
Abstract
While plant growth and productivity are known to derive from the interaction between genetic potential (G) and environmental factors (E), efforts to improve rice production have usually proceeded assuming a standard E that is created by conventional rice-growing practices. Genotypes have been assessed for their performance in continuously flooded paddy soils, with optimally dense plant populations, with reliance on inorganic fertilization to raise yields. The System of Rice Intensification (SRI) developed in Madagascar and now becoming accepted in much of Asia proposes that GxE interactions can be made more productive with different management practices: optimally sparse populations, established with very young seedlings carefully transplanted, intermittent flooding of paddies, with active soil aeration and with soil organic matter enhanced as much as possible. This article evaluates the effects of alternative SRI cultural practices on grain yield with particular attention to their impact on the growth and functioning of rice plant roots and on associated nutrient-use efficiencies that could be contributing to the observed higher grain yields. On-station experiments and on-farm surveys were conducted in Madagascar to evaluate SRI practices in comparison with standard cultural methods, considering how rice plants’ expression of their genetic potential was affected by different crop management practices. Controlling for both soil and farmer effects, rice plants cultivated with SRI methods produced average yields more than double those from standard practice (6.26 vs. 2.63 t ha−1). The most evident phenotypic difference was in plant root growth, assessed by root-pulling resistance (RPR), a summary measure of root system development. On average, uprooting single SRI plants required 55.2 kg of force plant−1, while pulling up clumps of three conventionally grown plants required 20.7 kg hill−1, or 6.9 kg plant−1. SRI plants thus offered 8 times more resistance per plant to uprooting. Direct measurements confirmed that SRI methods induced both greater and deeper root growth, which could be contributing to increased nutrient uptake throughout the crop cycle, compared with the shallower rooting and shorter duration of root functioning under continuous flooding. Rice plants grown with SRI methods took up more macronutrients than did the roots of conventionally managed plants, which was reflected in the higher SRI yields. When grain yield was regressed on nutrient uptake to assess nutrient-use efficiency, SRI plants achieved higher grain yield per unit of N taken up, compared to plants grown with conventional methods. The internal efficiency (IE) of SRI plants in utilizing macronutrients was 69.2 for N, 347.2 for P, and 69.7 for K, while the IE in plants conventionally grown was 74.9, 291.1, and 70.4 for these three macronutrients, respectively. Although no significant differences in IE were observed for N and K, the uptake of P was significantly greater, indicating more efficient use of P by SRI plants for grain production. More research needs to be done on such relationships, but this study indicates that productive changes in the structure and functioning of rice plants, particularly their roots, can be induced by alternative management methods.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Various innovations have been made in rice production systems in order to increase grain yield and meet the world’s continuing and growing demand for staple food. The most successful strategy for this has been the Green Revolution, which produced major yield increases in Asia where many farmers were able to adopt and benefit from this technology. However, this strategy has had markedly less impact in Africa, where farmers are constrained by their limited infrastructure and financial resources, plus less favorable soil and climatic conditions.
Changes made in rice genotypes (G) have produced relatively less yield improvement in Africa given the various conditions that constitute the crop environment (E) there. Renewed efforts are being made by governments and donor agencies to enable resource-limited farmers to improve their rice yields and thereby reduce food insecurity, so as to achieve the Millennium Development Goals of reducing hunger and poverty.
Lowland rice production has been practiced under continuously flooded soil conditions for millennia. Most research on rice that has sought to improve genetic potentials and/or management practices has accepted the long-established assumption that rice is best grown with standing water (Senewiratne and Mikkelsen 1961; Obermueller and Mikkelsen 1974; DeDatta 1981).
However, continuous saturation of soil can have suppressive effects on yield by causing alterations in rice root systems, most notably the deformation of their cortex and creation of aerenchyma (air pockets) (Kirk and Bouldin 1991), with consequent degeneration of roots (by as much as 78%—Kar et al. 1974). Hypoxic soil conditions caused by inundation can limit roots’ ability to respire and can slow down root metabolism, ion transport, and growth. Further, hypoxia leads to reduced soil conditions (low redox potential) that create low solubilities of some nutrient ions and high solubilities of others (Ponnamperuma 1984).
More than two decades ago, the System of Rice Intensification (SRI) was developed in Madagascar based on certain insights into how to improve the growing environment for rice plants by changing certain long-standing cultural practices (Laulanié 1993). Application of SRI principles has helped small farmers in that country to greatly increase their grain yields, from 2 t ha−1 to 8 t ha−1 and sometimes more (Hirsch 2000; Uphoff and Randriamiharisoa 2002) on soils that were evaluated as poor or very poor (Johnson 1994).
These increases were achieved not by introducing new varieties or increasing external inputs, but by changing the management of plants, soil, water, and nutrients as described below. SRI practices aim to provide rice plants with better growing environments both above- and below-ground so that previously unattained potential for root growth, tillering, and subsequent grain filling can be manifested.
The following practices are intended to produce a more favorable growing environment for rice plants, recognizing that some genotypes respond more positively than others to these changes in management. There can be some variation in seedling age, water application schedules, amount and kind of compost, etc. to suit local soil and other conditions and constraints. These practices are considered as a starting point for SRI rather than as a final, fixed recipe:
-
Transplanting very young seedlings, only 8–12 days old, rather than seedlings 3–4 weeks old or even more. Direct-seeded versions of SRI are also now being used, with no transplanting, which subject plant roots to no disturbance or trauma at all.
-
Planting single seedlings widely spaced, carefully and quickly, in a grid pattern, usually at least 25 × 25 cm, thereby reducing plant populations by as much as 80–90%.
-
Water management that avoids flooding throughout the entire crop cycle, with no continuously standing water during the vegetative growth phase and mostly aerobic soil conditions, thereby reducing water use by 25–50%.
-
Weeding with a simple mechanical implement that aerates the soil at the same time it eliminates weeds, rather than do this by hand or by use of herbicides.
-
Use of compost to enhance soil organic matter as much as possible, not relying on synthetic fertilizer, although this can be used if there is not sufficient biomass available.
Proponents of SRI consider these practices to work synergistically to give higher yield than conventional rice production systems, with more robust root systems supporting better above-ground growth, and vice versa (ATS 1992).
If substantially higher yields and factor productivity can be obtained from such changes in management practices, each fairly simple, with no requirement to purchase new-variety seeds or other inputs, this should be of interest to agronomists. In particular, it would be informative to know more about the nutrient dynamics within the soil–plant environment.
The System of Rice Intensification is currently practiced in over 40 countries located in Asia, Latin America, and Africa. Its dissemination in Madagascar was, however, constrained by certain factors, such as difficulties in exercising water control in many places, need initially for more labor during transplanting operations, and lack of government support. For many years, diffusion was left to NGO promotion and farmer-to-farmer spread, with no evident coordination nor systematic experience-sharing. Only since 2008 has the Ministry of Agriculture become more engaged with SRI, while collaboration among SRI promoters has been assisted by an SRI Secretariat supported by the Better U Foundation.
This study reported here compared the grain yield resulting from recommended SRI techniques with that from conventional cultural practices. It assessed the concomitant development of rice plant roots and analyzed the respective nutrient uptake and nutrient-use efficiency of plants grown with SRI compared to conventional cultural practices.
Two complementary data sets were generated, one from farmers’ fields in several locations and the other from a demonstration farm in central-eastern Madagascar. The on-station study evaluated the nutrient uptake of rice plants cultivated under SRI or conventional cultural systems, while the on-farm research investigated the impacts of cultivation practices on yield, root growth, and nutrient uptake under typical conditions.
By collecting data from matched pairs of fields, the methodology was designed to control for farmer and farm-field effects. The objective of both studies was to understand agronomic factors, particularly those below-ground, which could account at least in part for the observed differences in rice productivity that are associated with SRI’s alternative methods for management of plants, soil, water, and nutrients.
Although the System of Rice Intensification has been somewhat controversial, e.g., Dobermann (2004), Sheehy et al. (2004), and McDonald et al. (2006), a number of evaluations have shown substantial improvements in the productivity of land, labor, and water with these methods, e.g., Ceesay et al. (2006) in the Gambia; Sinha and Talati (2007) in India; Sato and Uphoff (2007) in Indonesia; Kabir and Uphoff (2007) in Myanmar; and Namara et al. (2008) in Sri Lanka. To date, few detailed agronomic analyses have examined the mechanisms involved, although several have initiated this study, such as Mishra and Salokhe (2008), Zhao et al. (2009), and Thakur et al. (2010a,b). Other contributions to this issue are also contributing to a better scientific understanding of empirical observations.
Materials and methods
An on-farm survey was undertaken during 2000–2001 in four different locations of Madagascar, involving 109 farmers who were using both SRI and conventional methods concurrently on their farms. Data were gathered on agronomic factors affecting the SRI and conventional systems under typical on-farm field conditions. At the same time, to better understand plants’ nutrient uptake and nutrient-use efficiency, an on-station study was conducted under controlled conditions as discussed below. This permitted more precise evaluation of grain yield production and its relationship with root growth and functioning.
The System of Rice Intensification results were compared with those of the Système de Riziculture Améliorée (SRA), which is the system of rice cultivation promoted by the Madagascar government. SRA, which is practiced now by ~20% of Malagasy farmers, relies on transplanting older (~25 days) seedlings, 3 per hill with row spacing of 20 cm, continuous flooding of ~5 cm, mechanical weeding and chemical fertilizer (~300 kg ha−1 of NPK 11-22-16) with some addition of cattle manure. At the same time, comparison was made also with prevailing farmer practice (FP). This, like SRA, maintains higher plant population and paddy flooding, but even older seedlings are transplanted, in a random manner and with less care, and with less provision of nutrients for the soil. These three different systems compared represent a continuum of intensified management.
On-station experiments
Experimental site, treatments, and design
These experiments were conducted during the 2000–2001 main growing season, November through April, at the Center for Diffusion of Intensified Agriculture (CDIA) in Beforona, located roughly midway between the capital Antananarivo in the central highlands and Tamatave on the eastern seacoast. The research was done in collaboration with the CDIA team implementing the Landscape Development Interventions (LDI) project, funded by USAID to help conserve biodiversity in Madagascar’s forest zone and improve farmers’ living conditions. The station is at an elevation of about 500 m, located at 48°30′E and 18°50′S. Its soils are mostly orthic and xantic ferrosols. Mean temperature during the cropping season ranges between 27 and 32°C.
The trials were done in a clay-sandy soil with 43.8 g organic matter kg−1, 27 g organic C kg−1, 1.88 g total N kg−1, 17.8 g available P kg−1 (Olsen method extraction), 2.6 cmol(+) kg−1 cation exchange capacity, and 0.15 cmol(+) kg−1 exchangeable K. The trial plots had been previously used for traditional rice cultivation until 1999, with no nutrient amendments, either manure or plant residues. During 1999 and 2000, beans and vegetables such as peppers and cabbage had been planted, with compost applications made from household waste. Prior soil fertility management was the same for all the plots used in the experiment.
Five treatments were evaluated in a completely randomized block design, each with three replications:
- T1::
-
SRI cultivation methods with compost application
- T2::
-
SRI methods without compost
- T3::
-
SRA methods with chemical fertilizer (NPK 11-22-16)
- T4::
-
SRA methods without fertilizer
- T5::
-
Conventional farmer methods of cultivation
The trials were designed to evaluate the effects of compost application with SRI and of chemical fertilizer application with SRA.
Compost-fertilized SRI plots received an incorporation of 10 t ha−1 of compost composed of bush vegetation, pig manure, and soil 8 days before transplantation (analyzed as having 8 g N kg−1, 45.5 mg P kg−1, and 9.25 K g kg−1). The fertilized SRA plots received 300 kg ha−1 of NPK 11-22-16 just before transplanting, and 67 kg ha−1 of urea at the panicle initiation stage. The differences among the three cultural systems being evaluated are specified in Table 1.
The size of plots was 20 m2 (4 × 5 m), and grain yields were measured from a 9 m2 subplot sample located in the center of each plot. Grains were weighed right after the harvest, with their moisture content measured and calculated grain yields adjusted to a standard 14% moisture content. Components of yield—tillers clump−1, panicles clump−1, and grains panicle−1—were also recorded, determined from 12 hills plot−1 distributed in 3 subseries of 4 hills.
Measurements of root-pulling resistance (RPR) and root length density (RLD)
Above-ground measurements were complemented at harvest time by an evaluation of both root-pulling resistance (RPR) and root length density (RLD).
Root-pulling resistance evaluates root growth and rooting density in a summary way, by measuring the amount of force that is required to uproot a single plant or set of plants in a hill (Ekanayake et al. 1986). While measurements of RPR, which evaluates the combined, cumulative effect of the respective crop management practices, can be affected by differences in soil conditions (structure, moisture), RPR is a meaningful indicator if such conditions are kept reasonably similar and if the differences observed are large, as they were in this study.
To minimize the effects of variability in soil characteristics, the soil for all the samples evaluated was kept at the same moisture level for a week before the plants were pulled up to assess RPR. For determining RPR, a sample of 3 plants per plot was taken. A scale was attached by a rope to the base of each plant to be uprooted, and the scale was pulled up slowly and evenly until the rice plant’s root was released from the soil. The force (in kg) required to pull up the plant was recorded at the moment of uprooting.
Root length density was determined from root samples taken when the crop was harvested. The most representative plants were chosen in each plot, and circles with diameters of 27.5 cm for SRI, 21 cm for SRA, and 17.5 cm for conventional system were delimited around the roots of each plant. (These diameters were determined after first making observations of the diameters of the respective root systems.) A trench was then dug, and soil layers were cut horizontally (and removed) at 5, 10, 20, 30, 40, and 50 cm depth.
These blocks of soil were then washed in a bucket of water in order to separate the roots from the soil. Roots were separated through repeated filtration using 1 mm and 0.5 mm mesh before weighing. A 1 g subsample was spread on graph paper, and the numbers of intersections between the root and the paper grids were counted, following the method of Goubran and Richards (1979).
Analysis of macronutrient contents
To evaluate the nutrient content of the respective rice plants, plant samples were analyzed for their macronutrient content (N, P, and K) at the maturity stage, separated into harvestable biomass (grains) and non-harvestable biomass (straw). After being oven-dried at 70°C, weighed and ground, N content was measured by micro-Kjeldahl digestion (Bremmer and Mulvaney 1982); P content by the molybdenum blue colorimetric method (Yoshida et al. 1972); and K content by spectrophotometer atomic adsorption (Yoshida et al. 1972).
In addition, soil samples were collected at the beginning of the growing season from five locations in each plot at a depth of 0–20 cm for the SRA and conventional systems, and at a depth of 0–30 cm for the SRI system, because the latter plant roots grew more deeply.
Statistical analysis
All data were statistically analyzed using analysis of variance (ANOVA) as applicable to a completely randomized block design. The significance of the treatment effect was determined using an F-test, and to determine the significance of the difference between the means of the two treatments, least significant differences (LSD) were calculated at the 5% probability level. MINITAB software was used for the data analysis.
On-farm evaluations
Work done by Witt et al. (1999) has shown grain yield to increase in an essentially linear way, correlated with increases in nutrient uptake until a certain level is reached where one or more of the critical nutrients becomes limiting. (This analysis assumes that other factors such as climate, soil moisture, and disease as well as micro-nutrients are non-constraining.) Once the plant’s efficient use of a particular nutrient is limited by others, marginal increases in grain yield relative to nutrient uptake start to decline.
Since plants cultivated with SRI methods appeared able to produce higher grain yield under the same soil conditions as those cultivated under the conventional system (Andriankaja 2001), it was hypothesized that under SRI management, the nutrients taken up by the plants are used more efficiently for grain production. Whether this is due to physiological changes, differential expression of genetic potentials, or phytohormonal promotion could not be investigated. In this study, we assessed the prior question: whether, or to what extent, the hypothesized changes in nutrient uptake and plant utilization were significantly associated with respective cultural practices.
Prior to our survey, a full list of farmers practicing SRI in the four study areas described below was obtained from the Ministry of Agriculture and from Association Tefy Saina, the Malagasy NGO that has been most active in promoting SRI in the country. Farmers were interviewed initially in order to get a factual characterization of their farming systems and to determine whether they were practicing both SRI and conventional systems in comparable ways. Only farmers who were using both systems similarly were maintained in our sampling population in order to hold both farm and farmer variables constant.
Interviews acquired data on the characteristics of each farmer’s SRI and conventional management practices, ascertaining the age of seedlings at transplantation, the number of seedlings per clump, mode of weeding, type of water management, and type of fertilization. Fields were classified as being cultivated conventionally or with SRI methods according to two criteria: (1) the age of seedlings transplanted, and (2) the number of seedlings per hill.
Fields that had seedlings >20 days old at transplanting and 3 or more seedlings transplanted hill−1 were considered as conventional cultivation. SRI fields, on the other hand, were those transplanted with seedlings 8–12 days old and had only 1 seedling hill−1. Other factors such as spacing, water management, and/or fertilization practices were not taken as criteria for classification in order to be able to assess the effects of any variability in these other factors. Farmers were selected based on whether they had at least one rice field that met each of these two sets of criteria.
The total number of households selected in the four study areas was 109 farmers. These areas were located in three of the most important rice-producing areas in Madagascar:
-
Ambatondrazaka region around Lake Alaotra, the largest rice-producing area in the country: one sample area (Zone I) was southeast of the lake area with a sample size of 40 farmers, each cultivating with both SRI and conventional methods, and the other (Zone II) was northeast of the lake with a sample size of 30;
-
Antsirabe region in the central highlands: this sample area (Zone III) was located to the north and northwest of this city with a sample size of 28; and
-
Fianarantsoa area in the southern highlands: this sample area (Zone IV) was located northwest of this provincial capital with a sample size of 11.
The System of Rice Intensification and conventional plots were selected without any reference to their productivity. The initial selection of plots was done at random within each farm included in the study, as much as possible identifying matching fields close to each other so as to reduce the effects of physical variability, soils, topography, and microclimate.
Yield was evaluated by measuring grain production on 5 sampled squares 1 × 1 m2 along two diagonals in the rice plot. Border effects were eliminated by leaving a 1 m-border along the plot. Rice plants within the 1 × 1 m2 were harvested, threshed, and winnowed. Paddy was then weighed, and its humidity was measured. Grain yield was estimated by averaging the 5 sampled squares. Macronutrient content of plants and soil were evaluated in the on-farm study using the same methods as described above for the on-station trials.
Harvest index was measured from random subsamples of 12 hills plot−1 distributed in 3 subseries of 4 hills. All plants were separated into non-harvestable (tillers, leaf blades and stems) and harvestable biomass (grains). Plant parts were then oven-dried at 70°C, and dry weights were recorded. Harvest index was calculated as a ratio of grain weight to total above-ground crop dry weight.
Results
On-station experiments
Grain yield and yield components
Substantial differences were observed in the grain production from the respective SRI, SRA, and conventional systems in on-station trials (Table 2). The highest average yield, 6.26 t ha−1, was obtained from those plots where SRI was used with compost applied. This was statistically significantly different from the yields obtained with SRA methods: 4.92 t ha−1 with NPK and urea applications, and 4.67 t ha−1 from non-fertilized SRA plot. In conventional practice, grain yield was only 2.63 t ha−1.
While farmer methods gave a higher number of panicles m−2, the number of grains per panicle and per m−2 was lower. With smaller panicle size, grain yield was reduced significantly.
Planting on the trial sites was done, unfortunately, 1 month later than recommended for the Beforona area because of logistical problems in getting the trials started. This delay would have had some effect on grain yield production for all five treatments, but probably more in the SRI trials because SRI performance depends in part on profuse tillering, and late planting would have constrained SRI plants’ tillering somewhat. The non-significant yield difference seen between fertilized and non-fertilized SRA plots could be attributed to the greater adverse effect that blast (Pyricularia oryzae) had in the fertilized plots during their grain-filling phase.
Root-pulling resistance and root length density
One of the key differences resulting from use of SRI methods in this study was better growth of root systems, with more proliferation as well as greater longevity of roots. As described below, differences in root growth between SRI and conventionally grown plants were assessed by two complementary methods of measurement.
Single SRI plants had demonstrably greater root-pulling resistance (RPR) compared to clumps of multiple rice plants grown with the two other methods evaluated in the on-station trials (Table 3). RPR for individual SRI plants ranged from 49.7 to 55.2 kg, whereas for SRA clumps (2–3 plants), average RPR was 30.0–34.1 kg. Rice plants grown with farmer practice in clumps of 4–6 had an RPR of 20.7 kg. These differences in RPR between SRI and the other cultivation methods were clear and statistically significant.
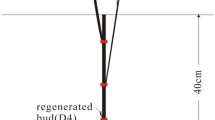
On a per-plant basis, the force needed to uproot SRI plants was 4–10 times more than for plants grown with the modern or traditional cultivation methods that farmers are now using. The difference can be observed in Fig. 1 which compares the root systems of conventionally grown and SRI plants.

Root growth of three conventionally grown rice plants transplanted at 28 days (on left) versus roots of a single SRI plant transplanted at 8 days (on right)
Given that the three cultivation systems had substantial differences in inter-plant spacing, root pulling by itself is not a sufficient or always accurate measure of better rooting (Morita and Suga 1988). Accordingly, we measured root length density (RLD) at different soil depths to assess the growth and proliferation of roots. With all of the different treatments, root growth was seen to vary greatly by soil depth, as seen in Fig. 2, which shows the different profiles of rice root depth associated with the alternative management systems.

Root distribution at different depths for rice plants grown under SRI, SRA, and conventional systems
Conventional and SRA root systems were found to have greater growth in the first 20 cm in comparison to SRI plants. Indeed, close to the soil surface (0–10 cm), the greatest root growth was seen in rice cultivated by conventional farmer methods. At a depth of 20–30 cm, however, the growth of plant roots was similar for all three methods.
Significant differences were seen below 30 cm. Much greater root growth was measured there for SRI plants, double and even three times what was seen for plants grown conventionally or with SRA methods. This indicates that rice plants’ root growth benefited from the cyclical application of water, with an alternation of aerobic and anaerobic soil conditions, as well as from the provision of more organic matter to the soil.
On-farm evaluation
Grain yield and harvest index comparisons
Harvest index (HI), the ratio of harvested yield to total plant biomass, has been a major concern of plant breeders who sought to achieve more productive rice varieties by reducing the proportion of non-consumable biomass. This would increase plants’ production of grain in relative terms. In the farmer-survey plots, we found that SRI grain yields were significantly higher, averaging 6.36 t ha−1, compared to 3.36 t ha−1 with conventional methods (Table 4).
This 89% increase over conventional grain yield was 218% higher than average rice yield in the country as a whole, 2 t ha−1.That the farmers in our sample had higher yields with their own methods than the national average can be explained at least in part by the larger number of farmers from the Ambatondrazaka region in the sample. Here, many ‘modern’ methods are already part of standard cultivation practice, and the soils are generally more productive than elsewhere in the country. But it is also possible that the farmers in our sample who were using both conventional and SRI practices were more capable farmers than average (Barrett et al. 2004). The norm with which SRI performance is being compared in this on-farm study is higher than typical in Madagascar.
The increase in grain yield from SRI plants was accomplished with a harvest index quite similar to that for conventionally grown rice plants, even though the SRI plants had a significantly higher number of tillers. The harvest index for plants grown with SRI methods was 0.48, while that for plants raised with conventional cultivation methods averaged 0.49 as seen in Table 4.
The HI distributions analyzed by quartile were also similar. Index values for the first and third quartiles of conventionally grown rice plants ranged from 0.32 to 0.63, while for SRI plants, the index numbers varied from 0.33 to 0.67, indicating practically no difference. Furthermore, comparisons of nutrient harvest indexes indicated very similar relationships. These indexes calculated for SRI plants were 0.68 g N g−1, 0.71 g P g−1, and 0.27 g K g−1, while the ratios calculated for conventional rice plants were 0.65 g N g−1, 0.72 g P g−1, and 0.25 g K g−1. These similarities made assessment of nutrient uptake, accumulation and partitioning all the more important.
Nutrient concentration and uptake by rice plants
Increased SRI grain yield could have been associated with dilution of nutrient concentrations in these plants’ shoots and grains. When the nutrient content in the foliage was evaluated, we found that plants cultivated with SRI methods accumulated 4.98 g N kg−1, 0.93 g P kg−1, and 14.98 g P kg−1 in their straw (Table 4). With conventional management practices, the average straw nutrient content was indeed slightly higher, with accumulation of 5.39 g N kg−1, 1.16 g P kg−1, and 15.29 g K kg−1, with the difference in amounts of N and P being statistically significant.
However, nutrient translocation, i.e., the ratio between nutrients in the grain and total above-ground nutrients, was almost the same for both SRI and conventional cultivation. The respective percentages were 68% N, 71% P, and 27% K for SRI plants, and 65% N, 72% P, and 25% K with conventional cultivation methods. While SRI plants significantly out-yielded conventional plants, their respective nutrient translocations were quite similar. This suggests that differences in root structure and functioning post-anthesis (Fig. 2 and Table 3) could be contributing to the divergent outcomes.
This would be consistent with the greater nutrient accumulation measured in SRI plants’ above-ground biomass (Table 5). With SRI management, total above-ground nutrient accumulation averaged 95.07 kg N ha−1, 21.03 kg P ha−1, and 108.64 kg K ha−1, while with conventional practice, these amounts were 49.99 kg N ha−1, 12.69 kg P ha−1, and 56.77 kg K ha−1. These data confirm that nutrient uptake can be considerably enhanced by management practices that affect tillering and root development, given that N and K accumulation were enhanced by 91%, and the uptake of P was increased by 66% with SRI practices.
Internal nutrient efficiency
Average internal nutrient efficiencies (IEs) are shown in Table 6. IE values with SRI management methods were 69.2 kg grain per kg plant N, 347.3 kg grain per kg plant P, and 69.7 kg grain per kg plant K. This was equivalent to 14.5 kg N, 2.9 kg P, and 14.3 kg K per 1000 kg grain. With conventional practices, on the other hand, average IEs were 74.9 kg grain per kg plant N, 291.1 kg grain per kg plant P, and 70.4 kg grain per kg plant K, equivalent to 13.4 kg N, 3.4 kg P, and 14.2 kg K per 1000 kg grain. Although nitrogen IE was a little higher with conventional management compared to the level with SRI, the difference was not significant at the 5% level (t-test P = 0.197).
A significant difference was observed with regard to P-use efficiency, on the other hand. More efficient use of P for grain production when using SRI cultivation methods is seen in Table 6. This is apparently the result of higher N uptake by SRI plants, evident from the measured N:P:K ratios, i.e., the ratio of N to P and of K to P. With SRI management, the nutrient ratio is considerably higher than with conventional practice—5.0:1.0:4.9 for SRI compared with 3.9:1.0:4.1 for the latter.
Regression analysis of rice grain yield on N uptake for plants grown, respectively, with SRI or with conventional methods, was done assuming a parabolic relationship. This analysis indicated that with regard to N uptake, conventionally grown plants have a much faster decrease of internal efficiency (Fig. 3). This decrease in grain production in response to marginal increases in N is expressed by the second degree of the parabolic equation having a coefficient of −0.229 for conventional practice and −0.064 for SRI methods. This means that increments to grain yield declined more rapidly in conventionally grown plants as these increased their uptake of N, compared with SRI plants. Calculations for P and K indicated the same differences in their relationship for conversion of the uptake of these macronutrients into grain (not shown).

Relationship between N uptake and grain yield for rice plants grown with SRI versus conventional methods (Poly polynomial regression line)
The coefficients of the first degree parabola—58.849 for SRI, and 45.631 for conventional practice—conversely reflected a steeper increase of the SRI grain yield as a function of N uptake. The difference in the respective abilities of these two categories of rice plant to convert the N taken up into grain yield reflects phenotypical divergence in terms of plant structure and physiology resulting from the same or similar genotypes (G). This warrants further consideration to assess what could account for such divergence in plants’ capacities in response to management changes in their growing environment (E).
Discussion
The relative increase of grain yield with SRI methods could have resulted from the farmers in our sample allocating their best sites, i.e., their best soils, to SRI production, or possibly to their applying more compost to their SRI plots. However, this could not explain the differences observed.
Soil analyses using standard measurements showed quite similar soil fertility in both the SRI and conventional plots using standard assessment methods (Table 7). In the plots where SRI methods were used, average soil nutrient content was 0.16% N, 8.51 ppm P-Olsen, and 0.08 cmol (+) kg−1 K, while where rice was grown conventionally, these values were 0.17% N, 9.39 ppm P, and 0.09 cmol(+) kg−1 K.
Further, from our investigation of farmers’ actual practices, we know that only six of the farmers in our sample used any compost on their fields, and their grain yields were practically the same as those of other farmers. That so few farmers used compost with their SRI practices suggests that the success of SRI does not depend primarily on compost use. Association Tefy Saina, the main proponent and promoter of SRI in Madagascar, has always considered the use of compost to be an accelerator, giving better results when used with the other practices, but not as necessary for the other SRI methods to give higher yields.
When we excluded from the comparison the results of those farmers who applied compost, average grain yields hardly changed. Without compost, SRI practices produced 6.35 t ha−1, still 89% more than conventional methods. The latter produced 3.36 t ha−1 on essentially the same soils, being managed by the same set of farmers.
Overall, the higher grain yield with SRI appears to be related to both greater and more balanced nutrient uptake. For plants to benefit from the uptake of macronutrients, they need to have sufficient, although often miniscule, supplies of micronutrients to supplement their macronutrient acquisition. More balanced nutrition of SRI plants could be due to differences in the indigenous supply of nutrients in the topsoil. However, this study’s on-station and on-farm results suggest that the differential uptake and output parameters for SRI versus conventionally grown rice plants may be related more to SRI plants’ root systems exhibiting greater activity, longer functioning, and deeper and more extensive proliferation (Fig. 1).
According to the ‘limiting factor’ explanation of plant nutrition and growth, the observed differences in SRI and conventional plants’ productivity could be influenced by their respective access to micronutrients, which would be affected by the size and functioning of plant root systems. However, because this study evaluated only macronutrient relationships, nothing conclusive can be said on this question of the role and contribution of micronutrients. This remains a subject for further investigation.
Root-pulling resistance
The differences measured in root-pulling resistance (RPR)—as much as 4–10 times more force per plant was needed to uproot SRI plants compared to the others (Table 3)—could result from the soil aeration maintained with the SRI water management regime. Keeping paddy soils wet but not continuously saturated during the vegetative phase would have maintained mostly aerobic soil conditions, enhanced also by the early and frequent mechanical weeding that disturbs the top layers of the soil between plants. These practices appear to have contributed to SRI plants having better access to nutrients and to plants meeting their nutrient requirements more adequately throughout the cropping cycle. With more space for both roots and canopy to grow, SRI plants developed larger and deeper rooting systems compared to SRA and conventionally grown plants. These differences were seen from measurements of RPR at critical stages of growth, from RLD at lower soil depths, and from visible comparison (Fig. 1).Footnote 1
Harvest index
In this study, harvest index considerations offered no explanation for the higher yield with SRI methods. In an assessment of the physiological effects of SRI practices (Thakur et al. 2010a), harvest index was found to play little role in the enhancement of SRI rice yields, although in another study by Thakur et al., reported in this volume, there was a significant difference in harvest index. Here in the Madagascar comparisons, we found that although the SRI plants had a higher number of tillers, which normally results in more non-harvestable biomass and thus in a lower HI, their harvest index was similar to, and in some cases even higher than the index for conventionally grown rice.
The more profuse tillering promoted by SRI management practices, including the use of young seedlings and wider spacing, is closely linked with a more extensive root system. In rice plants as in other grass-family (gramineae) species, both roots and tillers emerge concomitantly from the same meristematic tissue at the surface-level base of the plant. Nodal roots appear in association with the emergence of each newly formed tiller.
The System of Rice Intensification root systems appear to benefit—becoming larger and functioning longer—from the soil aeration that results from non-flooded water management and from mechanical weeding, as well as from the enhancement of soil organic matter. These practices at the same time support and promote more tillering. SRI practices in combination appear to help rice plants realize more fully their potential for tillering and root growth. Evidence of the differences that result from these alternative management practices is seen in Tables 2, 3, 4, 5, and 6 (also Thakur et al. 2010a).
Nutrient uptake
Larger root systems enable plants to access a greater volume of soil and to acquire more nutrients from various depths. Further, under aerobic soil conditions, root systems experience less deterioration and senescence, continuing to function beyond panicle initiation (Kar et al. 1974). Under continuous submergence with sustained hypoxia, as much as 75% of rice plant roots degenerate by the flowering stage, when grain formation begins. Aerobic soil conditions are generally more favorable for root functioning compared to anaerobic circumstances (Drew 1997; Kirk and Bouldin 1991; Kirk and Solivas 1997).
In plants cultivated with SRI methods, the nutrient accumulation in grains averaged 10.18 g N kg−1, 2.35 g P kg−1, and 3.96 g K kg−1, while with conventional methods, this accumulation was 9.90 g N kg−1, 2.69 g P kg−1, and 3.54 g K kg−1 (Table 4). Nutrient concentrations in sink storage were thus not very different between the two management systems. Grain yield was significantly higher, however, with SRI cultural methods. This could be attributable to conventionally cultivated plants having lower root capacity to take up nutrients in the later stages of plant growth, and/or to lower remobilization of previously stored shoot nutrients. But a full explanation for the significant differences in yield remains to be arrived at.
A relatively higher increase of accumulated N and K in SRI plants compared to their increase of accumulated P could be due either to (a) somewhat lower N and K uptake by conventionally grown plants or (b) relatively higher uptake of P with SRI management. Comparing grain yield with nutrient content gives some indications about the nutrient uptake constraints on yield, considering the concentration differences in grain produced by SRI versus conventional methods. With our data we can address the question of whether there is some dilution effect with the higher yield. But first we should consider these relationships in comparative perspective.
Previous estimates by Witt et al. (1999) of rice plants’ above-ground nutrient accumulation in subtropical and tropical Asia have showed a nutrient uptake of 91 kg N ha−1, 16 kg P ha−1 and 88 kg K ha−1, with average grain yield of 5.2 t ha−1. This N uptake was actually quite similar to what we found in our Madagascar study, although the estimated P and K uptake in Asian rice was somewhat higher. That the SRI grain yield which we measured was higher than that reported by Witt et al. could be due to differences in growing conditions. However, differences in the methods of cultivation appear to have contributed to the SRI results. The figures reported by Witt et al. are derived from what we are considering as conventionally grown rice plants, with limited functioning of root systems.
The data on nutrient uptake in rice plants that we analyzed were not much different from the figures reported by Witt et al. (1999), who found averages of 7.1 g N kg−1, 1.0 g P kg−1, and 14.5 g K kg−1. While these divergences from our results could be due to the effects of agroecological conditions, varieties used, or cultural methods, the SRI grain yield in the on-station trials was increased by 89% over conventional methods, similar to those that would have been used for growing the rice that was evaluated by Witt et al. Given that there were no substantial differences in the nutrients accumulated by the SRI rice plants compared to plants conventionally grown, there does not appear to have been any significant dilution of plant nutrients by the higher yield attained with SRI, an interesting observation.
There might be some increase of available soil N at various times during the growing season due to higher mineralization of organic-N when there were alternations between aerobic and anaerobic soil environments (Birch 1958). The juxtaposition of aerobic and anaerobic soil conditions has been shown to have a positive effect on biological nitrogen fixation (Magdoff and Bouldin 1970). These effects are considered below. Further, we note that it is reported that continuously anaerobic soil conditions can have the effect of immobilizing the N in soil organic matter (Schmidt-Rohr et al. 2004; Olk et al. 2006). All these are findings that warrant further study.
In the SRI plant-soil environment, we have seen that greater activity of endophytic N-fixing bacteria can occur within roots as well as within the root rhizosphere, substantially enhancing yield (Randriamiharisoa 2002; Randriamiharisoa et al. 2006). Biological N-fixation was not evaluated in this study; however, greater N uptake was evident in the plants’ growth and in their grain yield. The N for these increases had to come from somewhere. There were similar P levels measured in the soil of plots where both SRI and conventional rice were grown. Yet 66% more P was accumulated in the above-ground biomass of the SRI plants.
Enhanced root growth with SRI practices could have enabled these plants to avail themselves of otherwise-unavailable subsoil P that could not be accessed by plants grown with conventional methods. But these relationships could have involved microorganisms and not just the plants themselves. One possible factor might be that rice plants with aerobic root zones can benefit from the services of mycorrhizal fungi, which enhance plants’ access to P and other nutrients in the soil. Under continuously anaerobic soil conditions, these beneficial soil organisms are likely to be mostly absent (Ilag et al. 1987).
It is also possible that SRI practices, with alternate flooding and drying of soil, could increase the microbial solubilization of P. Most reported measures of P in soil samples are of available P. The amount of unavailable P, complexed in forms not accessible to plant roots, can be 20–30 times greater than what is available. Microbial activity supported by modified soil and water management practices could be making unavailable P available to the plants (Turner and Haygarth 2001; Turner et al. 2003). The article in this issue by Anas et al. showed increased populations of phosphate-solubilizing bacteria in the rhizospheres of rice plants grown with SRI methods, from research in both India and Indonesia.
In any case, the attainment of higher grain yield with SRI cultivation methods requires higher nutrient uptake, and this needs to be explained. Results from our on-farm survey indicated a doubling of N uptake by plants grown with SRI methods in comparison to conventional methods, even though the SRI and conventional plots had similar soil fertility in terms of chemical availability. The higher uptake of N by SRI plants suggests possibly greater activity of nitrogen-fixing bacteria living as endophytes within roots (Dazzo and Yanni 2006), or as free-living microbes within the rhizosphere (Boddy et al. 1995), or possibly even within the phyllosphere (Feng et al. 2006).
The System of Rice Intensification management practices, which include alternating the irrigation and drainage of soil, could be releasing more available N through mineralization processes (Birch 1958). Such processes could lead to a mining of the organic-N pool of the soil. This pool can be expanded through biological processes such as microbial N2-fixation and/or nitrogen cycling by soil organisms, e.g., protozoa and nematodes (Bonkowski 2004).
It is known that alternating irrigation and drainage leads to a fluctuation of NH4 + and NO3 − in the soil solution. This could render the SRI soil environment more prone to N loss. However, it is also known having a mix of NH4 +–N and NO3–N in the soil enhances rice production, by as much as 40–60% compared to having N available only in ammonium form, which is predominant in continuously flooded soil (Kronzucker et al. 1999). Also, losses of N from leaching could be offset by higher levels of soil N resulting from a mixing and juxtaposition of aerobic and anaerobic soil conditions (Magdoff and Bouldin 1970).
Much more remains to be known about the N dynamics in soils managed according to SRI recommendations and about other dynamic relationships governing nutrient availability and plant-soil-nutrient interactions. As more is learned about the impacts of SRI practices on plant performance (e.g., Thakur et al. 2010a; Zhao et al. 2009), more interest is likely to be elicited in understanding soil-plant-nutrient processes, with particular attention to the impacts and contributions of the soil biota (Randriamiharisoa et al. 2006).
Conclusions
Results from both on-station experiments and on-farm surveys showed consistently and significantly better performance of SRI rice plants relative to those grown with conventional rice practices. SRI cultivation methods appear to be contributing to better nutrient access and/or greater uptake by the rice plants, explainable by greater root growth, quantified by measured differences in root-pulling resistance and in root length density. Greater nutrient uptake is attributable to greater root growth and penetration to lower soil horizons, with higher root length density particularly below 30 cm depth. This enables SRI plants to exploit a greater volume of soil than can plants grown with conventional methods.
It is also likely that the flooding and draining of paddy soil results in faster mineralization of soil organic matter, which contributes to a greater supply of nutrients relative to conventional rice management. Soil microbial populations are likely to have been changed and even enhanced by SRI plant, soil, water, and nutrient management practices as these practices create more aerobic soil conditions and provide more substrate for the soil biota. However, such variables in plant-soil-microbial interactions were not studied here.
Two findings from this study are particularly noteworthy since they have not been documented previously. First, despite the increased tillering and higher grain yield of SRI rice plants, we did not find any difference in harvest index between SRI and conventional rice. All organs of the rice plant increased together under SRI management rather than some plant organs increasing faster than others. Second, nutrient-use efficiency under SRI cultivation methods was demonstrably higher, especially with respect to P. Research from China has reported similar higher nutrient-use efficiency for N with SRI practices (Zhao et al. 2009). These observations, in conjunction with the documented differences in root length density and root-pulling resistance, suggest that the improved performance of rice under SRI management practices is related, at least in part, to a proliferation of root systems under SRI cultivation methods and thus to better plant access to soil nutrients.
The measurements reported in this article add to the documentation of the effects of SRI practices in comparison with conventional ones for growing irrigated rice. From a single study, one cannot draw definitive conclusions about the mechanisms for the more productive phenotypes observed with SRI management, reported now from more than 40 countries (http://ciifad.cornell.edu/sri/).
A number of issues are identified here concerning SRI practices that should be further investigated, to develop a better understanding of the physiological, nutritional, edaphic, microbiological, and other scientific aspects underlying this cultural system. Detailed agronomic studies on SRI are beginning to appear in the literature, e.g., Mishra and Salokhe (2008), Thakur et al. (2010a,b), and Zhao et al. (2009). It appears likely that further studies on the under- and above-ground dynamics and the plant capacities that can result from alternative management practices will prove fruitful, both for farmers and for scientific knowledge.
Notes
We could not assess in this study the possible effects on root growth of the production by aerobic soil bacteria and fungi of phytohormones, which are known to stimulate root growth (Frankenberger and Arshad 1995). These effects remain to be investigated to see whether they contribute to differences in root growth and plant performance arising from alternative plant, soil, water and nutrient management practices. Soil organisms provide many benefits to plants beyond N2 fixation and P solubilization (Doebbelaere et al. 2003).
References
Andriankaja AH (2001) Mise en evidence des opportunités de développement de la riziculture par adoption du SRI, et évaluation de la fixation biologique de l’azote. Mémoire de fin d’etudes, Ecole Supérieure des Sciences Agronomiques, University of Antananarivo, 91 pp
ATS (1992) Presentation technique du système de riziculture intensive basée sur le modèle de tallage de Katayama. Fanabeazana Fampandrosoana Ambanivohitra. Association Tefy Saina, Antananarivo, 31 pp
Barison J (2003) Nutrient-use efficiency and nutrient uptake in conventional and intensive (SRI) cultivation systems in Madagascar. M.Sc. thesis for Department of Crop and Soil Sciences, Cornell University, Ithaca, NY
Barrett CB, Moser CM, Barison J, McHugh OV (2004) Better technologies, better plots or better farmers? Identifying changes in productivity and risk among Malagasy rice farmers. Am J Agric Econ 86:869–888
Birch HF (1958) The effect of soil drying on humus decomposition and nitrogen. Plant Soil 10:9–31
Boddy RM, Ocde O, Urquiaga S, Reis VM, Olivares F, Baldini VLD, Döbereiner J (1995) Biological nitrogen fixation associated with sugar cane and rice: contributions and prospects for improvement. Plant Soil 174:195–209
Bonkowski M (2004) Protozoa and plant growth: the microbial loop in soil revisited. New Phytol 162:616–631
Bremmer JM, Mulvaney CS (1982) Nitrogen-total. In: Page AL, Miller RH, Keeney DR (eds) Methods of soil analysis, part 2: chemical and microbiological properties. Agronomy Society of America, Madison, WI, pp 595–623
Ceesay M, Reid WS, Fernandes ECM, Uphoff NT (2006) The effect of repeated soil wetting and drying on low land rice yield with System of Rice Intensification (SRI) methods. Intl J Agric Sustain 4:5–14
Dazzo FB, Yanni YG (2006) The natural Rhizobium-cereal crop association as an example of plant-bacterial interaction. In: Uphoff N et al (eds) Biological approaches to sustainable soil systems. CRC Press, Boca Raton, FL, pp 109–127
DeDatta SK (1981) Principles and practices of rice production. McGraw-Hill, New York
Dobermann A (2004) A critical assessment of the System of Rice Intensification (SRI). Agric Syst 79:261–281
Doebbelaere S, Venderleyden J, Okon Y (2003) Plant growth-promoting effects of diazotrophs in the rhizosphere. Crit Rev Plant Sci 22:107–149
Drew MC (1997) Oxygen deficiency and root metabolism under hypoxia and anoxia. Ann Rev Plant Physiol Plant Mol Biol 48:223–250
Ekanayake IJ, Garrity DP, O’Toole JC (1986) Influence of deep root density on root pulling resistance in rice. Crop Sci 26:1181–1186
Feng C, Shen SH, Cheng HP, Jing YX, Yanni YG, Dazzo FB (2006) Ascending migration of endophytic rhizobia, from roots to leaves, inside rice plants and assessment of benefits to rice growth physiology. Appl Environ Microbiol 71:7271–7278
Frankenberger WT, Arshad M (1995) Phytohormones in soils: microbial production and function. Marcel Dekker, New York
Goubran FH, Richards D (1979) The estimation of root length in samples and subsamples. Plant Soil 52:77–83
Hirsch R (2000) La Riziculture Malgache Revisitée: Diagnostic et Perspectives (1993–99). Agence Française de Développement, Antananarivo, Annexes, pp 13–14
Ilag LL, Roasles AM, Elazegui FA, Mew TW (1987) Changes in the population of infective endomycorrhizal fungi in a rice-based cropping system. Plant Soil 103:67–73
Johnson BK (1994) Soil survey. In: Final report for the agricultural development component of the Ranomafana National Park Project in Madagascar. Soil Science Dept, North Carolina State University, Raleigh, NC, pp 5–12
Kabir H, Uphoff N (2007) Results of disseminating the System of Rice Intensification with Farmer Field School methods in Northern Myanmar. Exper Agric 43:463–476
Kar S, Varade SB, Subramanyam TK, Ghildyal BK (1974) Nature and growth pattern of rice root system under submerged and saturated conditions. Il Riso 23:173–179
Kirk GJD, Bouldin DR (1991) Speculations on the operation of the rice root system in relation to nutrient uptake. In: Penning de Vries FWT et al (eds) Simulation and systems analysis for rice production. Pudoc, Wageningen, pp 195–203
Kirk GJD, Solivas JL (1997) On the extent to which root properties and transport through the soil limit nitrogen uptake by lowland rice. Eur J Soil Sci 48:613–621
Kronzucker HJ, Siddiqui MY, Glass ADM, Kirk JGD (1999) Nitrate-ammonium synergism in rice: a subcellular flux analysis. Plant Physiol 119:1041–1045
Laulanié H (1993) Le système de riziculture intensive malgache. Tropicultura 11:110–114
Magdoff FR, Bouldin DR (1970) Nitrogen fixation in submerged soil-sand-energy material media and the aerobic-anaerobic interface. Plant Soil 33:49–61
McDonald AJ, Hobbs PR, Riha SJ (2006) Does the System of Rice Intensification outperform conventional best management? A synopsis of the empirical record. Field Crops Res 96:31–36
Mishra A, Salokhe VM (2008) Seedling characteristics and the early growth of transplanted rice under different water regimes. Exper Agric 44:1–19
Morita S, Suga T (1988) Analysis on root system morphology using a root length density model: II—Examples of analysis on rice root systems. Jpn J Crop Sci 57:755–758
Namara R, Bossio D, Weligamage P, Herath I (2008) The practice and effects of the System of Rice Intensification (SRI) in Sri Lanka. Quart J Intl Agric 47:5–23
Obermueller AJ, Mikkelsen DS (1974) Effects of water management and soil aggregation on the growth and nutrient uptake of rice. Agron J 66:627–632
Olk DC, Cassman KG, Schmidt-Rohr K, Anders MM, Mao JD, Deenik JL (2006) Chemical stabilization of soil organic nitrogen by phenolic lignin residues in anaerobic agroecosystems. Soil Biol Biochem 38:3303–3312
Ponnamperuma FN (1984) Effects of flooding on soils. In: Kozlowski TT (ed) Flooding and plant growth. Academic Press, New York, pp 9–45
Randriamiharisoa R (2002) Research results on biological nitrogen fixation with the System of Rice Intensification. In: Uphoff N et al (eds) Assessments of the System of Rice Intensification: proceedings of an international conference, Sanya, China, April 1–4, 2002. Cornell International Institute for Food, Agriculture and Development, Ithaca, NY. http://ciifad.cornell.edu/sri/proc1/sri_33.pdf
Randriamiharisoa R, Barison J, Uphoff N (2006) Soil biological contributions to the System of Rice Intensification. In: Uphoff N et al (eds) Biological strategies for sustainable soil systems. CRC Press, Boca Raton, FL, pp 409–424
Sato S, Uphoff N (2007) A review of on-farm evaluation of System of Rice Intensification (SRI) methods in eastern Indonesia. CAB review. CABI, Wallingford, UK
Schmidt-Rohr K, Mao JD, Olk DC (2004) Nitrogen-bonded aromatics in soil organic matter and their implications for yield decline in intensive rice crops. Proc Natl Acad Sci 101:6351–6354
Senewiratne ST, Mikkelsen DS (1961) Physiological factors limiting growth and yield of Oryza sativa under unflooded conditions. Plant Soil 14:127–145
Sheehy JE, Peng SB, Dobermann A, Mitchell PL, Ferrer A, Yang JC, Zou YB, Zhong XH, Huang JL (2004) Fantastic yields in the System of Rice Intensification: fact or fallacy? Field Crops Res 88:1–8
Sinha SK, Talati J (2007) Productivity impacts of the System of Rice Intensification (SRI): a case study in West Bengal, India. Agric Water Mgmt 87:55–60
Thakur AK, Uphoff N, Antony E (2010a) An assessment of physiological effects of System of Rice Intensification (SRI) practices compared with recommended rice cultivation practices in India. Exper Agric 46:77–98
Thakur AK, Rath S, Roychowdhury S, Uphoff N (2010b) Comparative performance of rice with System of Rice Intensification (SRI) and conventional management using different spacings. J Agron Crop Sci 196:146–159
Turner BL, Haygarth PM (2001) Phosphorus solubilization in rewetted soils. Nature 411:258
Turner BL, Driessen JP, Haygarth PM, McKelvie ID (2003) Potential contribution of lysed bacterial cells to phosphorus solubilisation in two rewetted Australian pasture soils. Soil Biol Biochem 35:187–189
Uphoff N, Randriamiharisoa R (2002) Reducing water use in irrigated rice production with the Madagascar System of Rice Intensification (SRI). In: Bouman BA, Hengsdijk H, Hardy B, Bindraban PS, Thuong TP, Ladha JK (eds) Water-wise rice production. International Rice Research Institute, Los Baños
Witt C, Dobermann A, Abdulrachman S, Gines HC, Guanghuo W, Nagarajan R, Satawatananont S, Son TT, Tan PS, Tiem LV, Simbahan GC, Olk DC (1999) Internal nutrient efficiencies of irrigated lowland rice in tropical and subtropical Asia. Field Crops Res 63:113–138
Yoshida S, Forno DA, Cock JH, Gomez KA (1972) Laboratory manual for physiological studies of rice, 3rd edn. International Rice Research Institute, Los Baños, Philippines
Zhao LM, Wu LH, Li YS, Lu XH, Zhu DF, Uphoff N (2009) Influence of the system of rice intensification on rice yield and nitrogen and water use efficiency with different N application rates. Exper Agric 45:275–286
Acknowledgments
This research, for an M.S. thesis in Crop and Soil Sciences from Cornell University, was conducted by Barison in cooperation with Oloro McHugh, who concurrently gathered data on water management issues, constraints and opportunities with SRI for his M.S. thesis in Biological and Environmental Engineering. Doing parallel studies in agronomy and agricultural engineering with the same on-station and on-farm data sets provided opportunities for cross-checking and cross-fertilization in the research. The findings reported here are from the work of Barison, who acknowledges the enrichment of his research made possible by this cooperation. This article summarizes the findings presented in Barison (2003), for which Fernandes and Uphoff served as advisors.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Barison, J., Uphoff, N. Rice yield and its relation to root growth and nutrient-use efficiency under SRI and conventional cultivation: an evaluation in Madagascar. Paddy Water Environ 9, 65–78 (2011). https://doi.org/10.1007/s10333-010-0229-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10333-010-0229-z