
Abstract
The little-known taxon Cyornis [banyumas] magnirostris has long been treated as a race of the widespread Hill Blue Flycatcher C. banyumas, with which it was thought to be allopatric during the breeding season. On the basis of morphology, magnirostris has lately been considered a full species, endemic as a breeder to northeastern India. Our recent field work during migration and the breeding season (September 2005 and June–July 2006) has, however, resulted in the first records of magnirostris from northern Burma/Myanmar, establishing that its breeding range broadly overlaps the range of C. banyumas whitei. We demonstrate how historical factors, sources of error, including fraud, errors of omission and commission, and inferences based on lack of data have all negatively affected assessment of species limits and conservation status in this speciose group of flycatchers. We also provide evidence that the taxon C. [banyumas] lemprieri is highly distinct and should be treated as a full species.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The blue flycatchers of the Oriental Region comprise a speciose assemblage of taxa that are very similar despite having brilliantly plumaged males, and their taxonomy has therefore long vexed systematists (at least since Rothschild 1926). Within the Indian subcontinent and Southeast Asia, most taxa are relatively well known and common, with the notable exception of the distinctive and enigmatic Cyornis [banyumas] magnirostris (Blyth, 1849). Despite its relatively early description, magnirostris has long seemed rare and hence has remained poorly known. Stresemann (1925) was the first to place magnirostris as a race of his newly enlarged C. banyumas “Formenkreis”, implying that magnirostris is entirely allopatric with other included taxa. Although disputed by Robinson and Kinnear (1928), Stresemann’s (1925) treatment of magnirostris within banyumas has since been unanimously followed until Rasmussen and Anderton (2005) considered magnirostris to be a full species primarily on the basis of its distinctive morphology, especially in comparison with C. banyumas whitei, the form geographically nearest to it that Stresemann (1925) placed within banyumas, but this treatment has not been adopted in some subsequent works (e.g., Wells 2007).
During fieldwork in Putao District, North Burma/Myanmar, in September 2005, we collected two flycatchers (an adult male and a juvenile female) that we later determined to be magnirostris. These are the first records of this taxon from anywhere in North Burma/Myanmar (Smythies 1953, 2001; Robson 2000), but given that these were taken during the migration period, we could not ascertain whether they were in transit from the only historically known breeding grounds of the taxon in Sikkim, or whether they were on their breeding grounds. However, during further fieldwork in the same areas in North Burma/Myanmar in June and July 2006, we found the species to be locally among the most common understory passerines. We netted several adult and juvenile magnirostris there, conclusively demonstrating for the first time that the taxon breeds in Burma/Myanmar.
We present details of records of magnirostris and whitei, discuss their implications on the taxonomic status of magnirostris, and present the first mensural analyses bearing on its specific status. We demonstrate how historical factors, fraud, errors of omission and commission, and inferences based on lack of data have all negatively impacted assessment of species limits and hence conservation status in this group. In addition, we re-evaluate the specific status of C. [banyumas] lemprieri of the Palawan area (treated as specifically distinct by Taylor 2006).
Methods
To conduct an inventory of the birds of the Hkakabo Razi region (Northern Forest Complex, in northern Kachin State, Burma/Myanmar; Renner et al. 2007) we undertook five expeditions to the region, in January–March 2001, February 2004, September 2005, March 2006, and June–July 2006. During this field work we mist-netted birds at selected locations and determined species, body mass, and where possible sex and age for each captured bird. Individuals diverging obviously from available sources (King et al. 1995; Robson 2000; Smythies 2001; Rasmussen and Anderton 2005) were collected for further analysis in museum collections. Two magnirostris collected in September 2005 were exported to the NMNH bird collection (acronyms explained in the Acknowledgments), and 12 magnirostris were netted in June–July 2006, and photographs, measurements and body mass were taken.
For each specimen, we took the following measurements with digital calipers to the nearest 0.5 mm: culmen length from base of skull and from distal-most feathers on culmen ridge (not narial area); bill width from distal-most feathers and at proximal end of gape; bill height from feathers; flattened wing; Kipp’s distance (distance from wingtip to outermost secondary); alula length (from bend of wing to distal tip of longest alular feather); shortfall of tip of primaries 1–10 (P1 being the outermost, measured ascendantly) to tip (longest feather) of folded wing; tarsus length; hallux phalanx 1 length; hallux claw (unguis) length; tail length from insertion point between central rectrices; and tail graduation (distance from longest to shortest fully grown rectrices). Not all measurements were taken from all individuals. We also measured specimens from other institutions for comparison (see Acknowledgments). We used Systat 8.0 for statistical analyses.
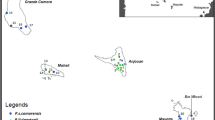
Geographic coordinates for the locations were derived from online resources (http://www.tageo.com/, accessed 5 September 2007), Collar et al. (2001), and The Times Atlas of the World: Comprehensive Edition (11th Edition, 1999). We converted all coordinates to decimal degrees if needed and plotted the location with ArcGIS 9.2 including attributes (taxon, collection date) to further analyze and model sympatric versus parapatric distribution of whitei and magnirostris (Fig. 2).
We compared plumage visually. Nomenclature of plumage parts basically follows Robson (2000). Ideally, we had specimens in hand and used artificial collection light to compare plumage colors; we used natural light under shady conditions in the field.
Results
Morphological analyses
Mensural analyses (Table 1) show that magnirostris is clearly distinct from all taxa now placed within banyumas (following the taxonomy of Dickinson 2003). It is larger in most bill measurements than all other taxa except the otherwise quite dissimilar lemprieri of Palawan (Table 1), and is most different from continental taxa including whitei. It is also significantly longer-winged than all taxa except lemprieri, and has significantly greater primary projection (Kipp’s distance) than all taxa in the study. Although long-winged and large-billed, magnirostris does not have longer tarsi or tail than most other taxa, and in fact, it is significantly shorter-tailed than whitei.
Principal component analysis (PCA) likewise shows the distinctness of magnirostris in comparison with all other taxa within banyumas (Fig. 1, Table 2). Principal component 1 (PC1) is a strong size axis, on which most measures are strongly positive. Only shortfall of primary 2 from tip, tarsus length, and tail length are negatively correlated with the other measures on this axis. On PC1, PC-scores of individual magnirostris show complete separation from all specimens of whitei (including lekahuni and deignani), and they overlap only with lemprieri. This axis shows that some banyumas (including ligus) and coeruleatus are almost as large as the smallest magnirostris. PC2 contrasts mainly shortfalls of primaries 1 and 2, wing, and tail versus bill width from nares, bill width from feathers, and culmen hook length; although this axis has a significant eigenvalue, it does not separate out any of the taxa. PC3, however, which is also a contrast axis, does segregate most lemprieri from magnirostris, and most banyumas (including ligus) from other taxa. This axis is a contrast primarily of culmen length, culmen hook length, tarsus length, and tail length versus Kipp’s distance (primary projection) and shortfalls of primaries 1 and 2. These contrasts largely reflect the different proportions of the two largest, longest-billed taxa, magnirostris and lemprieri. While magnirostris has very long primary projection and a long shortfall of primary 1, it has a relatively short tail and tarsi compared to lemprieri and the smaller taxa.

Principal component analysis of morphometrics of Cyornis taxa
Distributional analysis
Two magnirostris were captured and collected in September 2005, and 12 were netted in June and July 2006. We did not capture any magnirostris during the January–March trips despite intensive sampling. These results are consistent with data from elsewhere showing that magnirostris is highly migratory throughout its range. The fact that 11 individuals of magnirostris were netted at Nam Ti (27°24.61′N, 97°40.06′E; sometimes spelled Nam Hti) during just 7 days of sampling effort indicates that the species is actually locally common there at that time of year. In contrast, only one juvenile male was collected during 11 days of sampling effort in the Naung Mung plains (27°30.00′N, 97°48.13′E, 570 m a.s.l.; sometimes spelt Naung Mong or Naun Mong), suggesting that the species breeds primarily in the lower hills such as Nam Ti (950 m a.s.l.). We did not encounter any C. [banyumas] whitei during any of our field trips.
Despite the fact that we did not encounter whitei, specimen records (Fig. 2, Table 3) confirm the strongly migratory habit of magnirostris, and show that magnirostris and whitei are broadly parapatric at least. If the specimen records of whitei from northeastern India represent resident populations (which seems likely but requires further field work), the two taxa would be sympatric. There is a breeding record of whitei from as far north as Sinlum, Bhamo (24°25′N, 97°50′E), Burma/Myanmar (BMNH 1908.8.2.28, Harrington, “captured on nest”); in June 1901, a whitei specimen was collected at Kauri Kachin, Burma/Myanmar (approximately 25°45′N, 96°52′E; BMNH 1905.8.16.236, Rippon); and the species occurs in northwest Yunnan (Yangtze Valley, 27°20′N; BMNH 21.7.15.475, 18 September, Clarke) in summer. Non-breeding season specimen records of whitei from the north of its range include two specimens from Htingnan, Burma/Myanmar (26°36′N, 97°52′E; BMNH 1939.12.8.105-106, 11, 10 March 1939, Kaulback) and “Hpunkataung” (not definitely located, but probably Pumkahtawng at 25°22′N, 97°43′E, 658 m), northeast of Myitkyina (BMNH 1937.1.7.132, 25 February 1933, Stanford). Given that the Assam and Htingnan specimens are from almost as far north as any of the known breeding season records, whitei is at most a short-distance migrant in the northern part of its range and may well be resident throughout its range (Fig. 2).

Map of specimen locations of Cyornis banyumas whitei and magnirostris in Southeast Asia (minimum convex polygons link outermost localities from both species during breeding and non-breeding). Only museum specimens with clearly labeled localities were used. If available, data on specimen sex, age, and capture date is indicated also. Label numbers refer to localities as mentioned in Table 3
Comparison of relevant Cyornis taxa in Southeast Asia
To evaluate the relative distinctiveness of magnirostris within the entire banyumas species complex, we compared plumage characteristics at BMNH of the seven distinct taxa considered subspecies by Dickinson (2003). These (with range summaries from Dickinson 2003) are magnirostris (central and east Himalayas, Assam to south Burma/Myanmar, and Malay Peninsular), whitei (north and east Burma/Myanmar, south-central China, north Thailand, north and central Indochina), caerulifrons (central and south Malay Peninsular), banyumas (central and east Java), ligus (west Java), coeruleatus (Borneo), and lemprieri (Palawan, Balabac).
The remaining two taxa, deignani and lekahuni, both from southern Thailand, are arguably distinct at the subspecies level and were not considered here. The summary of plumage and other characteristics of these taxa in Table 4 shows that by far the most distinctive form of all those placed within banyumas in Dickinson (2003) is lemprieri of Palawan and associated islands. It differs strikingly in bill shape, overall color, and color pattern, especially in the female, although most of these differences are not apparent in illustrations accompanying Taylor (2006). The most distinct taxon in proportions is magnirostris, which is, however, quite similar in plumage to whitei. The Peninsular Malaysian form, caerulifrons, is similar to whitei except in its noticeably richer, deeper plumage overall. The two Javan taxa (banyumas and ligus) are similar to each other but fairly distinctive from other taxa, and the Bornean taxon (coeruleatus) is even more distinctive than the Javan forms, especially in female plumage.
Discussion
Distribution
Although we have not found whitei in exactly the same areas as magnirostris, the ranges of the two taxa show at least substantial overlap. While it remains to be conclusively demonstrated that the two overlap significantly in the breeding season, this broad geographic overlap of magnirostris and whitei supports the view of Rasmussen and Anderton (2005) that magnirostris is a full species. Stresemann’s (1925) original treatment of magnirostris as a subspecies of banyumas was almost certainly predicated on the supposition that it was allopatric with whitei. Further field work will be needed to determine whether the two replace each other elevationally, and the fact that only magnirostris was found at Nam Ti and Naung Mung suggests that this may be the case.
Future research should include sampling at higher elevations to determine the elevational limits of both taxa and whether they indeed occur together in the breeding season. In northern Myanmar, magnirostris and whitei differ distinctly in several aspects, more so than in, for example, races of banyumas from southern Thailand. Since magnirostris and whitei are highly distinct despite being at least parapatric (if not truly sympatric), it is unlikely that gene flow occurs at a level that would justify considering magnirostris as a subspecies. Therefore, we consider that, under the Biological Species Concept, the best treatment is to consider magnirostris as a full species.
Taxonomic re-evaluation of other taxa in the C. banyumas complex
The form lemprieri has sometimes been considered a full species, the Palawan Blue Flycatcher Cyornis lemprieri (e.g., Taylor 2006) and we strongly recommend adopting this treatment, because the form is far more distinct than are many other species-level taxa within Cyornis. It is especially highly differentiated from any of the Sundaic forms, including the geographically nearest Bornean caerulifrons, and there seems no obvious reason to link it with banyumas as opposed to several other regional Cyornis species.
Other taxa currently placed within banyumas are less obviously different, but still require further study to ensure that this species constitutes a monophyletic lineage. Those of particular concern are the two Javan subspecies banyumas and ligus, and the Bornean caerulifrons, which seems a likely candidate for full species status. Should banyumas and ligus be shown to be not closely related to the Southeast Asian taxa, whitei has priority as the species name for the latter group (when magnirostris is considered a separate species).
Causes of confusion regarding C. magnirostris
The case of Cyornis magnirostris illustrates many of the kinds of problems that cloud the understanding of species limits and therefore accurate estimation of avian diversity. These problems (summarized below for Cyornis) include error due to label switching, overlooked material, fraud, conclusions drawn in the absence of adequate data, misidentification, and mapping error. As for label switching, shortly after magnirostris was originally described by Blyth 1849 (date correct according to Dickinson 2004), a new supposedly African species Muscicapa riisii Hartlaub 1857 was described, but much later this was shown to be probably a female of C. magnirostris to which the legs and label of another species had been reattached (Sclater 1924; Winkler 2003).
Although correctly identified, the Assam specimens of whitei that suggested breeding season overlap with magnirostris were long overlooked or confounded with magnirostris. The near-total lack of knowledge of bird distributions in northern Myanmar, and particularly during the monsoon, resulted in Stresemann’s (1925) decision—based on the data available to him—to lump magnirostris within banyumas. The strongly migratory habit of magnirostris, known to or suspected by earlier ornithologists, was long confounded by specimens Meinertzhagen stole and re-labeled as having been collected in winter in Sikkim (Rasmussen and Prys-Jones, manuscript). There are several recent sight records of magnirostris from Nepal through Arunachal Pradesh in the Indian subcontinent, but none have been presented with sufficient detail to allow confident reassessment, and some are very likely to be whitei, especially given that it had long been overlooked for the Indian subcontinent; others (for example those from Nepal) could be the very similar C. tickelliae, from which identification of males is difficult under field conditions and especially before the advent of the modern field guides of the past decade. Rasmussen and Prys-Jones (manuscript) found that the only magnirostris specimens purportedly taken in the winter in Sikkim, which had led others to the conclusion that it is resident in the Himalayas, were in the Meinertzhagen collection and clearly bear fraudulent data (see also Rasmussen and Anderton 2005). Baker (1933) also claimed that magnirostris occurs in “eastern Assam” in winter, and that it breeds in the hills south of the Brahmaputra, but this claim must be evaluated in light of considerable evidence casting doubt on many of Baker’s records (Rasmussen and Anderton 2005), and misidentification is another possibility (see below). All genuine specimen evidence shows that magnirostris is present in the Himalayas only during the breeding season, and winters in Tenasserim and Peninsular Thailand. Hence, migratory status of magnirostris was suggested by Robinson and Kinnear (1928), considered established by Stresemann and Meyer de Schauensee (1936), and followed by, e.g., Riley (1938), but obscured largely by Meinertzhagen’s frauds in more recent works (Ali and Ripley 1968, 1983; Ali 1974; Grimmett et al. 1999; Kazmierczak 2000). The highly migratory habit of magnirostris therefore contrasts strikingly with the nearly or entirely resident status of whitei (see next paragraph).
We located two long-overlooked specimens of whitei collected in December 1901 from Margherita, Assam, northeastern India (Table 3). These specimens had previously been correctly listed (Robinson and Kinnear 1928) and mapped (Stresemann and Meyer de Schauensee 1936) as whitei, but were erroneously omitted from later influential regional literature: while Ripley (1961, p. 432) listed Muscicapa banyumas magnirostris, he did not mention whitei. The re-location of the Margherita whitei specimens suggested the possibility that whitei and magnirostris might be sympatric, but this could not be confirmed, as no breeding specimens of magnirostris were known from so far east in Assam. Subsequently, we have located another certain and two probable Indian specimens of whitei. While whitei is generally considered non-migratory (e.g., Robson 2000), and we consider this view to be probably correct, the basis for this assumption is unclear (see above). Despite clarifying the migratory and taxonomic status of magnirostris, Rasmussen and Anderton (2005) mistakenly mapped the species for Bhutan, although it had not yet been definitely recorded there. A recent sight and photographic record from Bhutan (Bishop 2006) is inconclusive, as the photograph cannot be located (K.D. Bishop, in litt. 30 November 2006). Another recent record (Farrow 2008) from April 2008, however, seems plausible, being accompanied by confirmatory identification details.
Baker’s (1933) report of magnirostris wintering in Assam probably refers to whitei, because there are no dated winter specimens of the former from the region, while we now know of three definite (Margherita and Lushai Hills) and two further probable (Sadiya and Tenga Pani, Manbum, near Sadiya; definitely not magnirostris) whitei specimens from north-eastern India, all from December to February. The fact that all are from winter does not prove the taxon only occurs in India at that time, because most specimen collection in the region has taken place during winter months when the climate is far more pleasant. Although Baker (1933) mentions collecting nests of magnirostris in Margherita, Assam, with H.N. Coltart, Coltart’s specimens from Assam (AMNH 450724, AMNH 450725) are whitei, not magnirostris. While this does not prove that Coltart only collected whitei, it does suggest that Baker may have been confusing the two taxa. Despite the fact that several egg sets in the BMNH labeled as magnirostris exist from the hills south of the Brahmaputra, their specific identity cannot be validated using currently available methods, since they are externally indistinguishable from whitei and rubeculoides, and possibly other species. DNA techniques that may allow specific identification of small eggs are in development (R. P. Prys-Jones, personal communication, July 2007). The fact that there are three Cachar specimens of magnirostris may be taken as evidence that the species breeds there (as indeed may be the case), but one is undated, and the other two are from dates consistent with birds taken in migration (25 April and 3 May). If magnirostris does indeed breed in Cachar and in the Khasi Hills, as contended by Baker, it is surprising that Koelz, who collected extensively there in the summer months (June–August), did not collect any. As with Baker’s other nesting records and non-voucher-based observations (some of which seem extremely improbable), independent corroboration is needed before they can be taken at face value (see Rasmussen and Anderton 2005).
Much of the confusion surrounding C. magnirostris can be directly attributed to the fact that very little ornithological fieldwork has been done during the summer months because of the unpleasant monsoon conditions and abundant leeches during this time, and thus this species has been overlooked on its breeding grounds. In fact, no definite records from the breeding grounds in the Indian subcontinent are known from the twentieth Century. However, the information that several specimens were taken by Mandelli’s collectors in Sikkim suggests that it is or was locally common there during the breeding season, as we have found to be the case at Nam Ti, Burma/Myanmar. We expect that further fieldwork at the appropriate elevations during the monsoon months will result in improved knowledge of this migratory species’ distribution in the eastern Himalayas, as well as resolving the issue of breeding sympatry with whitei.
Further taxonomic issues require re-evaluation in this group. The form occurring in Palawan and Balabac, C. [banyumas] lemprieri, is particularly distinctive in several respects and has at times been considered a full species, a treatment certainly justified. Of the other taxa recognized by Dickinson (2003) as conspecific, banyumas and ligus of Java as well as coeruleatus of Borneo, are distinctive, and further study including of vocalizations and genetics may consider it as one or two separate species.
In conclusion, we recommend the treatment of Cyornis magnirostris as a full species, for which the English name Large Blue Flycatcher is appropriate. This name was used in Rasmussen and Anderton (2005) in lieu of the common name Large-billed Blue Flycatcher used earlier for magnirostris in the twentieth century, because the name Large-billed Blue Flycatcher is currently widely used as the common name of Cyornis caerulatus (not C. banyumas coeruleatus) of Borneo and Sumatra (Dickinson 2003), even though C. caerulatus has a smaller bill than does C. magnirostris. We also strongly recommend that the highly distinctive C. lemprieri once again be treated as specifically distinct, as was done in Dickinson et al. (1991), and that the established common name Palawan Blue Flycatcher again be used (Taylor 2006).
Conservation status of C. magnirostris
Evaluation of the threat status of C. magnirostris is needed, given that it is so little known and that very few recent records exist. Due to its apparent scarcity, Rasmussen (2005) suggested that magnirostris may qualify for treatment as an endangered species. Although our findings from the breeding season in northern Myanmar show that, at least in Nam Ti, it is very locally among the most common forest birds, this may not be the case elsewhere. Further breeding season surveys in similar areas are needed to establish the presence and status of this species.
Zusammenfassung
Brutnachweis von Cyornis [banyumas] magnirostris im Norden von Kachin State (Birma/Myanmar), mit taxonomischen Betrachtungen zu Cyornis
Das wenig bekannte Taxon Cyornis [banyumas] magnirostris wurde lange als Unterart des sehr weit verbreiteten C. banyumas behandelt und beide galten bislang während der Brutsaison als allopatrisch. C. magnirostris wird allerdings oft aufgrund der Morphologie als eigenständige Art betrachtet und galt bislang als endemischer Brutvogel im Nordosten Indiens. Unsere Feldarbeit im südöstlichen sub-Himalaja während der Zug- und Brutzeit (September 2005 und Juni-Juli 2006) lieferten die ersten Beobachtungen von C. magnirostris im Norden Birmas/Myanmars, so dass jetzt davon ausgegangen werden muss, dass die Brutgebiete beider Taxa sich deutlich überschneiden. Wir zeigen ferner, wie historische Ereignisse und Fehlerquellen inklusive Fälschungen sowie Fehlschlüsse aufgrund mangelnder Informationen die Artabgrenzung, Taxonomie und den Schutz der Art und der ganzen Cyornis-Gruppe negativ beeinflusst haben. Abschließend diskutieren wir Unterschiede von C. [banyumas] lemprieri von C. banyumas und schlagen vor, diese Taxon ebenso als vollwertige Art zu behandeln.
References
Ali S (1974) Handbook of the birds of India and Pakistan, vol 10. Oxford University Press, Oxford
Ali S, Ripley SD (1968) Handbook of the birds of India and Pakistan. Oxford University Press, New York
Ali S, Ripley SD (1983) Handbook of the birds of India and Pakistan. Compact edn, Oxford University Press, Bombay
Baker ECS (1933) The nidification of birds of the Indian Empire, vol II. Turdidae-Sturnidae, Taylor & Francis, London
Bishop KD (2006) The birds and mammals recorded on the 2006 VENT Bhutan tour. http://www.ventbird.com/images/birdlists/birdlist_478.pdf. Accessed 21 January 2008
Collar NJ, Andreev AV, Chan S, Crosby MJ, Subramanya S, Tobias JA (2001) Threatened birds of Asia: the BirdLife International red data book. Part B:1517–3038. BirdLife International, Cambridge
Dickinson EC (ed) (2003) The Howard and Moore complete checklist of the birds of the world. 3rd edn. Princeton University Press, Princeton
Dickinson EC (2004) Systematic notes on Asian birds. 47. Blyth’s “Catalogue of the birds in the Museum Asiatic Society” and his 1849 supplemental note, with historical comments. Zool Verh Leiden 350:167–181
Dickinson EC, Kennedy RS, Parkes KC (1991) The birds of the Philippines, an annotated check-list. Tring: British Ornithologists’ Union (check-list No. 12)
Farrow D (2008) Bhutan. 9–28 April 2008. http://www.birdquest.co.uk/pdfs/report/BHUTAN%20REP%2008.pdf. Accessed 08 December 2008
Grimmett R, Inskipp C, Inskipp T (1999) Birds of India: Pakistan, Nepal, Bangladesh, Bhutan, Sri Lanka and the Maldives. Princeton Press, Princeton
Kazmierczak K (2000) Field guide to the birds of the Indian Subcontinent. Pica Press, Mountfield, Sussex
King B, Woodcock M, Dickinson EC (1995) Birds of Southeast Asia. Collins, New York
Rasmussen PC (2005) Biogeographic and conservation implications of revised species limits and distributions of South Asian birds. Zool Med Leiden 79–3:137–146
Rasmussen PC, Anderton JC (2005) Birds of South Asia: the Ripley Guide, 2 vols. Lynx Edicions, Barcelona
Renner SC, Rappole JH, Leimgruber P, Kelly DS, Nay Myo Shwe, Thein Aung, Myint Aung (2007) Land use in the Northern Forest Complex of Myanmar (Burma)—new insights for conservation. Oryx 41:27–37
Riley JH (1938) Birds from Siam and the Malay Peninsula. US Natl Mus Bull 172
Ripley SD (1961) A synopsis of the birds of India and Pakistan. Bombay Natural History Society, Bombay
Robinson HC, Kinnear NB (1928) Notes on the genus Cyornis, Blyth. Novit Zool 34:231–261
Robson C (2000) A field guide to the birds of South-East Asia. New Holland, London
Rothschild L (1926) On the avifauna of Yunnan, with critical notes. Novit Zool 33:189–343
Sclater (1924) Bulletin of the British Ornithologists’ Club CCXCII. December 29, 1925
Smythies BE (1953) The birds of Burma, 3rd edn. Oliver & Boyd, Edinburgh
Smythies BE (2001) The birds of Burma, 4th edn. Natural History Pub, Kota Kinabalu, Borneo
Stresemann E (1925) Über einige Cyornis-Arten. Ornithol Monatsberi 33:45–53
Stresemann E, Meyer de Schauensee R (1936) Notes on some South Asiatic species of the genus Cyornis. Proc Acad Nat Sci Philadelphia 88:337–351
Taylor PB (2006) Family Muscicapidae (Old World flycatchers), pp 56–163. In: del Hoyo A, Elliott A, Christie D (eds) Handbook of the birds of the world, vol 11. Lynx Ediciones, Barcelona
Wells DR (2007) The birds of the Thai-Malay Peninsula, vol 2. Passerines, Christopher Helm, London
Winkler R (2003) Kommentierte Liste der Typen in der Basler Vogelsammlung. 9 p. Naturhistorisches Museum Basel, Basel. http://www.nmb.bs.ch/aves.pdf. Accessed 21 January 2008
Acknowledgments
We thank the Nature and Wildlife Conservation Division of the Union of Myanmar Forestry Department, and especially Director U Khin Maung Zaw, for permission to conduct the study (Myanmar Collection and Export Permit #SI/4697/2004). We thank U Aung Khin and Daw Thandar Kyi (Myanmar Gateway Tours) who organized much of the expedition for us. The Smithsonian Institution, Office of the Undersecretary for Science, provided support for an expedition through the Abbott Fund. Additional support came from the Alexander Wetmore Fund, Bird Division, US National Museum of Natural History (NMNH), Smithsonian Institution, the National Geographic Society, and the Chapman Collection Study Grant (AMNH). Animal use activities were cleared by the CRC-IACUC on 03 January 2002 (IACUC Proposal # 01-34) and 20 December 2006 (# 06-27) as a continuation of our avian inventory in Myanmar. Field assistants included U Kyaw Lin, U Tu Myint U, U A Jo, U Myint Kyaw, Bran Shaung, and San Naing Dee. In particular we thank G.R. Graves, S.L. Olson, J.P. Dean, B.K. Schmidt, and C.M. Milensky (NMNH); R.P. Prys-Jones and M.P. Adams, The Natural History Museum (BNHM), Tring, UK; D. Willard, Field Museum of Natural History (FMNH), Chicago; P.R. Sweet, American Museum of Natural History (AMNH), New York; J.V. Remsen and S.W. Cardiff, Museum of Natural Science, Louisiana State University (LSUMNS), Baton Rouge; and N. Rice and L. Joseph, Academy of Natural Sciences of Philadelphia (ANSP), for assistance, loans, and use of collections. Information on putative field records of magnirostris and discussion was provided by K.D. Bishop and M. Catsis, and I. Heynen provided further discussion on previous versions of the manuscript.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Communicated by J. Fjeldså.
Rights and permissions
About this article
Cite this article
Renner, S.C., Rasmussen, P.C., Rappole, J.H. et al. Discovery of the Large Blue Flycatcher Cyornis [banyumas] magnirostris breeding in northern Kachin State (Burma/Myanmar) and taxonomic implications for the Cyornis-group. J Ornithol 150, 671–683 (2009). https://doi.org/10.1007/s10336-009-0395-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-009-0395-1