from night to night. The aim of these experiments was to test if the birds had calibrated their magnetic compass according to the memorized former centre of celestial rotation derived from their individual learning positions in the star test chambers as reported by Able and Able (1990b).
The birds belonging to the long learning group stayed in the window-less housing room over the winter and in the next spring, those birds were tested again for their orientation in the magnetic test huts between 14 March and 25 April 2011.
Analyses of the scratches on the thermal papers were done double-blindly by two researchers independently from one another. When the mean direction estimates differed by more than 30°, papers were reanalysed by a third person and eventually excluded when no agreement was possible. The same procedure was applied for papers with <100 scratches (considered as non-active), random or clearly bi-modally distributed scratches. Data from birds that produced less than two oriented tests in one test condition were excluded from further analyses for this condition. This resulted in slightly different numbers of birds providing data per condition. Mean directions over all tests per individual bird were calculated for each test condition and analysed in Matlab 6.1 by a Rayleigh test for circular distributions according to Batschelet (1981).
Results
We expected that, if the birds had detected the rotational centre of the star sky disk during the learning phase in summer, they should orient away from the former centre of rotation during autumn migration. When data were analysed relative to the former rotational centre with the direction of 0° meaning “stellar north”, birds from all groups tested in the star test chambers were always significantly oriented towards “stellar north” which is equivalent to the centre of the star test chambers (Table 2). Since this orientation was also seen in the control group where the birds were exposed to a stationary sky during the pre-migratory phase, and therefore received no information at all about the rotational centre of the artificial starry sky disk and thus had only magnetic information available, the observed orientation cannot be a true star compass orientation towards stellar north but it must be a strong, probably photo-tactic artefact. The same birds’ orientation was random when their orientation was analysed relative to the geomagnetic field (Table 2).
To overcome the artefact in orientation, we also tested the birds in a homogeneous light regime (light intensity 2.1 mW/m²) in the magnetic test huts without any stars visible. There, the only available cue for orientation was the local geomagnetic field. We expected that, if the magnetic compass would not need to be calibrated from celestial rotation, all four groups of birds should orient in the appropriate SW magnetic autumn migratory direction. If celestial calibration is necessary for setting the birds’ magnetic compass, we expected that, if the birds had learned the rotational centre in the star test chambers in summer, they should have calibrated their magnetic compass relative to the observed axis of celestial rotation and, thus, should be able to orient with their magnetic compass by the memorised stars even when they are not visible. The orientations of the three treatment groups as well as the orientation of the control group were all random when analysed relative to local geomagnetic north (Table 2; Fig. 3a–d). Thus, even though our birds had access to an undisturbed magnetic field during their entire life, they had not developed a functional magnetic compass which was independent of celestial calibration.

Orientation of hand-raised European Robins in magnetic test huts in autumn 2010. From left to right long learning group (a, e), middle learning group (b, f), short learning group (c, g), and control group (d, h). Top row (a–d) orientation relative to magnetic north (mN). Bottom row (e–h) orientation relative to the birds’ potentially learned star north (sN) as derived from the individual bird’s learning position in the star test chambers. The dots at the periphery of the circle represent the mean heading of a single bird, the arrow represents the group orientation mean vector (circle radius represents a vector length r = 1); the solid lines indicate the 95 % confidence intervals; the inner and outer dashed circles indicate the required length of r for significance levels of 5 and 1 % according to the Rayleigh test, respectively
However, when the orientation of the birds of the long learning group in the magnetic test huts was analysed relative to the direction in which the birds had observed the centre of rotation, they used the magnetic field to orient highly significantly away from the memorized stellar north (Rayleigh test: n = 7, mean direction = 183°, r = 0.804, p = 0.006, 95 % confidence interval 183° ± 36°; Fig. 3). The magnetic orientation of the other three groups was not significantly magnetically directed in relation to the direction in which these birds had observed the centre of the artificial sky (stellar north, Table 1; Fig. 3e–h).
To get a confirmation that the birds of the long learning group had really calibrated their magnetic compass from the centre of rotation of the artificial starry sky, these birds were also tested in the magnetic test huts in spring 2011 without having access to any orientation cues other than the geomagnetic field during the winter. During spring migration, the birds were expected to turn their migratory direction by 180° compared to their chosen directions in autumn, i.e. towards the memorised stellar north. Indeed, the long learning group showed a strong tendency to orient in the appropriate spring migratory direction relative to the memorized stellar north (Rayleigh test: n = 7, mean direction = 26°, r = 0.600, p = 0.076, 95 % confidence interval 26° ± 58°; Fig. 4). Their orientation in relation to the local geomagnetic field was random (Table 2; Fig. 4). When the orientation of each individual bird was analysed relative to the same individuals’ orientation in the previous autumn, the birds were significantly oriented in the opposite direction compared to their orientation the previous autumn (Rayleigh test: n = 7, mean change in direction = 159°, r = 0.650, p = 0.046, the 95 % confidence interval 159° ± 52° includes the expected 180° turn in orientation).

Magnetic field orientation tests performed in the magnetic test huts during spring 2011 with hand-raised European Robins that had observed an artificial starry sky that rotated for 21 nights during their first summer in 2010. a Orientation in spring 2011 relative to magnetic north (mN). b Magnetic orientation in spring 2011 depicted relative to the magnetic direction in which each bird had observed the centre of rotation (i.e. learned star north, sN). c The springtime orientation of each bird relative to the same bird’s individual orientation during the previous autumn. The orientation of a bird during the previous autumn is defined as 0°. Thus, an orientation towards 180° would mean that the bird oriented in the exact opposite direction in spring compared to the same bird’s orientation in autumn. For descriptions of circular diagrams, see Fig. 3
Discussion
When the birds that were exposed for 21 nights to a rotating artificial starry sky were tested in magnetic test huts, where only the local geomagnetic field was available as an orientation cue, they were randomly oriented relative to the local geomagnetic north but magnetically oriented in relation to the learned stellar north derived from where they had observed the rotational centre of the sky disk in the star test chambers during the learning phase in summer. The existence of a celestially calibrated magnetic compass was confirmed by a reversal of the birds’ celestially calibrated magnetic migratory direction in spring.
The control group that was exposed to a stationary artificial starry sky, and thus had access to magnetic cues only, was randomly oriented both in relation to magnetic north and in relation to the centre of the star test chamber (depicted in the figures as stellar north, “sN”, even though the control group is not expected to consider this as “stellar north”). The groups that had observed the rotating artificial starry sky disk for 1 week or 1 night in the star test chambers in summer also remained randomly oriented in the magnetic test huts when their orientation was analysed relative to the potentially learned stellar north as well as relative to geomagnetic north.
The fact that the group observing the rotating artificial starry sky for 21 nights could orient in the proper migratory direction is in line with other studies where night-migratory songbirds were exposed to a rotating starry sky for 2–4 weeks. Birds from these previous studies also oriented relative to the observed star pattern (Emlen 1970; Wiltschko et al. 1987; Weindler et al. 1996, 1997) and calibrated their magnetic compass according to the memorized stellar north (Able and Able 1990b). Surprisingly, it seems as though 1 night or even 7 nights are not sufficient for birds to determine the celestial centre of rotation and calibrate their magnetic compass accordingly.
However, there is an alternative explanation for our results that must be considered. Experiments by Emlen (1969) suggest that star compass learning may become impaired at the onset of autumn migration. Since the middle (7 nights) and the short (1 night) learning groups had their first exposure to the rotating artificial starry sky later in the pre-migratory period than the long (21 nights) learning group, it could be that the number of learning days was not the most important parameter for establishing a functional star compass. Alternatively, it could be that the birds of the long learning group were the only group that observed the rotating artificial starry sky in their sensitive period.
Previous experiments on star compass learning were done with long distance migrants like Garden Warblers (Wiltschko et al. 1987) or Pied Flycatchers Ficedula hypoleuca (Weindler et al. 1995) that have relatively short time available between fledging and onset of migration. In these species, the sensitive period is assumed to be 15–35 days after fledging, which corresponds to the phase of their life when Eurasian Reed Warblers Acrocephalus scirpaceus—another long distance migrant—are known to show pre-migratory nocturnal activity and to make exploratory flights around their natal site (Mukhin et al. 2005). As European Robins from our experiments fledged in June, and this species does not start migration before the beginning of September, the pre-migratory period spans more than 2 months in which the birds can learn the star compass. It is not exactly known when within these 2–3 months the sensitive period lies in this species.
However, in general, if the critical stimulus needed for learning is not experienced during the core sensitive period, the sensitive period can usually be prolonged (Bischof 2007). Therefore, we exposed our birds to celestial rotation late in the pre-migratory period. It should be mentioned here, however, that imprinting on the natal site, another important imprinting process which is known to take place in the pre-migratory period of long distance migrants such as Pied Flycatchers (Berndt and Winkel 1979) and Collared Flycatchers Ficedula albicollis (Löhrl 1959), the sensitive period is known to be limited to a short time before the post-juvenile moult, and it does not seem to be extendable (Löhrl 1959). However, in the mentioned studies, the relevant cues were generally constantly available during the process of natal imprinting, whereas, in our study, the relevant cues for star compass learning were not constantly available.
To sum up the last sections, birds in our study that were exposed to artificial stellar rotation for 1 week or less may have failed to learn the centre of celestial rotation either because star compass learning generally takes more than 1 week and/or because the exposures to the rotating artificial starry sky may not have taken place during the sensitive period for star compass learning in this species.
Our data also support the view that celestial rotation serves as the primary reference for calibrating the magnetic compass during ontogeny (e.g. Emlen 1970; Bingman 1983; Able and Able 1990a, b; Prinz and Wiltschko 1992; Weindler et al. 1995; but see Alerstam and Högstedt 1983) and that the magnetic compass derives directional significance from a celestial reference (Emlen 1970, 1972; Able and Bingman 1987; Shumakov and Zelenova 1988; Beason 1992; Weindler et al. 1995).
If the magnetic compass of European Robins was functional without calibration to a celestial reference, all four groups should have been oriented relative to magnetic north (Fig. 3a–d). None of the four groups were actually oriented relative to the geomagnetic field, despite the fact that they had access to an undisturbed magnetic field during their entire life. Furthermore, our long learning group was only orienting in the seasonally appropriate migratory direction when the learned position of the centre of the artificial starry sky was considered, but not to the geomagnetic field itself. This was observed in the long learning group during the first autumn as well as during the following spring, showing that they had celestially calibrated their magnetic compass. It should also be stressed that the visual cues provided to the control group observing the stationary starry sky during the pre-migratory period should be meaningless to the birds and should therefore not have been detected as stars and, thus, should not have affected the development of their magnetic compass. This group is therefore equivalent to having a group being exposed to the magnetic field alone. To sum up, we found no inherent magnetic direction preference even though an inherent magnetic direction preference has been reported for other passerine migrants (Wiltschko and Gwinner 1974; Wiltschko et al. 1980; Bingman 1981, 1983; Beck and Wiltschko 1982; Able and Able 1990a; Bletz et al. 1996). Instead, our study agrees with other studies suggesting that the magnetic compass needs to be calibrated by celestial rotation in order to be useful for migratory orientation (Emlen 1970, 1972; Able and Bingman 1987; Shumakov and Zelenova 1988; Beason 1992; Weindler et al. 1995).
In conclusion, our results suggest that either more than 7 nights and less than 21 nights of observation of stellar rotation is necessary for inexperienced European Robins to learn the centre of celestial rotation and to calibrate their magnetic compass accordingly, or that their sensitive period for learning the centre of celestial rotation ends in mid-August. Our results strongly indicate that celestial rotation is the fundamental directional reference for the magnetic compass in European Robins and that the magnetic compass of European Robins only seems to work after it has been calibrated to a celestial reference during ontogeny.
References
Able KP, Able MA (1990a) Ontogeny of migratory orientation in the Savannah Sparrow, Passerculus sandwichensis: calibration of the magnetic compass. Anim Behav 39:905–913
Able KP, Able MA (1990b) Calibration of the magnetic compass of a migratory bird by celestial rotation. Nature 347:378–380
Able KP, Bingman VP (1987) The development of orientation and navigation behavior in birds. Q Rev Biol 62:1–29
Åkesson S, Morin J, Muheim R, Ottosson U (2002) Avian orientation: effects of cue-conflict experiments with young migratory songbirds in the high Arctic. Anim Behav 64:469–475
Alerstam T, Högstedt G (1983) The role of the geomagnetic field in the development of birds’ compass sense. Nature 306:463–465
Batschelet E (1981) Circular statistics in biology. Academic, London
Beason RC (1987) Interaction of visual and non-visual cues during migratory orientation by the Bobolink (Dolichonyx oryzivorus). J Ornithol 128:317–324
Beason RC (1992) You can get there from here: responses to simulated magnetic equator crossing by the Bobolink (Dolichonyx oryzivorus). Ethology 91:75–80
Beck W, Wiltschko W (1982) The magnetic field as a reference system for genetically encoded migratory direction in Pied Flycatchers (Ficedula hypoleuca Pallas). Z Tierpsychol 60:41–46
Berndt R, Winkel W (1979) Verfrachtungs-Experimente zur Frage der Geburtsortsprägung beim Trauerschnäpper (Ficedula hypoleuca). J Ornithol 120:41–53
Berthold P, Helbig AJ (1992) The genetics of bird migration: stimulus, timing, and direction. Ibis 134:35–40
Bingman VP (1981) Savannah Sparrows have a magnetic compass. Anim Behav 29:962–963
Bingman VP (1983) Magnetic field orientation of migratory Savannah Sparrows with different first summer experience. Behaviour 87:43–53
Bingman VP (1984) Night sky orientation of migratory pied flycatchers raised in different magnetic fields. Behav Ecol Sociobiol 15:77–80
Bischof HJ (2007) Behavioral and neuronal aspects of developmental sensitive periods. NeuroReport 18:461–465
Bletz H, Weindler R, Wiltschko R, Wiltschko W, Berthold P (1996) The magnetic field as reference for the innate migratory direction in Blackcaps, Sylvia atricapilla. Naturwissenschaften 83:430–432
Chernetsov N, Kishkinev D, Kosarev V, Bolshakov CV (2011) Not all songbirds calibrate their magnetic compass from twilight cues: a telemetry study. J Exp Biol 214:2540–2543
Cochran WW, Mouritsen H, Wikelski M (2004) Migrating songbirds recalibrate their magnetic compass daily from twilight cues. Science 304:405–408
Emlen ST (1967a) Migratory orientation in the Indigo Bunting, Passerina cyanea. Part I: evidence for use of celestial cues. Auk 84:309–342
Emlen ST (1967b) Migratory orientation in the Indigo Bunting, Passerina cyanea. Part II: mechanism of celestial orientation. Auk 84:463–489
Emlen ST (1969) The development of migratory orientation in young Indigo Buntings. Living Bird 8:113–126
Emlen ST (1970) Celestial rotation: its importance in the development of migratory orientation. Science 170:1198–1201
Emlen ST (1972) The ontogenic development of orientation capabilities. In: Galler SR, Schmidt-Koenig K, Jacobs GJ, Belleville RE (eds) Avian orientation and navigation. NASA, Washington, pp 191–210 Special publication 262
Emlen ST, Emlen JT (1966) A technique for recording migratory orientation of captive birds. Auk 83:361–367
Engels S, Hein CM, Lefeldt N, Prior H, Mouritsen H (2012) Night-migratory songbirds possess a magnetic compass in both eyes. PLoS ONE 7:e43271
Gwinner E (1996) Circadian and circannual programmes in avian migration. J Exp Biol 199:39–48
Hein CM, Engels S, Kishkinev D, Mouritsen H (2011) Robins have a magnetic compass in both eyes. Nature 471:E11–E12
Kramer G (1950) Weitere Analyse der Faktoren, welche die Zugaktivität des gekäfigten Vogels orientieren. Naturwissenschaften 37:377–378
Liu X, Chernetsov N (2012) Avian orientation: multi-cue integration and calibration of compass systems. Chin Birds 3:1–8
Löhrl H (1959) Zur Frage des Zeitpunktes einer Prägung auf die Heimatregion beim Halsbandschnäpper (Ficedula albicollis). J Ornithol 100:132–140
Merkel FW, Wiltschko W (1965) Magnetismus und Richtungsfinden zugunruhiger Rotkehlchen (Erithacus rubecula). Vogelwarte 23:71–77
Moore FR (1978) Sunset and the orientation of a nocturnal migrant bird. Nature 274:154–156
Mouritsen H (2003) Spatiotemporal orientation strategies of long-distance migrants. In: Berthold P, Gwinner E, Sonnenschein E (eds) Avian migration. Springer, Berlin, pp 493–513
Mouritsen H, Larsen ON (2001) Migrating songbirds tested in computer-controlled Emlen funnels use stellar cues for a time-independent compass. J Exp Biol 204:3855–3865
Mouritsen H, Mouritsen O (2000) A mathematical expectation model for bird navigation based on the clock-and-compass strategy. J Theor Biol 207:283–291
Mouritsen H, Feenders G, Hegemann A, Liedvogel M (2009) Thermal paper can replace typewriter correction paper in Emlen funnels. J Ornithol 150:713–715
Muheim R, Phillips JB, Åkesson S (2006a) Polarized light cues underlie compass calibration in migratory songbirds. Science 313:837–839
Muheim R, Moore FR, Phillips JB (2006b) Calibration of magnetic and celestial compass cues in migratory birds—a review of cue-conflict experiments. J Exp Biol 209:2–17
Mukhin A, Kosarev V, Ktitorov P (2005) Nocturnal life of young songbirds well before migration. Proc R Soc Lond B 272:1535–1539
Prinz K, Wiltschko W (1992) Migratory orientation of Pied Flycatchers: interaction of stellar and magnetic information during ontogeny. Anim Behav 44:539–545
Sauer F (1956) Zugorientierung einer Mönchsgrasmücke (Sylvia a. atricapilla, L.) unter künstlichem Sternenhimmel. Naturwissenschaften 43:231–232
Sauer F (1957) Die Sternenorientierung nächtlich ziehender Grasmücken (Sylvia atricapilla, borin und curruca). Z Tierpsychol 14:29–70
Shumakov ME, Zelenova NP (1988) Ontogenesis of nonvisual orientation of Blackcaps (Sylvia atricapilla) In: Abstracts of the XII eastern Baltic ornithological conference, Vilnius, pp 255–257
Weindler P, Beck W, Liepa V, Wiltschko W (1995) Development of migratory orientation in Pied Flycatchers in different magnetic inclinations. Anim Behav 49:227–234
Weindler P, Wiltschko R, Witschko W (1996) Magnetic information affects the stellar orientation of young bird migrants. Nature 383:158–160
Weindler P, Baumetz M, Wiltschko W (1997) The direction of celestial rotation influences the development of stellar orientation in young Garden Warblers (Sylvia borin). J Exp Biol 200:2107–2113
Wiltschko W, Gwinner E (1974) Evidence for an innate magnetic compass in Garden Warblers. Naturwissenschaften 61:406
Wiltschko W, Wiltschko R (1972) Magnetic compass of European Robins. Science 176:62–64
Wiltschko W, Wiltschko R (1976) Interrelation of magnetic compass and star orientation in night-migrating birds. J Comp Physiol 109:91–99
Wiltschko W, Gwinner E, Wiltschko R (1980) The effect of celestial cues on the ontogeny of non-visual orientation in the Garden Warbler (Sylvia borin). Z Tierpsychol 53:1–8
Wiltschko W, Daum P, Fergenbauer-Kimmel A, Wiltschko R (1987) The development of the star compass in Garden Warblers, Sylvia borin. Ethology 74:285–292
Zapka M, Heyers D, Hein CM, Engels S, Schneider NL, Hans J, Weiler S, Dreyer D, Kishkinev D, Wild M, Mouritsen H (2009) Visual but not trigeminal mediation of magnetic compass information in a migratory bird. Nature 461:1274–1277
Acknowledgments
We thank Thomas Geiger and co-workers of the electronic and mechanical workshops of the University of Oldenburg for constructing the star test chambers and Dr. Nils-Lasse Schneider for technical support. We also thank Prof. Dr. Franz Bairlein for his generous support in allowing us to use the excellent facilities of the Institute of Avian Research, Wilhelmshaven, Germany, Kane Brides for assistance to find nests, and Andreas Lischke who provided essential help with hand-raising birds. We thank Inka Spiller, Maike Gärtner, Laura Ziegenbalg and Hanna Seekamp for help with conducting the experiments. Financial funding was provided by the German Federal Ministry of Education and Research (BMBF; “Varying Tunes”, 01 GQ 0962 to H.M.), the DFG (MO 1408/1-2 to H.M.), and the Volkswagenstiftung (Lichtenberg Professur to H.M.).
Author information
Authors and Affiliations
Corresponding authors
Additional information
Communicated by N. Chernetsov.
A. Michalik and B. Alert contributed equally to this paper.
Rights and permissions
About this article
Cite this article
Michalik, A., Alert, B., Engels, S. et al. Star compass learning: how long does it take?. J Ornithol 155, 225–234 (2014). https://doi.org/10.1007/s10336-013-1004-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-013-1004-x