
Abstract
One hypothesized function of conspicuous mobbing of intruders by bird nest owners is to attract neighbouring birds (“calling for help” hypothesis) or third-party predators (“attract the mightier” hypothesis). These may help the nest owners by distracting and/or attacking the mobbed intruder. To date, these hypotheses have been studied solely during the mobbing of predators. Here, for the first time, I have studied mobbing attraction in the context of brood parasitism. I experimentally tested the Blackcap (Sylvia atricapilla), a small passerine with a highly aggressive and conspicuous nest defence behaviour. I elicited the aggressive responses of Blackcaps by presenting stuffed dummies of the brood parasitic Common Cuckoo (Cuculus canorus) and controls near their nests. At 32% of the nests (n = 75), the responses of the Blackcaps to dummies attracted up to 15 birds per trial from 21 passerine species. Most of the attracted birds were heterospecifics and rarely participated in mobbing; thus the “calling for help” hypothesis was not supported. No potential predators of the Cuckoo were attracted despite them living in the study area and despite prolonged mobbing by Blackcaps; thus rejecting the “attract the mightier” hypothesis. I argue that this hypothesis is unlikely to apply to typical avian predators during nest predation acts because these only last for several seconds. The number of attracted birds was a positive function of the owner’s intensity of nest defence as measured by the rates of alarm calling but not visual cues (rates of attacks). Suitable and unsuitable Cuckoo hosts did not differ in their behaviour in the vicinity of defended nests. The observed pattern of the positive correlation between the intensity of nest defence and the number of attracted birds is most likely a proximate by-product of the conspicuous nest defence by Blackcaps (but may well be adaptive for recruiting neighbours themselves). Thus, the mobbing behaviour of the Blackcap is directed towards the brood parasite and not towards other audience predators or potential recruits to the mob.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
When faced with a dangerous enemy, e.g. a predator, many bird, mammal, fish and insect species perform “mobbing”, i.e. they approach the enemy, rapidly move to and from it and sometimes even physically contact it (Curio 1978; Caro 2005). In birds, e.g. those defending their nest, another typical feature is alarm calling with a broad frequency spectrum (Curio 1978; Hurd 1996). It is believed that acoustically and visually conspicuous mobbing may also serve as signals from owners of the nest (hereafter “focal pair”) not only to the mobbed predator but also to third-parties—neighbouring birds (“calling for help” hypothesis; Rohwer et al. 1976; Hurd 1996) as well as additional predators (“attract the mightier” hypothesis; Curio 1978; Högstedt 1983).
Under the former scenario (Hurd 1996), calls may serve as a request by the focal pair for help from other conspecifics and heterospecifics. Attracted birds should not just observe the focal pair but should actively help to mob the predator by alarm calling (to confuse it) or even attacking it (to drive it away). The “calling for help” hypothesis predicts positive correlations between the intensity of mobbing by the focal pair and the number of and intensity of mobbing by attracted birds. However, even the mere presence of more individuals near the intruder was hypothesised to inform the intruder that it was discovered (“perception advertisement” hypothesis; Flasskamp 1994) and/or to confuse it (“confusion” hypothesis; Curio 1993). Both the focal pair and the recruits lured to the nest environs may also benefit from a dilution of risk to them (“risk dilution” hypothesis; Curio 1993).
Under the “attract the mightier” hypothesis (Curio 1978), the behaviour of the focal pair should attract other physically stronger individuals that may deter the primary intruder, i.e. especially predator(s), either avian or mammalian. This may be especially important when there is a high discrepancy between the size, armament and the physical capability of the defending parent and the dangerous primary intruder (Bourne 1977; Krams and Krama 2002). It is predicted that the probability of predator attraction and the number of attracted predators should be a positive function of the mobbing intensity by the focal pair. To fit the “attract the mightier” hypothesis, the attracted predators should actively attack the primary intruder and ignore the defended nest of the focal pair. In general, it is predicted that in structurally complex environments (e.g. in forests with dense undergrowth) the main cue for attraction should be acoustical (e.g. call rates) rather than visual (e.g. number of contact attacks).
To summarise, the “calling for help” scenario predicts both conspecific and heterospecific individuals should take part in mobbing, whereas “attract the mightier” scenario in principle does not concern either conspecifics or non-predatory heterospecifics but predicts an attraction of heterospecific predators. Obviously, these scenarios are not mutually exclusive because the focal pair may attract both conspecifics, non-predatory heterospecifics and predators. Moreover, the predator may be attracted not primarily (by activities of the focal pair) but secondarily (by other attracted birds) as well.
Alternatively, attraction of neighbours or predators by mobbing performed by the focal pair may not provide any benefits to these nest owners, and the attraction may be just a proximate response of attracted individuals to conspicuous mobbing (which itself may be beneficial to the focal pair). In other words, focal pair mobbing responses may be positively selected for because of their direct effect on an intruder, but not because they would attract any other mobbers from the neighbourhood. Here, in contrast to the focal pair, the attracted visitors may gain benefits themselves (e.g. may learn about the appearance of dangerous enemies or gain information on the presence of predators in the vicinity of their own territories; see “Discussion”).
The majority of avian nest defence studies to date have focused primarily on nest owners (Montgomerie and Weatherhead 1988), with marginal attention being paid until recently to attracted birds and their behaviour (Winkler 1994; Krams and Krama 2002; Krams et al. 2006). To my knowledge, no study so far has focused on the attraction of other individuals in relation to mobbing of brood parasites (but see Payne et al. 1985, for group mobbing by “helpers at the nest” in a cooperatively breeding species). Here, it is predicted that brood parasite hosts should be attracted more often than non-hosts. Alternatively or additionally, potential hosts should respond more strongly (i.e. participate more actively in mobbing) than non-hosts.
I tested the predictions of the “calling for help” and “attract the mightier” hypotheses experimentally in a small European passerine, the Blackcap (Sylvia atricapilla), which shows highly aggressive and conspicuous nest defence (Røskaft et al. 2002; Grim 2005a). I provoked the Blackcap’s aggressive responses by presenting stuffed dummies of the Common Cuckoo (Cuculus canorus) near their nests in an area where this brood parasite is found commonly in sympatry with the study species. The Blackcap is substantially smaller than the Cuckoo (∼20 vs ∼120 g) and nests at high densities in Central European forests (personal observations). Therefore, this species appears to be a good model to test both the “calling for help” (see the high breeding densities) and “attract the mightier” hypotheses (see the body size discrepancy).
I tested the host’s responses throughout laying, incubation and nestling periods. The Cuckoo is a threat to Blackcaps not only during laying and early incubation stages when successful parasitism is possible, but also at the stage of late incubation and nestlings, as the Cuckoo is known to predate both eggs and nestlings at unparasitised nests (e.g. Wyllie 1975). In line with this, Blackcaps are aggressive towards the Cuckoo throughout the nesting cycle (see discussion in Grim 2005a).
Currently, the Blackcap is only rarely parasitised by Cuckoos (for references see Honza et al. 2004b, p. 178), mostly in Eastern Europe (I. Krams, personal communication). However, several lines of evidence clearly show that the Blackcap had an evolutionary interaction with the Cuckoo: (1) Blackcaps show a fine-tuned ability to reject alien eggs both in my study area (Honza et al. 2004b) and elsewhere, e.g. in Norway (Moksnes and Røskaft 1992) and Spain (Soler et al. 2002), (2) there is no evidence for conspecific parasitism in Blackcaps showing that rejection of Cuckoo eggs is not a by-product of adaptations that have evolved due to parasitism by conspecifics (Yom-Tov 2001), (3) Blackcaps are much more aggressive towards the Cuckoo than to innocuous sympatric species in my study area (but, unsurprisingly, they make recognition errors when faced with innocuous species very similar to the Cuckoo; for details see Grim 2005a), (4) there is ample direct evidence for Cuckoo parasitism in Blackcap nests dating from the fourteenth until early twentieth century in the Czech Republic (see discussion in Honza et al. 2004b) and elsewhere (Moksnes and Røskaft 1995), (5) Cuckoo eggs found in Blackcap nests are mimetic (Moksnes and Røskaft 1995; Honza et al. 2004b). Frequencies of natural Cuckoo parasitism reported in the above mentioned studies are certainly underestimated because Blackcaps are able to reject alien eggs within a few hours (Soler et al. 2002). The current apparent absence of the Cuckoo parasitism on the Blackcap in my study area has no bearing on the results of the current study because Blackcaps obviously had an evolutionary interaction with the Cuckoo until recently (see Honza et al. 2004b and above). Importantly, the colonisation of a new host and the decrease in the frequency of parasitism of an old host may happen very quickly (Brooke et al. 1998; Honza et al. 2004a), and a relic character of host anti-parasitic behaviour is common among brood parasite hosts in general (e.g. Rothstein 2001).
In sum, I predicted that more aggressive and conspicuous nest defence should attract more neighbours than a less aggressive response by the focal pairs. Furthermore, attracted birds should be neighbouring conspecifics or heterospecifics (“calling for help” hypothesis) and/or predators potentially dangerous to the Cuckoo (“attract the mightier” hypothesis). Under both hypotheses, these attracted birds should actively mob the dummy near the nest. Suitable Cuckoo hosts should arrive more often and/or respond more aggressively than birds unsuitable as hosts because for the latter the Cuckoo is not a threat.
Methods
I studied Blackcaps’ behaviour in the south-eastern part of the Czech Republic in a deciduous forest in the vicinity of Dolní Bojanovice village (48°52′N, 17°00′E), about 60 km south-east of Brno. Fieldwork was carried out from late April to late June 2000 and 2001. A portion of the data collected during this study has already been published (results of enemy recognition tests; Grim 2005a), but all data on attracted birds are entirely novel.
I generally followed the experimental procedure suggested by Sealy et al. (1998) except for the use of the predator dummy (for explanation and discussion, see Grim 2005a). When the Blackcap nest owners were not present at the nest, the taxidermic dummy, stuffed in a life-like position, was attached to a branch about 0.5 m from each nest, level with it and facing the nest rim. Responses of the focal pair (nest owners) were observed for 5 min (from the moment when the first parent arrived) from the hide set up at least 15 m from the nest. I tested each host nest (n = 75) with both the Cuckoo and a control dummy: the Pigeon (Columba livia) or the Blackbird (Turdus merula) (male). Both species were used as controls because they are neither parasites nor predators of Blackcaps and they do not compete with Blackcaps for either food or nest-sites; thus, they are totally innocuous from the Blackcaps’ point of view (see also Sealy et al. 1998). Furthermore, I have chosen the Pigeon because it is of similar colour, size and shape to the brood parasitic Cuckoo (the rationale suggested by Sealy et al. 1998), whereas the Blackbird was chosen because it is of different overall colour than the Cuckoo or Pigeon (black vs grey). This enabled two series of experiments to be run: (1) Cuckoo vs Pigeon (with close similarity of enemy and innocuous intruders) and (2) Cuckoo vs. Blackbird (with very different appearance of enemy and innocuous intruders). This design aimed to answer the questions “how good are enemy recognition abilities of the host?” and “how can the selection of particular control stimuli influence the results and interpretation of enemy recognition studies?” (see Grim 2005a).
Blackcaps attacked both the Cuckoo and the Pigeon at similar rates but totally ignored the Blackbird (Grim 2005a); therefore I concluded that Blackcaps recognized the Cuckoo as a special enemy (sensu Sealy et al. 1998). However, it should be noted that the Cuckoo plumage, its body size and its iris colour are similar to that of the Sparrowhawk (Accipiter nisus). Previously I showed that Blackcaps did not distinguish between a Cuckoo and a Pigeon dummy, suggesting that they might also not distinguish between a Cuckoo and perhaps a more similar Sparrowhawk. Nevertheless, it should be stressed that the level of similarity as perceived by humans is irrelevant to the issue whether Blackcaps perceive the Sparrowhawk as similar to the Cuckoo at all—obviously any similarity is in the eye of the beholder (for detailed discussion, see Grim 2005b). Specifically, experiments clearly showed that birds can finely discriminate between—by human standards—very similar stimuli and at the same time do not discriminate between stimuli that were perceived by humans as much more different (Honza et al. 2007). In general, passerines will recognize the Sparrowhawk from other cues because the costs of not recognizing this common and deadly predator are too high. Although some birds mob the Sparrowhawk (but still among several avian predators “fear most the European Sparrowhawk”, Curio et al. 1983, p. 86) others do not attack them at all (e.g. Duckworth 1991) or fear them (e.g. Smith and Hosking 1955, pp. 39–41) and either decrease their nest visitation rates (e.g. Ghalambor and Martin 2001) or even “freeze” (i.e. inhibit locomotion; e.g. Klump and Curio 1983). Rytkönen and Soppela (1995) showed in female Willow Tits (Parus montanus) that “the nest defence intensity was decreased when the number of encounters with Sparrowhawks increased”. Also, Blackcaps refrain from attacking deadly predators of adults (e.g. marten Martes martes; Bures and Pavel 2003) including the Sparrowhawk in another two study areas in the Czech Republic (V. Bicik and V. Pavel, respectively, personal communications). The existence of a specific “aerial-predator call” in the Blackcap is also suggestive of its enemy recognition abilities (Shirihai et al. 2001). Silent non-calling and non-attacking Blackcaps would obviously be useless for the testing of “calling for help” and similar hypotheses. Interestingly, during a recent literature survey I was unable to find a bird species that regularly attacks the Sparrowhawk by contact (see above). In contrast, Blackcaps very frequently (43% pairs) and repeatedly (dozens of contacts within the 5-min experiment in some tested pairs) physically contacted the Cuckoo dummy and spent the most time during experiments within 1 m of the Cuckoo dummy (Grim 2005a). In a striking contrast, both average (∼4–6 m) and minimum (∼2–3 m) approach distances to the Sparrowhawk dummy by small passerines are much longer (Curio et al. 1983; Rytkönen and Soppela 1995). Most importantly, in a study area close to my own field study site, V. Bicik (personal communication) found in a paired experiment design that Blackcaps fiercely attacked the Cuckoo mount but did not even approach the Sparrowhawk dummy. In a cue isolation experiment (see Gill et al. 1997), V. Bicik showed that even an isolated head of the dummy (without the body and tail) elicited strikingly different responses by Blackcaps; the Cuckoo head was attacked whereas the Sparrowhawk head was not approached. Therefore, I conclude that Blackcaps—just like any other passerines—do not mistake the Cuckoo for the Sparrowhawk. Even if the Cuckoo was mistaken by Blackcaps for the Sparrowhawk, this would still not pose a serious problem for the current study because testing the “calling for help” and the “attract the mightier” hypotheses for mobbing Blackcaps can be done independently of whether the mobbing is directed towards a perceived brood parasite or a perceived predator. In other words, I did not study why Blackcaps attract their neighbours but what are the proximate mechanisms of neighbour attraction during mobbing.
I separated the dummy presentations by 30-min intervals (Sealy et al. 1998) and randomised the order of dummies. There was no significant effect of the order of presentation on any of the studied behavioural variables (Grim 2005a), including the number of attracted birds to the Cuckoo dummy (U 36,39 = 1.20, P = 0.23). Therefore, I pooled the data from the first and second presentations of the Cuckoo dummy. Moreover, when I used only data from the first experiment at each nest I obtained qualitatively identical results.
First, I categorised behaviours according to relative scales. I scored the alarm rates on an ordinal scale from 0 to 2: 0 = no alarms, 1 = overall time spent calling <3 min, 2 = calling >3 min. Attacks (i.e. physical contacts by strikes) were scored from 0 to 2: 0 = no attacks, 1 = <5 attacks, 2 = >5 attacks). I also noted delay in arrival of nest owner(s) in minutes and the number of individuals that responded.
The total level of nest defence was measured in two ways. First, I ranked the total level of nest defence on a scale depending on the risk taken by tested bird(s) (see Montgomerie and Weatherhead 1988) from 0 to 2: 0 = silent watching of a dummy or only few (<∼10) alarms and bird(s) >5 m from the dummy, 1 = more (∼dozens) alarms and bird(s) <5 m from the dummy with close passes (i.e. mobbing), 2 = frequent (∼hundreds) alarms and attacks (i.e. bird physically contacted the dummy and pecked it especially around the eyes and on the nape, which is typical for small songbirds attacking an intruder near the nest; see Smith and Hosking 1955).
Second, I created a composite measure of nest defence by means of principal component analysis (PCA) on alarm calls, attacks and the number of nest owners responding. PC1 had Eigenvalue 1.70 and was strongly positively correlated with all three included variables (P < 0.0001).
I recorded all other birds attracted by Blackcap responses to dummies and also their behaviour as a dichotomous variable: (1) silent watching of the dummy and aggressive Blackcaps, or (2) participation in the nest defence by means of mobbing. Mobbing included alarm calls (passive defence) and close passes and dives (active defence; sensu Winkler 1994). A substantial proportion of the attracted birds appeared only in the control experiments (see difference between the Cuckoo and all trial datasets in Table 1). Therefore, in the descriptive part of the study, I pooled data from the Cuckoo and control experiments within each nest to increase sample sizes. Thus, there was no pseudoreplication as each nest was treated as a single data point.
When testing for the possible effects of host behaviour on the number of attracted birds (log-transformed data), I used both ordinal (the index of nest defence intensity, alarms, and contacts) and continual (PC1) measures of host responses. To increase sample size and thus the power of the test, I also fitted general linear mixed models (PROC MIXED in SAS; normal error distribution, parameters estimated by REML, degrees of freedom calculated using Kenward-Roger method; SAS Institute 2004) with the PC1 as predictor, the log-transformed number of attracted birds as a response and the brood identity as a random effect. The Blackcaps’ responses to the Pigeon dummy were similar to the Cuckoo dummy response, but they completely ignored the Blackbird dummy (Grim 2005a). Therefore, the inclusion of the Pigeon and Blackbird dummy data in the analyses provides a stronger test of the hypotheses under scrutiny as it spreads out the level of the independent variable (i.e. increases variation in Blackcaps’ nest defence behaviour; Kamil 1988).
There are various factors that may influence nest defence intensity and thus confound results of nest defence studies (reviewed in Montgomerie and Weatherhead 1988). However, in the current study, I was not interested in the effects of, e.g. season, brood age and size or number of previous visits on the intensity of nest defence, but on the effect of the nest owners’ responses to intruders on other birds in the vicinity of the defended nest. Thus, my aim was to study the second link in the chain “independent variables → the intensity of nest defence by the focal pair → the number and behaviour of the attracted birds” but not the first one (that link was studied separately; see Grim 2005a). However, I also conservatively tested for potential effects of various possibly confounding variables (see “Results”).
I also tested for relationships between abundance and frequency (proportion of experiments where birds appeared) of attracted birds during experiments and their respective availability, i.e. breeding densities (pairs/ha). Breeding densities for the nine most common species breeding in the study area were kindly provided by M. Čapek. Finally, I tested whether there were any differences in abundance, frequency and participation in mobbing of attracted birds and their suitability as Cuckoo hosts. Suitability of observed species as Cuckoo hosts was assigned according to Moksnes and Røskaft (1995), Røskaft et al. (2002) and Grim (2006).
Results
Characteristics of attracted birds “community”
At 32.0% of the nests (n = 75, Cuckoo and control experiments pooled at each nest), the vigorous nest defence activity of the Blackcaps attracted in total 108 individuals of 21 passerine species (Table 1). At 15 of these 24 nests, both the Cuckoo and control experiments attracted neighbours. At some nests (n = 11), particular species appeared during both the first and second experiment; thus, it is possible that these were the same individuals (attracted individuals were not individually marked which would be impossible due to logistic reasons either way). After correcting for this possibility, the total number of attracted birds was 83 (i.e. 25 individuals might have been attracted to both experiments within the particular nest; Table 1). However, at 9 out of 15 nests where both experiments attracted neighbours, these attracted birds’ “communities” differed in species composition and in particular species abundance between the first and second experiment within a nest. Thus, there was a substantial turnover of attracted species and individuals between the experiments within a nest. Therefore, the total and corrected abundance (Table 1) should be taken as an upper and lower limit, respectively, of the actual range of abundance.
Taking breeding densities as a surrogate measure of bird availability, the Blackcaps formed 25.7% of breeding pairs in the study area while only 8.4% of attracted birds were conspecifics (Table 1). Out of 75 first experiments only 5 attracted a conspecific Blackcap. Out of these five conspecifics only two approached the vicinity of the focal pair during both the first and second trial at the same nest. Another two Blackcaps were attracted only to the second trial at another two nests. None of attracted Blackcaps attacked the dummy and all birds alarm called very infrequently.
The most individuals attracted during one experiment were 15 birds of 6 different species, during a Cuckoo presentation. In the total sample, the smallest species was the Goldcrest (Regulus regulus), the biggest was the Jay (Garrulus glandarius). Attracted birds usually stayed about 3 to 5 m from the focal nest and remained silent. However, individuals of several species consistently alarm called (Chaffinch Fringilla coelebs, Great Tit Parus major, Blackbird, Robin Erithacus rubecula, and Blackcap; Table 1). Such passive defence was observed only at 24.0% of the nests (n = 75), and at 10 of these nests (n = 18) the passive defence was elicited in both experiments at the particular nest. I observed only one case of active defence: one Nuthatch (Sitta europaea) vigorously dived down from above on the dummy in one trial. During one experiment, a female Blackcap from the tested nest tried to drive away a “watching” Nuthatch.
Breeding density of the particular attracted species predicted neither their abundance nor frequency (both total and corrected; Table 1) during the experiments (Spearman rank correlations, n = 9, all P > 0.60). The abundance of attracted birds during the experiments and their frequency during experiments were strongly positively correlated (total: r s = 0.97, n = 21, P < 0.0001; corrected: r s = 0.94, n = 21, P < 0.0001).
Out of 21 species of attracted birds, 9 can be considered suitable while 12 are unsuitable Cuckoo hosts due to their hole-nesting habits or diet composition (Table 1). The abundance and frequency (both total and corrected) of suitable and unsuitable hosts did not differ (Mann–Whitney tests: all P > 0.50). Suitable (3 out of 9) and unsuitable (3 out of 12) hosts participated in mobbing with the same probability (Fisher exact test: P = 1.00).
Determinants of attraction
The number of attracted birds significantly increased with the ordinal measure of the intensity of nest defence in the Cuckoo dummy experiments (Fig. 1a; Kruskal–Wallis test: χ 2 = 8.29, df = 2, P = 0.02). The trend for attracted birds that helped with mobbing was similar but not significant (Fig. 1a; χ 2 = 4.13, df = 2, P = 0.13). As the index of nest defence combines potential vocal and visual cues triggering the attraction of other birds, I analysed these hypothesised effects of alarm calls and attacks separately (only attacks were considered as visual cues because Blackcaps neither hovered nor dived above dummies; Grim 2005a). Although the rates of calling and frequencies of contacts were positively correlated (r s = 0.35, n = 75, P = 0.002), it was still possible to disentangle their respective effects. Higher alarm rates attracted more birds (Fig. 1b; χ 2 = 8.26, df = 2, P = 0.02). There was a similar trend for the number of attracted birds that participated in mobbing (Fig. 1b; χ 2 = 5.01, df = 2, P = 0.08). However, there was no effect of contact attacks on the total number of attracted birds (Fig. 1c; χ 2 = 0.19, df = 2, P = 0.91). In contrast, the number of attracted active mobbers tended to increase with the intensity of attacks by nest owners (Fig. 1c; χ 2 = 4.90, df = 2, P = 0.09). When no alarm calls were given by Blackcaps other birds were never attracted (Fig. 1b), whereas birds were attracted even when no contacts were performed (Fig. 1c). These contrasting results indicate that the cue for attraction of alien birds was vocal (alarm calls) and not visual (attacks). However, when attracted birds were confronted with Blackcaps showing a higher intensity of nest defence (attacks) they too increased the intensity of their reaction (Fig. 1c).

The total number of attracted birds (open bars) and number of attracted birds that participated in mobbing (solid bars) of the Cuckoo (Cuculus canorus) dummy (mean + SE) as a function of a the intensity of nest defence, b alarm calling rates, and c the frequency of contact attacks (for details see “Methods”). Sample sizes are shown above columns
I also compared the number of nests with low (alarm = 1) and high (alarm = 2; see “Methods” and Fig. 1b) rates of alarm calling by nest owners. The proportion of nests which attracted at least some neighbours did not differ between trials with low (44.1%, n = 34) and high (30.8%, n = 26) alarm rates by nest owners (χ 2 = 1.11, df = 2, P = 0.29). Similarly, the rates of alarm calling did not affect the proportion of nests where neighbours participated in mobbing (26.5 vs 26.9%; χ 2 = 0.002, df = 2, P = 0.97). These results suggest that higher rates of alarm calling at the nest do not benefit the nest owners by attracting additional neighbours to join a mobbing group.
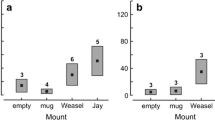
Dummy type had no significant influence on the number of attracted birds both when only the first trial per nest was analysed (Kruskal–Wallis test: χ 2 = 4.82, df = 2, P = 0.09) and when all data were analysed and the nest identity was included as a random effect (GLMM: F 2,87.1 = 1.77, P = 0.18) (Fig. 2). This was the case because in most experiments there were no attracted birds. The same conclusion was reached when only attracted birds participating in mobbing were analysed both in the first trial per nest analysis (Kruskal–Wallis test: χ 2 = 2.52, df = 2, P = 0.28) and all data analysis (GLMM: F 2,86.8 = 1.00, P = 0.37) (Fig. 2).

The total number of attracted birds (open bars) and number of attracted birds that participated in mobbing (solid bars) of different dummies (mean + SE). Sample sizes are shown above columns
Additional analyses also showed a consistent positive relationship between the number of alien birds attracted to the focal nest and the intensity of that nest owners’ responses to the Cuckoo dummy (PC1). There was a significant increase in the number of attracted birds with the intensity of reaction to the Cuckoo dummy (F 1,73 = 5.72, P = 0.02). The correlation on the same data was also positive and significant (r s = 0.25, n = 75, P = 0.03). When I restricted the analyses to attracted birds that mobbed, the relationship was again consistent (F 1,73 = 6.41, P = 0.01; r s = 0.27, n = 75, P = 0.02). However, the number of attracted birds that participated in mobbing only weakly increased with the group size (r s = 0.30, n = 24, P = 0.17).
When I analysed all data (both the Cuckoo and control experiment from each nest) and controlled for the effect of the particular nest, there was again a significant positive relationship between the intensity of reaction (PC1) and the total number of attracted birds (GLMM: F 1,144.5 = 10.33, P = 0.002). Excluding the non-mobbing attracted birds showed identical results (GLMM: F 1,144 = 9.04, P = 0.003). In addition, I found that an increase in the number of attracted birds with the intensity of reaction (PC1) remained significant in a general linear mixed model (GLMM) with explanatory variables including the dummy type (Cuckoo, Pigeon, Blackbird), the rank of presentation (first vs second), dummy × rank interaction, breeding stage (laying, incubation, nestlings), dummy × stage interaction, date in season, clutch/brood size, nest age and nest concealment (poor, average, excellent) as fixed effects and nest identity as a random effect. This remained true even when all these non-significant terms were conservatively kept in the model (the effect of PC1 in the full model: F 1,122 = 6.49, P = 0.012). The final model, which was selected according to corrected Akaike’s information criterion (AICc), contained only PC1 (the effect of PC1 in the final reduced model: F 1,144.5 = 10.33, P = 0.002).
The positive relationship between the intensity of nest defence and the number of attracted birds cannot be explained by an increasing availability of potential mobbers during the nesting season (see Pavel and Bures 2001) as there was no effect of date in season on the number of attracted birds per Cuckoo dummy experiments (r s = −0.03, n = 75, P = 0.79) or per nest (Cuckoo and control experiments pooled at each nest; r s = 0.01, n = 75, P = 0.93). Sex of the defending parent may influence the number of attracted birds (Winkler 1994). However, there were no differences in the number of attracted birds or attracted aggressive birds both per Cuckoo trials and per nest between nests divided into categories of (1) female more aggressive, (2) male more aggressive and (3) both nest owners similarly aggressive (Kruskal–Wallis tests, all P > 0.05).
Discussion
As predicted by the “calling for help” hypothesis, more intense nest defence by Blackcaps attracted more neighbouring birds to the defended nest. The cue for this attraction was most likely the rate of alarm calling, which concurs with results of previous studies in birds (Winkler 1994; Hurd 1996). The rate of contact attacks seemed to be unimportant for the attraction process itself, but might have affected the behaviour of foreign birds once attracted to the focal nest by increasing the probability of their alarm calling (which was the primary behavioural response of attracted birds). No predators were attracted despite living in the study area forest (e.g. Sparrowhawk Accipiter nisus, Buzzard Buteo buteo, stoat Mustela erminea; own observations), which falsifies the “attract the mightier” hypothesis. However, due to the field experimental nature of the current study, it was not possible to control for the presence of a potential predator in the area. Despite this, I believe the negative result is robust: nest defence activity by Blackcaps at experimental nests lasted for several minutes whereas both the predation by avian predators (Schaefer 2004; my own video-recordings) and egg-laying visits by the Common Cuckoo (Wyllie 1975) lasted only several seconds. Thus, under these experimental conditions, the chance of detecting the focal nest by both other birds and predators was larger by some two orders of magnitude than that under natural conditions. Because of the extremely short duration of avian predation acts (Schaefer 2004), the “attract the mightier” hypothesis can hardly apply to typical avian predators like the Jay during predation on eggs or nestlings. However, it might be applicable to mammalian predators, which spend several minutes at predated nests (Schaefer 2004), and also to perched avian predators (Curio et al. 1983).
Under the “calling for help” hypothesis, two scenarios were suggested: mobbing responses by genetically related individuals may be explained by kin selection (Rohwer et al. 1976), whereas reciprocal altruism may in principle explain aggressive responses by both unrelated conspecifics (Krams et al. 2006) and heterospecifics (Krams and Krama 2002). Because I studied an un-marked population of Blackcaps, I could not test these hypotheses directly. However, some results suggest that neither kin selection nor reciprocal altruism play a major role in shaping mobbing behaviour by Blackcaps. Although the kin relationships between Blackcap nest owners and attracted Blackcaps are unknown, the level and frequency of their “helping” with nest defence seems to be too low to be favoured by kin selection, even in the potential case of close relatedness between nest owners and attracted Blackcaps (moreover, extra-pair matings were never reported for the Blackcap, thus males helping their own extra-pair chicks in cuckolded nests are unlikely). Blackcaps are highly aggressive at their own nests—42.7% of the tested pairs (n = 75) physically attacked (contacted and pecked) the dummy near their nests—but not a single Blackcap attracted to an alien nest attacked the dummy there (but sample size is low; see Table 1). Furthermore, three out of five Blackcaps that were attracted to the first experiment at the nest ignored focal pair responses during the second trial at the same nests. Thus, there was no remarkably helpful behaviour on the part of attracted Blackcaps that could be reciprocated by the focal pair to attracted conspecifics in the future. Therefore, the “reciprocal altruism” version of the “calling for help” hypothesis also appears unsupported.
Mobbing based on reciprocal altruism between nest owners and attracted neighbours was reported recently in Pied Flycatchers (Ficedula hypoleuca) (Krams et al. 2006). In this species, conspecific neighbours arrived at tested nests in 100% of the cases (under natural conditions when they were not experimentally constrained from arriving), whereas conspecifics arrived in only 9.3% of cases at Blackcap nests in my study area (Table 1). In another study (Krams and Krama 2002), heterospecific neighbours attended 100% of the mobbing trials with Chaffinches whereas Blackcaps attracted whichever neighbours at only 32% of the nests in my study. This shows that Blackcaps nest defence is not very effective in attracting other birds.
The probability of being attracted did not correlate with the breeding density of attracted bird species in the study area (see also Winkler 1994). Breeding densities may be too rough a measure of attracted birds’ “availability”. It is more likely that the attracted birds were just those whose territories overlapped with territories of tested Blackcaps, or birds which happened to be near tested nests, e.g. when foraging there.
From the methodological point of view, the results of the current study should be conservative. Unsurprisingly, it was harder to note all behaviours of the more aggressive and active individuals in comparison to passive ones that just silently watched the dummy. Thus, there was a higher risk of overlooking some attracted birds in the former in comparison to the latter experiments. This trade-off may have caused an underestimation of the true relationship between the nest defence intensity and the number of attracted birds. Coupled with the consistently significant results of the analyses of various data sets (Cuckoo trials, all data) and the various measures of the nest defence intensity (ordinal scales, PCA), this suggests that the results of the current study are robust.
Costs and benefits of attractions for focal and attracted birds
The generally passive behaviour of attracted birds and their arrival after Blackcaps started to respond aggressively does not support a hypothesis that the intensity of nest defence by Blackcaps was influenced by potential mobbers (see Halupka and Halupka 1997). I also did not notice any differences in Blackcap nest owners' behaviour before and after other birds approached the focal nest. Interestingly, even in species with obligatory helpers these helpers may have no effect on the nest defence intensity of the breeding pair (Veen et al. 2000).
Through such a passive behaviour the attracted birds may gain information on the presence of predators in, or in the vicinity of, their own territories. Another benefit for attracted mobbers that alarm called may be “moving on” the predator or brood parasite from the general area, i.e. away from their own nests (Flasskamp 1994). Moreover, attracted birds participating in the “passive defence” (sensu Winkler 1994) may force the intruder to move on by “perception advertisement”, i.e. by informing it that its presence is revealed and the probability of its hunting success is consequently decreased, not only at the particular nest but also at those around (Flasskamp 1994). Attracted birds may also directly benefit from alerting their mates (Yasukawa 1989) and/or informing their own chicks at nearby nests that there is a risk of predation. Thus, chicks may be silenced by parental vocal signals and the probability of their death decreased (Davies et al. 2004). Attracted mobbers might in principle benefit in the future if parents whom they helped will reciprocate. However, this is unlikely in some short-lived small passerines (Winkler 1994) and in those showing low philopatry (e.g. the Blackcap population in the current study; see Grim 2005a). All these benefits may compensate for time, energy and risk of injury costs resulting from taking part in mobbing of a predator at alien nests (for review, see Curio 1978).
Loud alarm calling may result in additional costs of revealing the nest location to eavesdropping predators (Krama and Krams 2005). Blackcap alarm calling attracted very few potential predators of their nests—only one Jay (Table 1) and no Great-spotted Woodpeckers (Dendrocopos major). Both these nest predators are common in the study area (personal observations). Even the attracted Jay did not find the defended Blackcap’s nest (this can hardly be explained by “confusion” from mobbing party because in that particular trial the Jay was a sole attracted bird and the Blackcap focal pair showed only a weak calling response). Thus, there seems to be very low immediate costs of conspicuous mobbing to Blackcaps apart from time and energy expenditures. However, Krama and Krams (2005) showed experimentally that loud mobbing may inform eavesdropping predators about the location of the nest with the predation event taking place at a later time.
The attraction of neighbouring birds may also benefit nest owners as some of the attracted birds alarm called and even attacked the dummy, thus slightly increasing the intensity of nest defence in favour of the nest owners. The limitation of the current study to test this is the use of stuffed dummies, which cannot, in contrast to live intruders, be forced to leave the vicinity of the tested nest (Flasskamp 1994).
Nevertheless, potential benefits of predator distraction would probably be very low for Blackcaps under natural conditions, as the attracted birds participated infrequently both in active and passive nest defence (at 1 and 24% of the nests, respectively). However, according to the “risk dilution” and the “confusion” hypotheses, even the mere presence of more birds may be helpful to the focal pair. Under an assumption that Blackcaps do not confuse the Cuckoo with the Sparrowhawk (see above), one may suggest that these hypotheses can hardly apply in the case of the Cuckoo (in contrast to interactions with predators of adults). First, laying female Cuckoos do not attack adult hosts (thus there is no “risk” to be diluted). Second, female Cuckoos may succeed in parasitising a host nest even when the nest owners attack the laying Cuckoo female physically by pecking (Wyllie 1975; see also Moksnes et al. 2000). Thus, it is hard to imagine how the inactive presence of more birds could have any “confusing” effect on the Cuckoo when much more aggressive behaviour is ignored by the parasite. The low frequency of neighbours’ responses, coupled with low documented success of even very aggressive behaviours to brood parasitic adults, suggest that benefits to nest owners of attracting neighbours are negligible if any. Therefore, the observed pattern of positive correlation between the nest defence intensity and the number of attracted birds may be, from nest owners’ point of view, best explained as a proximate by-product of attracted birds to the noise around defended nests. Universal structure of alarm calls is generally thought to explain such interspecific attractions (Högstedt 1983; Hurd 1996).
To summarise, the attraction of neighbouring birds per se by nest defending Blackcaps seems to be neither costly (e.g. no predator attacking Blackcap adults or nest was attracted) nor beneficial (see low activity of attracted neighbours) for the tested species. It is important to stress that energy output and foregone opportunities due to mobbing are costs of mobbing itself and not costs of attraction, i.e. there are no extra costs related solely to the attraction of neighbours per se. Thus, from the point of view of “calling for help”, the nest defence by the Blackcap seems to be selectively neutral. In other words, the only function of aggressive mobbing by focal pairs at their nests seems to be to drive away the intruder and not to attract any neighbours which may be unhelpful in the context of brood parasitism (Wyllie 1975).
Attracted birds and their suitability as Cuckoo hosts
Four of the attracted species that alarm called (Chaffinch, Blackbird, Robin and Blackcap) are all known to be occasionally parasitised by the Cuckoo (Moksnes and Røskaft 1995). However, the Nuthatch which was the only species attacking the dummy and the Great Tit that alarm called have never been reported as successful Cuckoo hosts and may safely be considered unsuitable for Cuckoo parasitism due to their hole-nesting habits (Moksnes and Røskaft 1995). In contrast, no Chiffchaff (Phylloscopus collybita), Icterine Warbler (Hippolais icterina) or Whitethroat (Sylvia communis) ever participated in mobbing despite being in the vicinity and being suitable Cuckoo hosts. Even Cuckoo hosts known to show highly aggressive responses to Cuckoo dummies at their own nests (e.g. Phylloscopus warblers, Yellowhammer Emberiza citrinella, Blackbird, Chaffinch, Blackcap; Grim 2005a; Grim and Honza 2001; Røskaft et al. 2002; own unpublished data) did not physically attack the dummy near alien nests. This provides additional evidence against “calling for help” hypothesis (if we assume that the Cuckoo was not mistaken for the Sparrowhawk). The very low observed activity of suitable hosts near tested nests may in principle be explained by the “nesting-cue” hypothesis, which suggests that brood parasites use nest-defence responses directed towards them by hosts as cues to locate nests to parasitise (Gill et al. 1997). However, this is unlikely to apply for the current study because attracted potential hosts would be aggressive near alien but not their own nests.
In general, there were no differences between suitable and unsuitable Cuckoo hosts in abundance, frequency or participation in mobbing (Table 1). This may result from two reasons: (1) attracted birds did not recognize the Cuckoo as a special threat and ignored it (acceptance errors, sensu Sherman et al. 1997), or (2) attracted birds simply observed Blackcaps' nest defence to gain information about possible predators/parasites in their territories overlapping with Blackcap territories, and did not participate in nest defence at alien nests which would provide them with no benefits but would be costly in terms of energy and potential injuries. As the alarm rate by Blackcaps was strongly positively correlated with some of their risk-taking behaviours (attacks, time spent <1 m from the dummy, both P < 0.0001), the alarm calling in the Blackcap is obviously directed at the intruder near the nest and not at any secondary predators, kin or reciprocal altruists.
In summary, results of the present study support neither the “attract the mightier” (Högstedt 1983) nor “calling for help” (Rohwer et al. 1976) hypotheses. However, this does not preclude the adaptiveness of neighbour attraction in other species (Pavel and Bures 2001; Krams and Krama 2002) or accruance of benefits (e.g. information on presence of predators and parasites) for attracted birds themselves. The question of whether mobbing of a brood parasite attracts primarily suitable hosts and affects their behaviour would be a fruitful area for future research, especially in hosts uttering functionally referential alarm calls denoting brood parasites (Gill and Sealy 2004).
Zusammenfassung
Rufen Mönchsgrasmücken (Sylvia atricapilla), die ihr Nest verteidigen, auch ihre Nachbarn zu Hilfe?
Es wird vermutet, dass bei Vögeln eine Funktion des auffälligen Mobbens von Eindringlingen durch Nestbesitzer darin besteht, Nachbarn des Nestbesitzers („Hilferuf-Hypothese”) oder Fressfeinde des Eindringlings („Anlocken-des-Stärkeren-Hypothese”) anzulocken. Diese könnten dem Nestbesitzer durch Ablenken und/oder Attackieren des gemobbten Eindringlings zu Hilfe kommen. Bislang wurden diese Hypothesen lediglich beim Mobben von Prädatoren getestet; hier habe ich erstmals das Anlocken Dritter durch Mobbing im Zusammenhang mit Brutparasitismus untersucht. Ich habe ein Experiment mit Mönchsgrasmücken (Sylvia atricapilla) durchgeführt, einem kleinen Sperlingsvogel mit höchst aggressivem und auffälligem Nestverteidigungsverhalten. Ich habe aggressive Reaktionen der Mönchsgrasmücken durch das Präsentieren von ausgestopften Attrappen des brutparasitischen Kuckucks (Cuculus canorus) beziehungsweise von Kontrollen in der Nähe ihrer Nester hervorgerufen. Bei 32% der Nester (n = 75) lockten die Reaktionen der Mönchsgrasmücken auf die Attrappen bis zu 15 Vögel pro Versuch an, die insgesamt 21 verschiedenen Sperlingsvogelarten angehörten. Die meisten der angelockten Vögel waren artfremd und beteiligten sich kaum am Mobbing, d.h. die „Hilferuf-Hypothese” wurde nicht unterstützt. Es wurden keine potentiellen Prädatoren des Kuckucks angelockt, obwohl diese im Untersuchungsgebiet vorkamen und obwohl die Mönchsgrasmücken anhaltend mobbten, d.h. auch die „Anlocken-des-Stärkeren-Hypothese” wurde zurückgewiesen. Ich vertrete die Auffassung, dass diese Hypothese wahrscheinlich nicht auf typische Vogel-Prädatoren während Nestprädationsereignissen zutrifft, da diese nur wenige Sekunden dauern. Die Anzahl angelockter Vögel korrelierte positiv mit der Intensität, mit welcher der Besitzer sein Nest verteidigte, gemessen als Rate der Alarmrufe, nicht als visuelle Signale (Rate der Angriffe). Arten, die als Wirt für den Kuckuck geeignet waren, unterschieden sich in ihrem Verhalten in der Nähe verteidigter Nester nicht von als Wirt ungeeigneten Arten. Die beobachtete positive Korrelation von Nestverteidigungsintensität und Anzahl angelockter Vögel ist sehr wahrscheinlich ein proximates Nebenprodukt der auffälligen Nestverteidigung bei Mönchsgrasmücken (das Anlocken von Nachbarn an sich könnte jedoch sehr wohl adaptiv sein). Folglich ist das Mobbingverhalten der Mönchsgrasmücke gegen den Brutparasiten selbst gerichtet und nicht an Fressfeinde des Brutparasiten oder potentielle Rekruten für den Mob.
References
Bourne WRP (1977) The function of mobbing. Br Birds 70:266–267
Brooke M de L, Davies NB, Noble DG (1998) Rapid decline of host defences in response to reduced cuckoo parasitism: behavioural flexibility of reed warblers in a changing world. Proc R Soc Lond B 265:1277–1282
Bures S, Pavel V (2003) Do birds behave in order to avoid disclosing their nest site? Bird Study 50:73–77
Caro T (2005) Antipredator defenses in birds and mammals. University of Chicago Press, Chicago
Curio E (1978) The adaptive significance of avian mobbing I. Teleonomic hypotheses and predictions. Z Tierpsychol 48:175–183
Curio E (1993) Proximate and developmental aspects of antipredator behavior. Adv Stud Behav 22:135–238
Curio E, Klump G, Regelmann K (1983) An anti-predator response in the great tit (Parus major): is it tuned to predator risk? Oecologia 60:83–88
Davies NB, Madden JR, Butchart SHM (2004) Learning fine-tunes a specific response of nestlings to the parental alarm calls of their own species. Proc R Soc Lond B 271:2297–2304
Duckworth JW (1991) Responses of breeding reed warblers Acrocephalus scirpaceus to mounts of sparrowhawk Accipiter nisus, Cuckoo Cuculus canorus and jay Garrulus glandarius. Ibis 133:68–74
Flasskamp A (1994) The adaptive significance of avian mobbing. V. An experimental test of the “move on” hypothesis. Ethology 96:322–333
Ghalambor CK, Martin TE (2001) Fecundity-survival trade-offs and parental risk-taking in birds. Science 292:494–497
Gill SA, Sealy SG (2004) Functional reference in an alarm signal given during nest defence: seet calls of yellow warblers denote brood-parasitic brown-headed cowbirds. Behav Ecol Sociobiol 56:71–80
Gill SA, Grieef PM, Staib LM, Sealy SG (1997) Does nest defence deter or facilitate cowbird parasitism? A test of the nesting-cue hypothesis. Ethology 103:56–71
Grim T (2005a) Host recognition of brood parasites: Implications for methodology in studies of enemy recognition. Auk 122:530–543
Grim T (2005b) Mimicry vs. similarity: which resemblances between brood parasites and their hosts are mimetic and which are not? Biol J Linn Soc 84:69–78
Grim T (2006) Cuckoo growth performance in parasitized and unused hosts: not only host size matters. Behav Ecol Sociobiol 60:716–723
Grim T, Honza M (2001) Differences in behaviour of closely related thrushes (Turdus philomelos and T. merula) to experimental parasitism by the common cuckoo Cuculus canorus. Biologia 56:549–556
Halupka K, Halupka L (1997) The influence of reproductive season stage on nest defence by meadow pipits (Anthus pratensis). Ethol Ecol Evol 9:89–98
Högstedt G (1983) Adaptation unto death: function of fear screams. Am Nat 121:562–570
Honza M, Grim T, Capek M, Moksnes A, Røskaft E (2004a) Nest defence, enemy recognition and nest inspection behaviour of experimentally parasitised reed warblers Acrocephalus scirpaceus. Bird Study 51:256–263
Honza M, Prochazka P, Stokke BG, Moksnes A, Røskaft E, Capek M, Mrlik V (2004b) Are blackcaps current winners in the evolutionary struggle against the common cuckoo? J Ethol 22:175–180
Honza M, Polacikova L, Prochazka P (2007) Ultraviolet and green parts of the colour spectrum affect egg rejection in the song thrush (Turdus philomelos). Biol J Linn Soc 92 (in press)
Hurd CR (1996) Interspecific attraction to the mobbing calls of black-capped chickadees (Parus atricapillus). Behav Ecol Sociobiol 38:287–292
Kamil AC (1988) Experimental design in ornithology. In: Johnston RF (ed) Current ornithology, vol 5. Plenum, New York, pp 313–346
Klump GM, Curio E (1983) Reactions of blue tits Parus caeruleus to hawk models of different sizes. Bird Behav 4:78–81
Krama T, Krams I (2005) Cost of mobbing call to breeding pied flycatcher, Ficedula hypoleuca. Behav Ecol 16:37–40
Krams I, Krama T (2002) Interspecific reciprocity explains mobbing behaviour of the breeding chaffinches, Fringilla coelebs. Proc R Soc Lond B 269:2345–2350
Krams I, Krama T, Igaune K (2006) Mobbing behaviour: reciprocity-based co-operation in breeding pied flycatchers Ficedula hypoleuca. Ibis 148:50–54
Moksnes A, Røskaft E (1992) Responses of some rare cuckoo hosts to mimetic model cuckoo eggs and to foreign conspecific eggs. Ornis Scand 32:17–23
Moksnes A, Røskaft E (1995) Egg-morphs and host preference in the common cuckoo Cuculus canorus: an analysis of cuckoo and host eggs from European museum collections. J Zool 236:625–648
Moksnes A, Røskaft E, Hagen LG, Honza M, Mork C, Olsen PH (2000) Common cuckoo Cuculus canorus and host behaviour at reed warbler Acrocephalus scirpaceus nests. Ibis 142:247–258
Montgomerie RD, Weatherhead PJ (1988) Risks and rewards of nest defence by parent birds. Q Rev Biol 63:167–187
Pavel V, Bures S (2001) Offspring age and nest defence: test of the feedback hypothesis in the meadow pipit. Anim Behav 61:297–303
Payne RB, Payne LL, Rowley I (1985) Splendid wren Malurus splendens response to cuckoos: an experimental test of social organization in a communal bird. Behaviour 94:108–127
Rohwer S, Frettwell SD, Tuckfield RC (1976) Distress screams as a measure of kinship in birds. Am Midl Nat 96:418–430
Røskaft E, Moksnes A, Stokke BG, Bicik V, Moskat C (2002) Aggression to dummy cuckoos by potential European cuckoo hosts. Behaviour 139:613–628
Rothstein SI (2001) Relic behaviours, coevolution and the retention versus loss of host defences after episodes of avian brood parasitism. Anim Behav 61:95–107
Rytkönen S, Soppela M (1995) Vicinity of sparrowhawk nest affects willow tit nest defence. Condor 97:1074–1078
SAS Institute (2004) SAS online doc. SAS Institute, Cary
Schaefer T (2004) Video monitoring of shrub-nests reveals nest predators. Bird Study 51:170–177
Sealy SG, Neudorf DL, Hobson KA, Gill SA (1998) Nest defense by potential hosts of the brown-headed cowbird: methodological approaches, benefits of defence, and coevolution. In: Rothstein SI, Robinson SK (eds) Parasitic birds and their hosts, Oxford University Press, New York, pp 194–211
Sherman PW, Reeve HK, Pfenning DW (1997) Recognition systems. In: Krebs JR, Davies NB (eds) Behavioural ecology, Blackwell, Oxford, pp 69–96
Shirihai H, Gargallo G, Helbig AJ (2001) Sylvia warblers. Princeton University Press, Princeton
Smith S, Hosking E (1955) Birds fighting. Experimental studies of the aggressive displays of some birds. Faber and Faber, London
Soler M, Martin-Vivaldi M, Perez-Contreras T (2002) Identification of the sex responsible for recognition and the method of ejection of parasitic eggs in some potential common cuckoo hosts. Ethology 108:1093–1101
Veen T, Richardson DS, Blaakmeer K, Komdeur J (2000) Experimental evidence for innate predator recognition in the Seychelles warbler. Proc R Soc Lond B 267:2253–2258
Winkler DW (1994) Antipredator defense by neighbors as a responsive amplifier of parental defense in tree swallows. Anim Behav 47:595–605
Wyllie I (1975) Study of cuckoos and reed warblers. Br Birds 68:369–378
Yasukawa K (1989) The costs and benefits of a vocal signal: the nest-associated ‘chit’ of the female red-winged blackbirds, Agelaius phoeniceus. Anim Behav 38:866–874
Yom-Tov Y (2001) An updated list and some comments on the occurrence of intraspecific nest parasitism in birds. Ibis 143:133–143
Acknowledgments
I thank two anonymous referees, S. A. Gill, M. E. Hauber, I. Krams, V. Pavel, and V. Remeš for comments on the draft. I am especially grateful to E. Curio for his extensive and insightful comments and to V. Pavel for helping me with the literature. I am indebted to M. Čapek for providing breeding density data and to D. Campbell for correcting the language. I was supported by grants MSM6198959212 and GACR 206/03/D234.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by T. Friedl.
Rights and permissions
About this article
Cite this article
Grim, T. Are Blackcaps (Sylvia atricapilla) defending their nests also calling for help from their neighbours?. J Ornithol 149, 169–180 (2008). https://doi.org/10.1007/s10336-007-0257-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-007-0257-7