
Abstract
We analysed female body mass change, corrected by tarsus length (body condition) in tawny pipits Anthus campestris during the nesting period in a population subject to high nest predation rates (between 70 and 85%), which leads to the need for replacement clutches. Decrease in female body condition over the nesting stage (6.8 g, around 27% of the initial mass during the whole nesting process) was related to laying date, clutch size and nesting period (incubation and nestling phases). Data from recaptured females indicated a decrease during each of the three nesting phases considered (the last days of incubation and first and last days of the nestling phase), with body mass always being higher in the first of the two measurements taken in each of these phases. The observation of a continuous decrease in body condition during the last days of incubation and first and last days of the nestling phase does not support the programmed anorexia hypothesis, but adjusts well to predictions of the stress hypothesis. These results suggest that the costs accumulated during the entire nesting stage in ground passerines subjected to high nest predation rates are linked to a superimposed effect of the cost of replacement clutches.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Body mass change during the nesting period has been considered an indicator of the reproductive stress suffered by the adults. Female mass loss, which is usually observed during the reproductive period, has been positively associated with clutch size and therefore with the cost of reproduction (stress hypothesis; Hussell 1972; Ricklefs 1974; Bryant 1979; Moe et al. 2002). Nevertheless, it has been suggested that the mass loss observed in the post-hatching period could be programmed, i.e. adaptive (Freed 1981; Norberg 1981; Moreno 1989a; Slagsvold and Johansen 1998; Cichon 2001), given that it would confer advantages to the adults in reducing the costs of flying when feeding young (programmed anorexia hypothesis, in the terminology of Jones 1994). The two hypotheses are not mutually exclusive, given that mass loss could be programmed, but, below a certain threshold, could still indicate reproductive stress (Jones 1994), especially if the high energy cost to females of incubation and nestling care is taken into account (Monaghan and Nager 1997; Tinbergen and Williams 2002, and references therein).
The timing of mass loss determines which of the two hypotheses best applies (Croll et al. 1991; Jones 1994). In the programmed anorexia hypothesis the mass losses have to occur before the period of maximum energy demand (Moe et al. 2002). This is during the nestling and chick-rearing period (see, however, Williams and Dwinnel 2001) in those passerines with monoparental incubation. According to the programmed anorexia hypothesis, mass losses would occur at the end of the incubation period or just at the beginning of the nestling one (Gaston and Jones 1989, Phillips and Furness 1997). In contrast, under the stress hypothesis, the mass losses would occur throughout the whole nestling period, from the first day to the last (Weldeln and Becker 1996), although some mass loss during the final days of incubation might also be observed, because of the high energy costs incurred during this period (Monaghan and Nager 1997; Williams and Dwinnel 2001; Tinbergen and Williams 2002).
We explored two basic ideas:
-
1.
Is the pattern of change of body condition (body mass corrected by tarsus length) of female tawny pipits Anthus campestris best explained by the anorexia hypothesis or the stress hypothesis? We expect that if the observed loss of body condition fits the anorexia hypothesis, the pattern during the last days of the incubation period or the first days of nestling phase will show a decrease in one or both of the two periods. Also, it will not involve a mass loss during the final days of the nestling period. If the loss pattern fits the stress hypothesis there will be a decrease in female body mass at the end of the incubation period and throughout the nestling one.
-
2.
Does the laying date affect the body mass condition of the females? The nests in our population are subjected to high predation rates (between 70 and 85%, Suárez et al. 1993), which lead to a need for replacement clutches in a species, which normally lays only one or two clutches (Cramp 1988). Therefore laying date must be positively correlated with the number of replacement clutches in the population and it would also be expected that body condition would be negatively correlated with laying date, if a cost associated with the number of clutches exists.
Methods
The species
The tawny pipit is a medium sized passerine (females: 25 g) whose natural history traits are summarised in Cramp (1988). The species is ground-nesting, and in our population only females incubate and feed nestlings (J. Herranz, unpublished data). Fledglings are semiprecocial and are cared for by both sexes (Cramp 1988). The potential number of replacement clutches is unknown.
Study area and methods
The study area was located in central Spain (Layna, Soria, 41°05′N, 1°50′W; 1,200 m a.s.l.). It has a continental Mediterranean climate, with an annual rainfall of 500 mm and an annual mean temperature of 10.2°C. The landscape is flat or gently undulating and the vegetation is a shrub-steppe dominated by Genista pumila, G. scorpius, Thymus spp., Poa spp. and Stipa spp.
Nests were found through systematic searches and by observations of adult behaviour during May–June 1995. The earliest laying date registered was 18 May, the modal date falling within the period 27–31 May (dates grouped by 5-day classes). Laying dates of successive or second clutches continued until 4 July (n=43 nests). The females were caught at the nest during incubation (early incubation, age of the nest 1–6 days, n=4 different females; late incubation, 7–12 days, n=17) and/or when feeding chicks in the nest (early nestling phase, age of the nest 13–18 days, n=15; late nestling phase, >18 days, n=17; total number of different females caught, n=43). Traps were only used for 30 min a day to avoid excessive disturbance at the nest. Birds were colour-ringed and sometimes recaptured. Individuals were weighed on an electronic balance with a precision of 0.01 g, and tarsal lengths were measured with callipers to the nearest 0.01 mm.
Clutch sizes and dates of first eggs were recorded for each nest. For nests discovered during incubation, clutch size was taken as the maximum number of eggs found. For nests found during the nestling period, a clutch size equal to the maximum number of chicks recorded was assumed. The date of the first egg laid was estimated assuming a laying rate of 1 per day (Cramp 1988). In nests found during the post-hatching stage, the laying date was estimated assuming a 12-day incubation period and using growth curves of the chicks, with an estimated error of ±1 day (e.g., Shkedy and Safriel 1992). Nest mortality in our population during the study years was 74.9%, estimated by means of the Mayfield method (Mayfield 1961).
Data analysis
Two separate analyses were made, one using data from all birds combined and the second using only those birds which were recaptured. In the first, we used General Linear Models (GLMs), using the body mass of individuals as the dependent variable (n=43). To avoid pseudoreplication problems resulting from measuring individuals on successive days, we used average mass and average capture dates, as long as the data related to the same phase of nesting (incubation or the nestling phase). Clutch size and nesting period were considered as fixed factors and laying date and nest age (taken as the number of days from the first laying date in the population and the number of days for which each nest was occupied, respectively) as covariates. We used the quadratic term of both covariates because it was more explanatory than the linear one. To eliminate possible effects resulting from individual differences in body mass, tarsus length was also included in the analyses as a covariate (Freckleton 2002).
A second analysis was undertaken on the variation in body mass of recaptured females (n=17) during different phases of nesting. We defined four phases: (1) first 6 days of incubation, (2) last 6 days of incubation, (3) first 6 days of the nestling period, and (4) the nestling period after the 6th day. We have used the first and last mass measurement of recaptured females taken in each phase, except for phase (1), where data were lacking. Data for females captured only once during each phase were not considered. We used a repeated measures ANOVA test, with the first and last body mass measurements in each phase as dependent variables and the nesting phase as a fixed factor. The number of days between captures and the quadratic term of laying date were included as covariates, as well as the individual tarsal lengths.
To determine whether there was any change in the body mass loss of recaptured females between nesting phases we analysed the daily body mass change (first minus last measurements divided by the number of intervening days) using a one-way ANOVA. We used the quadratic term of the laying dates, the number of days elapsed between measurements and the tarsal lengths as covariates.
In all GLM analysis we always tested the two-way interaction terms and removed them from the model when insignificant. The number of females in each test is given in the text. The significance level considered was P=0.05. All analyses were undertaken using the STATISTICA 6.0 statistical package (StatSoft 2002).
Results
The females showed a decrease in body condition throughout the breeding period with respect to the quadratic term of the laying date, nesting period and clutch size (Table 1). Body mass decreased by approximately 2 g between the first and last dates (Fig. 1a), being greater during incubation than during the nestling period (Fig. 1b). Female body mass also decreased as clutch size increased (Fig. 1c). Nest age was not significant.

Female tawny pipit Anthus campestris body mass variation in relation to a laying date, showing the line from a curvilinear regression only as mass trend, b nesting period (marginal mean±SE) and c clutch size (marginal mean±SE). Marginal means come from the residuals of the model, once corrected by the covariates
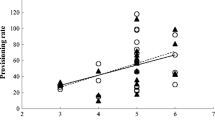
When considering recaptured females, the vast majority lost mass during the whole nesting period. The great majority lost body mass during the final days of the incubation period (83.3%, n=6). Nearly all did so during the first days of the nestling phase (88.9%, n=9) and all lost mass during the final days of that phase (100%, n=10). Female body mass varied significantly between the first and the last days within each phase, and across the three phases (Table 2), body mass always being higher in the first of the two measurements taken in each phase (Fig. 2). The number of days elapsed between captures was not significant in any case (P>0.05), but the quadratic term of the laying date was significant for the final mass variable, showing that the longer the season progressed the more that female body condition at the end of each phase decreased.

Female body mass variation (mean±SE) between first and last measurements in the three nesting phases considered. Blank squares represent first measurements in each phase and blank triangles represent last measurements in each phase
Daily mass loss rates were not significantly different between nesting phases (phase 2: 0.17±0.26 g/day, n=6; phase 3: 0.24±0.20 g/day, n=9; phase 4: 0.37±0.32 g/day, n=10; one-way ANOVA, df=2; F=1.476; P=0.252). Given these daily mass losses and the duration of the two nesting periods (12 days incubation and 12 days of nestling period), females lose 2.8 g during incubation and 4.0 g during the post-hatching phase, representing an average total of 27% of the female body mass during the nesting period.
Discussion
The results of the first GLM analysis showed that the quadratic term of laying date, clutch size and nesting period are determinant variables in the variation of body condition. Also, the fact that the recaptured females lost body condition significantly during the three nesting phases considered shows that mass loss in females is related to nest age. All these results support the stress hypothesis, showing the cost to female body condition being associated with the number of clutches laid.
Although the total number of replacement clutches laid is unknown, females in the studied population may extend their nesting period until 1.5 months after the first laying date (extreme laying dates recorded were 18 May and 4 July; Fig. 1). This fact, together with the high nest predation rates recorded (nearly 75% of the nests), suggests that laying date may be indicative of successive clutches being laid and that the decrease in body condition is linked to the high energetic costs of laying the previous clutches, which must be elevated in a species with monoparental incubation, brooding and nestling feeding. The quadratic term of the laying date seems to indicate that the body condition decrease is sharper as the season progresses, which was also observed in the results of the analysis of body masses of recaptured females.
The continuous decline in body condition of the recaptured females during the final days of incubation and first and last days of the nestling period also fits the stress hypothesis predictions well. Moreover, all females lost a high proportion (approx. 4%) of their mass during the last phase of the nestling period, which cannot easily be explained as programmed anorexia. The total mass lost over the whole nesting period (27%) is also indicative of the application of the stress hypothesis, especially since the mean mass losses estimated for a range of other passerines was approximately 10% (Moreno 1989b). This fact indicates that the cost (in body mass terms) of the entire nesting period is high and perhaps does not allow female tawny pipits to recover their body condition after their first clutches and it agrees with observations of the energetic costs incurred by females of various species during the incubation and nestling periods (Tinbergen and Williams 2002).
The pattern in which mass loss increases with clutch size, so that maximum female body masses correspond with the smallest clutch sizes, is also compatible with the stress hypothesis. According to this hypothesis, the larger the clutch size the greater the energy expenditure of the females involved (Monaghan and Nager 1997), with a knock-on effect on body condition when females produce replacement clutches. Besides, and especially where only one parent incubates, the energy demands of incubation may be higher as clutch size increases (see review in Monaghan and Nager 1997). However, this relationship is not always so obvious, given that uncontrolled underlying factors such as female age or territory quality may cause phenotypic correlations between traits (Partridge and Harley 1985; Reznick 1985).
The two models of body mass changes during nesting proposed by Moreno (1989b) are not supported by our data from recaptured females. The mass losses observed during incubation and during the final stages of the nestling phase do not fit the incubatory mass constancy model, in which females theoretically maintain their body masses during incubation, losing it later during the nestling period and with some degree of recovery at the end of that stage. The body mass losses observed during the nestling period also do not fit the incubatory mass loss model, in which the females would lose body mass during incubation and then maintain it during the post-hatching period. However, as suggested by the same author, a continuous loss of female body mass during both nestling phases may be observed in poor environments providing only a low quality or quantity of feeding resources, which applies to our data. Similar results have been found for thekla larks Galerida theklae and lesser short-toed larks Calandrella rufescens in a semi-arid habitat (Suárez et al. 2005).
Ground passerines, such as the tawny pipit, whose movements and foraging for their nestlings and fledglings are basically terrestrial, have a lower apparent advantage in losing mass to reduce flying demands than forest or marine species, which may spend a lot of time flying when travelling or feeding (e.g. Jones 1994; Kullberg et al. 2002). Data which support the programmed anorexia hypothesis have been accumulated for these latter species (see, e.g., Croll et al. 1991; although see also Moe et al. 2002). However, our results, and the absence of further data on ground-nesting passerines subjected to high nest predation rates, do not allow us to extend its validity to such species. Instead, the existence of lower flying demands combined with the accumulated cost of raising successive clutches, seem to show that the stress hypothesis is more applicable to terrestrial passerines.
Zusammenfassung
Änderung der Körpermasse bei weiblichen Brachpiepern (Anthus campestris) während des Nistens
Wir analysierten die Änderung der Körpermasse von weiblichen Brachpiepern über die Nistzeit in einer Population mit hohen Nestpredationsraten (70–85%), die Nachbruten nötig machten. Die Körpermasse wurde an der Tarsuslänge normiert und als Körperverfassung bezeichnet. Eine Verschlechterung der Körperverfassung der Weibchen (6.8 g, etwa 27% des Ausgangsgewichts über die gesamte Nistzeit) stand in Beziehung zum Legedatum, der Gelegegröße und der Nistphase (Brut- und Nestlingsphase). Daten von wieder gefangenen Weibchen zeigten eine Verschlechterung während jeder der drei betrachteten Nistphasen (die letzten Tage des Brütens und die ersten und letzten Tage der Nestlingsphase), wobei die Körpermasse für die jeweils erste von zwei Messungen in jeder dieser. Phasen höher war. Der Befund einer sich stetig verschlechternden Körperverfassung über diese Phasen unterstützt nicht die Hypothese der programmierten Anorexie, sondern passt gut auf die Stress-Hypothese. Diese Ergebnisse legen nahe, dass die akkumulierten Kosten, die bodenbewohnenden Singvögeln unter einem hohen Nestpredationsdruck während der gesamten Nistzeit entstehen, verknüpft sind mit einem überlagernden Effekt der Kosten für Nachbruten.
References
Bryant DM (1979) Reproductive cost in the house martin Delichon urbica. J Anim Ecol 48:655–676
Cichon M (2001) Body-mass changes in female collared flycatchers: state-dependent strategy. Auk 118:550–552
Cramp S (ed) (1988) The birds of the western Palearctic, vol 5. Oxford University Press, Oxford
Croll DA, Gaston AJ, Noble DG (1991) Adaptive loss of mass in thick-billed murres. Condor 93:496–502
Freckleton R (2002) On the misuse of residuals in ecology: regression of residuals vs. multiple regression. J Anim Ecol 71:722
Freed LA (1981) Loss of mass in breeding wrens: stress or adaptation? Ecology 62:1179–1186
Gaston AJ, Jones IL (1989) The relative importance of stress and programmed anorexia in determining mass loss by incubating Ancient Murrelets. Auk 106:653–658
Hussell DTJ (1972) Factors affecting clutch size in artic passerines. Ecol Monogr 42:317–364
Jones IL (1994) Mass changes of least auklets Aethia pusilla during the breeding season: evidence for programmed loss of mass. J Anim Ecol 63:71–78
Kullberg C, Houston DC, Metcalfe VB (2002) Impaired flight ability—a cost of reproduction in female blue tits. Behav Ecol 13:575
Mayfield H (1961) Nesting success calculated from exposure. Wilson Bull 73:255–261
Moe B, Langseth I, Fyhn M, Gabrielsen GW, Bech C (2002) Changes in body condition in breeding Kittiwakes Rissa tridactyla. J Avian Biol 33:225–234
Monaghan P, Nager RG (1997) Why don’t birds lay more eggs? Trends Ecol Evol 12:270–274
Moreno J (1989a) Body-mass variation in breeding Northern Wheatears: a field experiment with supplementary food. Condor 91:178–186
Moreno J (1989b) Strategies of mass change in breeding birds. Biol J Linn Soc 37:297–310
Norberg RA (1981) Temporary weight decrease in breeding birds may result in more fledged young. Am Nat 118:838–850
Partridge L, Harvey P H (1985) Costs of reproduction. Nature 316:20
Phillips RA, Furness RW (1997) Sex specific variation in the loss of mass by breeding Arctic skuas. J Avian Biol 28:163–170
Reznick D (1985) Costs of reproduction: an evaluation of the empirical evidence. Oikos 44:257–267
Ricklefs RE (1974) Energetics of reproduction in birds. In: Paynter R (ed) Avian energetics. Nuttall Ornithological Club, Cambridge, Mass., pp152–297
Shkedy Y, Safriel UN (1992) Nest predation and nestling growth rate of two lark species in the Negev Desert, Israel. Ibis 134:268–272
Slagsvold T, Johansen MA (1998) Mass loss in female Pied Flycatchers Ficedula hypoleuca during late incubation: supplementation fails to support the reproductive stress hypothesis. Ardea 86:203–212
StatSoft (2002) STATISTICA (data analysis software system), Version 6. www.statsoft.com
Suárez F, Yanes M, Herranz J, Manrique J (1993) Nature reserves and the conservation of Iberian shrubsteppe passerines: the paradox of nest predation. Biol Con 63:77–81
Suárez F, Sánchez AM, Herranz J, Traba J, Yanes M (2005) Parental body mass changes during the nesting stage in two lark species in a semi-arid habitat. J Arid Env (In press)
Tinbergen JM, Williams JB (2002) Energetics of incubation. In: Dreeming DC (ed) Avian incubation: behaviour, environment, and evolution. Oxford University Press, Oxford, pp 298–313
Weldeln H, Becker PH (1996) Body mass change in breeding Common Terns (Sterna hirundo). Bird Study 43:85–95
Williams JB, Dwinnel B (2001) Field metabolism of free-living Savannah Sparrow during incubation: a study using doubly labeled water. Physiol Zool 63:353–372
Acknowledgements
We wish to thank three anonymous referees whose comments helped us to improve the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by R. Hoberton
Rights and permissions
About this article
Cite this article
Suárez, F., Traba, J. & Herranz, J. Body mass changes in female tawny pipits Anthus campestris during the nesting stage. J Ornithol 146, 372–376 (2005). https://doi.org/10.1007/s10336-005-0092-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-005-0092-7