
Abstract
In 1995 to 2000, site-fidelity of colour-marked Nordic greylag geese (Anser anser) was examined during spring migration in the Rheiderland, an important stopover site in NW Germany. The results demonstrated that site fidelity in greylag geese depends very much on the mating status of the geese. Paired birds showed a significantly higher site fidelity than unpaired adults. There was no difference in the length of the roosting period between the two social groups. The year-to-year return rate of individuals showed significant differences in relation to the individual breeding success. Birds with high numbers of young showed a significantly higher return rate than birds with low numbers of young.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Every year, greylag geese (Anser anser) migrate along the North Sea coast to their wintering grounds in Spain. From there, they start their return migration in February. They stop from March to April in the Netherlands and along the German North Sea coast, before returning to their northern breeding grounds in May (Andersson et al. 2001). Ringleben (1957) reported that “geese and their offspring use the same roosting sites every year”. Based on 8 years of observations of marked greylag geese at a spring staging site, site-fidelity of individuals to particular feeding areas has been investigated in relation to the social status of the focal birds.
Site fidelity, particularly at breeding sites, has been observed in a variety of waterbirds. Site fidelity has also been demonstrated for waterfowl on staging sites during their annual migration. Geese and swans are well known examples (Greenwood and Harvey 1982), e.g. greylags on autumn migration (Nilsson and Persson 1992) and on the wintering grounds. Site fidelity has also been noted for other species, such as the Greenland white-fronted goose (A. albifrons flavirostris; Fox et al. 2002), bean goose (A. fabalis; Nilsson and Persson 1991), barnacle goose (Branta leucopsis; Ganter 1994) and brent goose (B. b. bernicla; Prokosch 1984). Rees and Bacon (1996) noted high site fidelity in marked Bewick’s swans (Cygnus bewickii) during the entire winter period. Unpaired individuals changed sites more often than paired birds with families. Ganter (1994) found mobile and sedentary barnacle geese in the Wadden Sea, but could not discover any social or age dependent patterns. Marked individuals of Greenland whitefronts stop over in Iceland on their migration from Greenland to Scotland or Ireland. Fox et al. (2002) demonstrated that 90% of juvenile birds use feeding sites within a circle of 4 km jointly with their parents and siblings during spring migration. High site fidelity has been similarly proven for the pink-footed goose (A. brachyrhynchus) (Fox et al. 1994). Some studies show that site fidelity is age related (Wilson et al. 1991; Warren et al. 1992). Using observations of colour-marked greylag geese from the Ems estuary (NW Germany), we examined both the site fidelity of spring staging and possible social mechanisms.
Material and methods
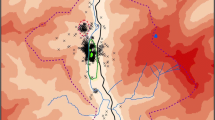
The investigations were carried out in the pasture areas of the Rheiderland and the Moormerland (Landkreis Leer, Lower Saxony; Fig. 1). The feeding areas of greylags are situated near the river Ems or close to the Dollard dike, as well as along the larger water bodies in the study site area (see Kruckenberg et al. 1996; Gerdes 2000; Borbach-Jaene et al. 2002). The total size of the area is about 170 km2.

Study area with ringing sites of Nordic greylag geese (Anser anser)
In the winters of 1994/1995, 1996/1997 and 1997/1998, the study site was surveyed every second day and all marked birds were recorded (Kruckenberg et al. 1996; Borbach-Jaene et al. 2001).
Only geese with blue neck collars of the Scandinavian goose ringing project recorded during spring migration have been included in the analyses. The marked birds are all of the north Norwegian subpopulation.
Whenever feasible, the social status (paired or unpaired) and the number of offspring were noted for each ringed individual. The age of the marked birds has been generated from the ringing data. For birds with unknown year of birth a minimum age of 2 years has been assumed when ringed as adult.
The site fidelity of the geese to their feeding sites was investigated. Only those birds, which were observed more than 3 times in the study site (52 individuals, 550 observations) have been taken into account. The feeding areas of the geese have been included in a GIS database with precise data on the visited field and the time frame. The measure for site fidelity was defined by the average of the distances between locations including all observations of that particular individual in a given year. A special GIS-extension tool (Jenness 2001) has been used. At the same time, the number of days between sightings and the minimum duration of stay of the geese has been generated.
The analysis distinguished between juvenile (young birds of the last season), subadult (2 years) and adult (>2 years). The birds have also been separated in “stationary” (always in a very small area, not more than 4 km2) and “movers” (birds mobile in the study area) and compared to various different criteria (social status, age). Additional weekly observations, carried out in the same way as described above in the period 1998–2002 have been included (in total 81 individuals, 937 observations).
Results
Site fidelity to feeding areas
Figure 2 shows the average distance between each observation in regard to social status (n=52) in a box-plot demonstration. The results show that unpaired birds use larger areas than paired geese (Kruskal-Wallis test P>0,05) or juvenile birds. However, there is no difference in the mobility of mated adults and juvenile birds (U-test, P=88) which are roosting together with their families. There also appears to be no difference in the mobility between the age structures. Subadult birds show the highest mobility, but due to the low number of subadult birds (n=2) no statistical test could be carried out.

Distances between observation places during roosting time at Rheiderland of neck-banded greylags with social status (juvenile birds = 6, mated adults = 50, unmated adults = 11)
Duration of stopover
Figure 3 shows the staging duration of the focal individuals in the study area. The three social groups do not show any significant differences (Kruskal-Wallis test P=0.42).

Roost duration of neck-banded greylags at Rheiderland with known social status (n=67)
Stationary birds and movers
Figure 4 shows the classification of the stationary and non-stationary birds. Only birds with more than three observations have been included in the data set. The majority of stationary birds are paired adults. Only one unpaired adult and nine young birds were observed in this group. This is different for the group of movers: 11 out of 13 movers were unpaired adult birds or subadult. Only two birds in this category were paired. The differences between social status of stationary and mobile birds are highly significant (χ2=70.06, df=3, P<0.001).

Distribution of moving and stationary individuals in social groups during spring migration roost at Rheiderland
Feeding site fidelity depending on age
Figure 5 shows the age classes of stationary and mobile birds. It is important to note that the age classifications 3, (i.e. 4–10 years or >10 years of age) are often the minimum age of the birds, whereas the classifications of 1 and 2-year-old birds show the actual age. No young bird was found in the mobile group, but 25% of the mobile birds were 2 years old. 75% of the sedentary birds were older than 3 years. Differences between both groups in age are highly significant (n=92, χ2=30.38, df=4, P<0.001).

Age dependent distribution of mobile and stationary individuals during spring migration roost at Rheiderland
Site fidelity and rate of re-sighting
Figure 6 shows the return rate of site faithful individuals between years. Of the adult birds with a high site fidelity within one season, 34% did not return during another season. However, 18% returned 1 out of 7 and 6% 2 out of 7 years. The large majority of 58% returned to their known feeding areas in more than 3 years.

Return rates of stationary individuals at Rheiderland
We also examined whether differences in the breeding success between those returning only once or twice and those three and more times are dependent on age. Both groups have been tested. The differences between both group are not significantly dependant on age (t-test P<0.15).
The average number of juveniles of marked birds is shown in Table 1 for birds differing in their return rates. The higher the average number of juveniles the more likely it becomes that the birds return to the study area. There is a significant difference between the birds staging one or two seasons in the area and birds with two or more returns to the site involving juveniles still seen with their parents in spring (Mann-Whitney U-Test, P>0.05).
Discussion
Biological importance of traditional site use
On spring staging sites of Nordic greylag geese we found a traditional site use of mated adult greylag geese and their offspring. Unpaired birds were significantly less site faithful (Fig. 4), but this was not a result of differences in staging duration (Fig. 3). Site fidelity of mated birds on spring migration may cause a higher breeding success.
Wild geese carry out long distance migration every year. In order to acquire the necessary energy levels to carry out these flights, they need to feed in nutrient rich feeding areas. Because of the shortness of the arctic summer, the geese have to optimise their timing of fattening and migration (Madsen 2001). Staging sites during spring migration are very important in this respect. During spring staging, the resources are acquired for migration, egg laying, breeding and care of young. Small differences in the acquisition of spring body reserves can determine success or failure of breeding (Ebbinge 1989). For successful breeding it is important to optimise energy acquisition in spring.
Favourable feeding conditions in the wintering areas increase the breeding success of the female in the following season (Ankey and MacInnes 1978). Hence it can be profitable for paired birds, aiming at successful breeding for the coming summer, to use well-known sites for feeding. During the first winter, offspring migrate together with their parents (Bergmann et al. 1994) and it therefore comes as no surprise that site fidelity of offspring is similar to site fidelity of mated (parental) birds. In the second winter most of the subadult birds migrate on their own and do not show levels of site fidelity characteristic of their first winter behaviour (Fig. 5). They are more mobile and potentially develop their own knowledge of sites. When reaching breeding age the birds tend to be more sedentary again. During spring migration breeding pairs have to optimise their feeding behaviour. Non-breeders, and young or old birds without partners, may meet different objectives. They can probably afford to test new feeding areas or even unknown sites. In this way they may gain individual experience in preparation for future breeding and explore new roosting sites at the same time
Dominance and social structure
In geese, families dominate over single birds and big families dominate over smaller ones (Boyd 1953; Raveling 1970; Lamprecht 1986). Stahl et al. (2001a) confirmed similar observations in the brent goose with larger families dominating over smaller families or over pairs in spring on salt marshes. Pairs with many young showed a higher return rate than those with fewer young. The same has been confirmed for greylag geese in the Dollard region. A bird is more likely to return to the same site when more young have been produced. Family ties can prove to be advantageous when defending profitable established feeding sites against single birds or pairs without young. Subdominant birds are pushed towards suboptimal sites and hence their willingness to look for other sites may increase (Rees and Bacon 1996). On a small scale, subdominant birds show more exploratory behaviour and find new food sources earlier (Stahl et al. 2001b). These are good reasons for assuming that subdominant birds also explore new roosting sites and start new traditions.
Staging site fidelity over many years
In our study, individual greylags which have been observed in the area over many years are seen with more juveniles in spring than those pairs only observed once or twice in the study area. There are two possible explanations. Long-term site-faithful geese have a high breeding success. They know the area well and occupy optimal feeding sites. The feeding area has proved to be a good site for them in preparation for migration and they return every year. Those birds breeding less successfully switch to other areas to improve their spring feeding opportunities. Pink-footed geese which left spring staging areas in poor condition changed spring staging strategy in the next season; birds in good condition were less likely to change (Madsen 2001). So, the number of the returning geese could be an indirect measure for the quality of the site for staging and feeding on spring migration.
The other explanation refers to the successful winter survival of young geese for the heavily hunted greylag goose population rather than referring to the breeding success. Geese with high return rates could also be those which choose areas with low hunting pressure. In this case, the return rate of birds is more likely to express the ability of the parents to avoid predation and protect their young rather than the quality of the spring feeding area in terms of forage quality. Those birds using well-known areas use their site knowledge to avoid predation and hunting and thus enhance their survival rate (Robertson and Cooke 1999). The greylag goose “G01 blue” provides an example for this hypothesis. Every year it was seen in the Dutch province “Waterland Oost”, where goose damage to agricultural fields was prevented through increased hunting in 1992 (Visbeen and van der Waal 1997). Since 1993, G01 has been observed in the Dollard area. The goose is now returning to this area in spring ever since (more than 9 years). This shows that geese keep traditions as long as it is advantageous, but are able to abandon them when disadvantages start to prevail.
References
Andersson A, Follestad A, Nilsson L, Persson H (2001) Migration pattern of Nordic Greylag Geese Anser anser. Orn Svecica 11:19–58
Ankney CD, MacInnes CD (1978) Nutrient reserves and reproductive performance of female lesser snow geese. Auk 95:459-471
Bergmann H-H, ten Thoren B, Stock M (1994) Ringelgänse—Arktische Gäste an unseren Küsten. Aula, Wiesbaden
Borbach-Jaene J, Kruckenberg H, Lauenstein G, Südbeck P (2001) Arktische Gänse als Rastvögel im Rheiderland—Eine Studie zur Ökologie und zum Einfluss auf den Ertrag landwirtschaftlicher Kulturen. Landwirtschaftsverlag Weser-Ems, Oldenburg
Borbach-Jaene J, Kruckenberg H, Becker C (2002) Ergebnisse des Gänsemonitorings in der Ems-Dollart-Region 1996–2001. Vogelkd Ber Niedersachs 17:128–154
Boyd H (1953) On encounters between White-Fronted Geese in winter flocks. Behaviour 5:85–129
Ebbinge BS (1989) A multifactorial explanation for variation in breeding performance of Brent Geese Branta bernicla. Ibis 131:196–204
Fox AD, Mitchell C, Stewart A, Fletcher JD, Turner JVN, Boyd H, Shimmings P, Salmon DG, Haines WG, Tomlinson C (1994) Winter movements and site-fidelity of Pink-footed Geese Anser brachyrhynchus ringed in Britain, with particular emphasis on those marked in Lancashire. Bird Study 41:221–234
Fox AD, Hilmarsson JO, Einarsson O, Walsh AJ, Boyd H, Kristiansen JN (2002) Staging site fidelity of Greenland White-fronted Geese Anser albifrons flavirostris in Iceland. Bird Study 49:42–49
Ganter B (1994) Site tenacity and mobility of staging Barnacle Geese. Ardea 82:231–240
Gerdes K (2000) Die Vogelwelt des Landkreis Leer im Dollart und der Insel Borkum. Schuster, Leer
Greenwood PJ, Harvey PH (1982) The natal and breeding dispersal of birds. Annu Rev Ecol Syst 13:1-21
Jenness J (2001) Extension tool “Whitefronted geese” for ArcView 3.2. Jenness, Flagstaff, Ariz.
Kruckenberg H, Jaene J, Bergmann H-H (1996) Rastphänologie und Raumnutzung der Wildgänse am Dollart im Winter 1994/95. Vogelkd Ber Niedersachs 28:63–74
Lamprecht J (1986) Social dominance and reproductive success in a goose flock (Anser indicus). Behaviour 97:50–65
Madsen J (2001) Spring migration strategies in Pink-footed Geese Anser brachyrhynchus and consequences for spring fattening and fecundity. Ardea 89: 43–55
Nilsson L, Persson H (1991) An increasing breeding population of Greylag Geese Anser anser in Southern Sweden; a neck-banding study. Ardea 79:239–242
Nilsson L, Persson, H (1992) Feeding areas and local movement patterns of post-breeding Greylag Geese Anser anser in South Sweden. Orn Svecica 2:77–90
Prokosch P (1984) Population, Jahresrhythmus und traditionelle Nahrungsplatzbindung der Dunkelbäuchigen Ringelgans (B. b. bernicla) im Nordfriesischen Wattenmeer. Ökol Vögel 6:1-76
Raveling DG (1970) Dominance relationships and agonistic behaviour of Canada Geese in winter. Behaviour 306:291–319
Rees EC, Bacon PJ (1996) Migratory tradition in Bewick‘s Swans (Cygnus columbianus bewickii). Proc Anatidae 2000. Gibier Faune Sauvage Game Wildl 13:407–420
Ringleben H (1957) Saatgänse (Anser fabalis) als Durchzügler und Wintergäste in Deutschland. Vogelring 26:65–73
Robertson GJ, Cooke F (1999) Winter philopatry in migratory waterfowl. Auk 116:20–34
Stahl J, Drent RH, Ganter B, Ebbinge BS (2001a) Site fidelity and local dominance of brent geese during spring staging – a progress report. In: Stahl J (eds Limits to the co-occurence of avian herbivores. PhD Rijksuniversity Groningen, Haren
Stahl J, Tolsma PH, Loonen MJJE, Drent RH (2001b) Subordinates explore but dominant profit: resource competition in Arctic Barnacle Goose flocks. Anim Behav 61:257–264
Visbeen F, van der Waal W(1997) De Grauwe Gans (Anser anser) in Waterland-Oost. Graspieper 17:112–118
Warren SM, Fox AD, Walsh AJ, Merne OJ, Wilson HJ (1992) Wintering site interchange amongst Greenland White-fronted Geese Anser albifrons flavirostris captured at Wexford Slobs. Bird Study 39:186–194
Wilson HJ, Norriss DW, Walsh A, Fox AD, Stroud DA (1991) Winter site fidelity in Greenland White-fronted Geese Anser albifrons flavirostris, implications for conservation and management. Ardea 79:283–297
Acknowledgements
We like to thank Leif Nilsson, the coordinator of the Nordic Greylag Goose Project in Lund, Sweden, for information on his marked geese and for help with questions. For comments on an earlier draft we like to thank Barbara Ganter, Kees Koffijberg, Hans-Heiner Bergmann and Julia Stahl. Special thanks to Christoph Zöckler and Julia Stahl for help with the English manuscript. We thank the Lower Saxony Agency for Environment (NLÖ) for their financial contribution. Without this support this study would not have been possible.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by F. Bairlein
Rights and permissions
About this article
Cite this article
Kruckenberg, H., Borbach-Jaene, J. Do greylag geese (Anser anser) use traditional roosts? Site fidelity of colour-marked Nordic greylag geese during spring migration. J Ornithol 145, 117–122 (2004). https://doi.org/10.1007/s10336-004-0021-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-004-0021-1