
Abstract
Habitat use by crop-raiding Japanese macaques (Macaca fuscata) was studied in western Japan from December 2005 to February 2006, a food-scarce season. To examine how different vegetation types affect habitat use by monkeys, two crop-raiding troops were compared: the first troop inhabited a habitat involving more wild food resources; the second troop inhabited a habitat providing fewer wild food resources. It was hypothesized that monkeys living in the habitat with fewer wild food resources are more likely to utilize human settlements and areas around them (i.e. adjacent zones), with a dependence on crop foods. Comparisons of observed and expected habitat use frequencies showed that the first troop selected evergreen broad-leaved forests and conifer plantations, and avoided adjacent zones, rice fields, and golf courses. The second troop selected adjacent zones and avoided conifer plantations, pine forests, and deciduous broad-leaved forests. Both troops moved rapidly in avoided habitat types. These results suggest that monkeys living in the habitat with fewer wild food resources are more likely to utilize areas around human settlements during a food-scarce season.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Japanese macaques (Macaca fuscata) utilize various habitat types, such as cool-temperature deciduous forests, warm-temperature evergreen forests, alpine scrubs, artificial conifer plantations, and human settlements including farmlands (Yamagiwa and Hill 1998; Imaki et al. 2000; Izumiyama et al. 2003). Typically, Japanese macaques prefer evergreen and deciduous broad-leaved forests as foraging sites and avoid coniferous plantations (Imaki et al. 2006), while they may use conifer plantations as resting and/or sleeping sites (Chijiiwa 2002), especially under severe physical conditions such as low winter temperatures (Wada and Tokida 1981).
This study examined habitat use by crop-raiding Japanese macaques, with a focus on the use of human settlements and areas around them. Crop damage by Japanese macaques has been increasing for several decades (Watanabe and Muroyama 2005). However, information on the ecology of crop-raiding monkeys, such as habitat use and feeding patterns, is still limited. Previous studies reported that crop-raiding monkeys frequently use forest edges around human settlements throughout the year (Yamada 2004; Imaki et al. 2006), while they also use deciduous broad-leaved forests throughout the year (Imaki et al. 2006) or in both spring and autumn (Yamada 2004). On the other hand, the use of human settlements and/or farmlands by monkeys may differ among study sites of different vegetation types: monkeys living in the habitat dominated by deciduous broad-leaved forests are less likely to damage crops in winter (Hasegawa et al. 1977; Kanamori and Inoue 1993), while monkeys inhabiting the habitat dominated by conifer plantations utilize human settlements from late autumn to early summer (Chijiiwa 2002). These studies suggest that different vegetation types affect habitat use of crop-raiding monkeys, in particular, the use of human settlements including farmlands, which may provide crop foods with high digestibility and also enable efficient foraging (Forthman-Quick 1986; Altmann and Muruthi 1988; Saj et al. 1999), though they may be a significant survival risk to monkeys (Horrocks and Baulu 1994; Newmark et al. 1994; Strum 1994; Hill 2000).
To examine how different vegetation types affect habitat use by monkeys, two crop-raiding troops were compared. One troop inhabited an area, of which a large proportion is covered by evergreen broad-leaved forests, assumed to be a habitat which involves more wild food resources for monkeys, while the other troop inhabited an area covered mainly by conifer plantations and pine forests, assumed to be a habitat which provides fewer wild food resources (Takasaki 1981; Furuichi et al. 1982; Nakagawa et al. 1996). It was hypothesized that monkeys living in a habitat with fewer wild food resources are more likely to utilize human settlements and/or areas around them, with a dependence on crop foods. The study was conducted in winter (December–February) when the differences between the two study sites are the greatest because only evergreen broad-leaved forests provide ample wild food resources (i.e., mature leaves) during this time of year (Nakagawa et al. 1996).
We also examined how habitat type affects travel speed of monkeys. It was predicted that monkeys are more likely to travel slowly in habitat types with more wild food resources to forage and/or with good shelters against severe meteorological conditions.
Methods
Study areas
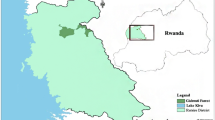
This study was conducted in two areas in western Japan (Fig. 1). One study site was the Kameyama area, located in Kameyama and Suzuka cities (135°45′E, 34°86′N) in Mie Prefecture. The annual mean temperature is 14.5°C, the annual mean rainfall is 1879.5 mm, and the elevation ranges from 50 to 400 m. This area is largely covered by two types of vegetation: evergreen broad-leaved forests dominated by Castanopsis cuspidate, Machilus thunbergii, and Quercus glauca, and coniferous plantations of Cryptomeria japonica and Chamaecyparis obtusa. There are some golf courses covered mostly by grasses and some red pine (Pinus densiflora) clumps. Troops of Japanese macaques are distributed continuously in and around this study site. The other study site was the Nabari area, situated in Nabari city in Mie prefecture and in Uda city in Nara prefecture (136°01′E, 34°34′N). The annual mean temperature is 13.9°C, the annual mean rainfall is 1366.5 mm, and the elevation ranges from 200 to 500 m. This site is covered with coniferous plantations of Cryptomeria japonica and Chamaecyparis obtusa, red pine forests, and deciduous broad-leaved forests dominated by Quercus serrata. The area is located at the far west end of the Chubu-Kinki population of Japanese macaques (Muroyama et al. 1999). Both areas have little snowfall in winter and are located approximately 50 km apart. Crop raiding by monkeys has been reported for several years in both areas (Yamada 2004; Kameyama city office personal communication). Farm practices in the two study areas were for household consumption, and there were no substantial differences in crops between them during study period. In both areas, human settlements included farmlands for crops (e.g. Raphanus sativus, Brassica rapa, etc.) and fruit trees such as Diospyros kaki and Citrus spp., all of which were raided by monkeys during this time of year. Rice fields provided no foods for monkeys, because rice fields were harvested in autumn and left unplanted over the winter.

Map of the location of the Kameyama and Nabari areas
Study subjects
Two troops were followed: the KH troop in the Kameyama area and the NB troop in the Nabari area. The KH troop was composed of 18 members at the beginning of the study (7 adult females, 1 adult male, 1 young female, 1 young male, 6 juveniles, and 2 infants). The NB troop was composed of 62 members at the onset of the study (22 adult females, 3 adult males, 3 young females, 4 young males, 23 juveniles, and 7 infants).
After local government authorization to capture monkeys was granted, in each study area one adult female in each troop was captured with a box trap (60 × 80 × 120 cm). Captured females were immobilized by ketamine hydrochloride (10 mg/kg), weighed, measured, fitted with a radio collar (M2950, ATS) and then released. Manipulation and care of animals followed the Guideline for Field Research of Non-human Primates of Primate Research Institute, Kyoto University. Each female returned to her troop in a few days—this was confirmed by direct observations and by vocalizations of troop members around location points found by radio-tracking.
Analysis
Home range
From December 2005 to February 2006, we followed each monkey troop from 07:00 to 17:00 for 5–7 days each month. Troop locations were obtained by triangulation using a hand-held three-element Yagi antenna (Model F147-3FB, AF Antronics) and a portable receiver (VR-500, Standard). Locations of the troops were recorded once an hour. Location errors due to animal movement were minimized by locating fixes from different points within 15 min (Börger et al. 2006). Almost all of the locations identified by radio-telemetry were confirmed by direct observation or monkey vocalizations. When following the troops, we recorded plant species and its parts that monkeys ate at the moment and between the times of fixing location.
Locations of the troops were plotted and analyzed with a geographic information system (GIS; Arc GIS 8.3; Environmental Systems Research Institute, Redlands, CA, USA). Home range size of each troop was estimated by use of the 95% minimum convex polygon (MCP) in the animal movement extension for ArcView (Hooge and Eichenlaub 1997).
Habitat selection
A vegetation map of each of the study areas was made from the 1:50000 vegetation map published by Ministry of the Environment of Japan in 1999, correcting the map according to data collected by a field vegetation survey and aerial photographs by the Geographical Survey Institute. Seven habitat types were identified as available in the Kameyama area: evergreen broad-leaved forests, conifer plantations, adjacent zones, human settlements, rice fields, golf courses, and others. In the Nabari area six habitat types were identified: deciduous broad-leaved forests, conifer plantations, pine forests, adjacent zones, human settlements, and others. In the Kameyama area, rice fields were distinguished from human settlements as different habitat type because rice fields lay along the valleys in hilly terrain far from human settlements. In contrast, in the Nabari area rice fields were categorized as human settlements because rice fields were within human settlements.
Because previous studies reported that areas surrounding human settlements may provide particular functions for crop-raiding monkeys irrespective of their actual habitat type category (Yamada 2004), we demarcated buffer zones of 100 m from the edge of human settlements using GIS, and defined such zones as adjacent zones, irrespective of actual habitat type. Adjacent zones in Kameyama area consisted of rice fields, conifer plantations, evergreen broad-leaved forests, abandoned farmland (i.e. grassland or wasteland), golf courses, and others. In Nabari area, adjacent zones consisted of conifer plantations, pine forests, abandoned farmlands, deciduous broad-leaved forests, and others.
To evaluate habitat selection we defined the borders of each of the two study areas by plotting troop locations and obtaining a 95% MCP. The proportions of each habitat type in the MCP were regarded as availability. We superimposed the locations of each troop on each study area and evaluated the distribution of troop locations in each habitat type in relation to their availability. Selection of habitat types was assessed with a χ 2 test. The level of significance was P < 0.05. We also calculated Manly’s standardized selection ratio (α) to determine if monkeys select particular habitat types. The formula for this index is:
where α i is the selective index for habitat type i, n i is the relative availability of habitat i, r i is the relative use of habitat i, n j is the relative availability of habitat j, r j is the relative use of habitat j, and m is the number of habitat types. Values of α i range from 0 with no probability of selecting a given habitat type to 1, meaning the habitat type will always be selected. After that, we calculated the Bonferroni confidence interval for each habitat type using values of α i and standard errors of α i to determine the selection (+, −, or neutral) of that habitat type (Manly et al. 2002). Statistical significance by using the Bonferroni confidence interval is determined by both of the values of α i and standard errors of α i .
To examine the effects of snowfall on the use of conifer plantation, we assessed the selection with a χ 2 test and the Bonferroni confidence interval, and calculated Manly’s standardized selection ratio (α) for the data of points selected for conifer plantations when it was snowing (snowing points at the time of observation) and those when it was not snowing.
We also calculated travel speeds in each of the habitat types based on the hourly location of points recorded on their travel routes (KH troop, n = 156; NB troop, n = 159). When monkeys traveled completely within the same habitat type between two points (KH troop, n = 78; NB troop, n = 69), we used the distance between two points for calculations of travel speed (m/h) in that habitat type. When monkeys traveled in multiple habitat types between two points, we used the distance between two points as data for travel speed in the habitat type where the percentage of the distance traveled within it was equal to or more than 50% (KH troop, n = 68; NB troop, n = 87). If the percentage of the distance traveled within any of the habitat types was less than 50%, we discarded the data because of poor quality (KH troop, n = 10, NB troop, n = 3). We used Kruskal–Wallis one-way analysis of variance to examine differences in travel speed between habitat types, and Spearman’s correlation coefficient between Manly’s α and travel speed except for other habitat type to examine how habitat type affects travel speed of monkeys.
Results
Home range and habitat selection
During the study period, 186 fixes were obtained for the KH troop and 185 fixes were obtained for the NB troop. Home range sizes by MCP of the KH troop and the NB troop were 5.4 and 7.6 km2, respectively (Fig. 2).

Vegetation map of each study site, the home range of each troop by MCP, and location points of each troop. Vegetation map and home range in a the Kameyama area and b the Nabari area. Location points, human settlements, and home range in c the Kameyama area and d the Nabari area
As shown in Table 1, the most prominent habitat type in the MCP area of the KH troop was evergreen broad-leaved forests (33%) followed by golf courses (23%), rice fields (15%), conifer plantations (13%), and adjacent zones (10%). In this troop, observed habitat use frequencies differed significantly from expected frequencies based on habitat availability (χ 2 = 168.05, df = 6, P < 0.001). The troop selected evergreen broad-leaved forests and conifer plantations, and avoided golf courses, rice fields, and adjacent zones. Adjacent zones in the Kameyama area consisted mainly of rice fields (65.6%) (Table 2), which may affect monkeys’ avoidance of adjacent zones because of higher risk in the central open area of rice fields (Table 1). Manly’s α indicated that the KH troop was less likely to use human settlements and adjacent zones than evergreen broad-leaved forests and conifer plantations.
In the MCP area of the NB troop, adjacent zones (30%) were most followed by conifer plantations (28%), pine forests (21%), and human settlements (17%). Deciduous broad-leaved forests covered only 3% of this area. Habitat use frequencies observed differed significantly from expected frequencies based on habitat availability (χ 2 = 64.63, df = 7, P < .001). The troop predominantly selected adjacent zones and avoided conifer plantations, pine forests, and deciduous broad-leaved forests. Adjacent zones in the Nabari area consisted mainly of conifer plantations (48.9%) and pine forests (23.1%) (Table 2), but both of these were habitat types avoided outside the adjacent zones (Table 1). This indicates that the NB troop selected adjacent zones irrespective of habitat types involved. Manly’s α indicated that the NB troop was more likely to use adjacent zones and human settlements than conifer plantations, pine forests, and deciduous broad-leaved forests.
If monkeys use conifer plantations as shelter, this habitat type could be more likely to be selected under severe physical conditions such as snowing. This was the case for the KH troop, which selected conifer plantations when it was snowing (n = 48), but not when it was not snowing (n = 138) (Bonferroni confidence interval, statistically significant). Manly’s α indicated no differences between two conditions for both troops (KH troop: snowing points at the time of observation (n = 48), Manly’s α = 0.213, points with not snowing (n = 138), Manly’s α = 0.213; NB troop: snowing points at the time of observation, n = 15, Manly’s α = 0.128, points with not snowing, n = 170, Manly’s α = 0.066), however. This was because statistical significance by using Bonferroni confidence interval was determined by both of the values of α i and standard errors of α i and the values of standard errors of α i differed between two conditions.
Travel speeds in habitat types
Mean travel speed was 150.7 ± 142.3 m for the KH troop (range 2.2–909.6 m) and 158.3 ± 142.4 m for the NB troop (range 1.6–820.5 m). Travel speed differed between habitat types for both troops (Table 3, Kruskal–Wallis test, KH troop: χ 2 = 19.927, df = 6, P < 0.003; NB troop: χ 2 = 11.297, df = 5, P < 0.05). To eliminate the possibility that habitat types with small sample sizes affected the results of the Kruskal–Wallis test, the test was run without the habitat types with N < 10 and the difference of travel speeds between habitat types was found again (Kruskal–Wallis test, KH troop: χ 2 = 4.312, df = 3, P = 0.23; NB troop: χ 2 = 7.973, df = 2, P < 0.02).
The KH troop traveled quickly in adjacent zones and on golf courses, both of which were avoided habitat types, and moved relatively slowly in selected habitat types such as evergreen broad-leaved forests and conifer plantations. Travel speed in rice fields and human settlements was the lowest, although this habitat type was avoided or neutral.
The NB troop moved most rapidly in conifer plantations, which was an avoided habitat type. In contrast, travel speed in human settlements and adjacent zones, both of which were selected or neutral habitat types, was relatively slow. This troop also traveled rather slowly in the other avoided habitat types, for example deciduous broad-leaved forests and pine forests.
There was no significant correlation between the Manly’s α and travel speed for the KH troop (Spearman test, ρ = −0.23, n.s., n = 6). For the NB troop, no significant correlation was found between the Manly’s α and travel speed, though the relationship was slightly negative (Spearman test, ρ = −0.5, n.s., n = 5).
Food items
During the study periods, we observed that monkeys of the KH troop fed on fruits of kaki (Diospyros kaki) in human settlements and adjacent zones planted by local people, but they raided no crops. Monkeys also foraged on acorns of Castanopsis cuspidate, seeds of Rhus spp, beans of Pueraria lobata, fruits of Rubus buergeri, Aucuba japonica, and Euria japonica, and leaves of Euria japonica, Rumex acetosa, and Taraxacum officinale in and around evergreen broad-leaved forests and conifer plantations.
In contrast with the KH troop, we observed frequent crop raiding by NB troop monkeys when they used human settlements at the moment and between the times of fixing location. Monkeys of the NB troop moved back and forth between adjacent zones and human settlements during crop raiding. Monkeys damaged crops, such as Japanese radishes (Raphanus sativus), turnips (Brassica rapa), scallions (Allium fistulosum), carrots (Daucus carota), onions (Allium cepa), nappa cabbages (Brassica rapa var. amplexicaulis), and broccoli (Brassica oleracea), all of which were general vegetables cultivated by local people. However, monkeys of the NB troop were not observed to feed in adjacent zones, and this suggested that they did not use this habitat type as feeding site during the study period. In human settlements, including just outside of forest edges, monkeys fed on fruits of kaki (Diospyros kaki), chestnut (Castanea crenata), and kiwi (Actinidia chinensis), fruits and leaves of Citrus spp., and buds of figs (Ficus carica), all of which were planted by local people. Furthermore, monkeys damaged and consumed plants, for example Nandina domestica or Camellia japonica, in the gardens. Monkeys of the NB troop also foraged on wild foods such as beans of Pueraria lobata and Wisteria barachybotrys, fruits of Rubus buergeri and Euria japonica, and leaves of Pueraria lobata and Rumex acetosa in and around pine forests and conifer plantations.
Discussion
Each of the two study troops showed preferences for particular habitat types. The KH troop used evergreen broad-leaved forests more frequently than expected, as previously reported for Japanese macaques (Imaki et al. 2006; Toda 2007), used habitat types around human settlements less frequently, i.e., adjacent zones mostly composed of rice fields (Table 2), and avoided open space such as rice fields and golf courses outside the adjacent zones. The KH troop selected positively conifer plantations which are expected to be a habitat with fewer wild food resources. This may be partly because monkeys stayed inside conifer plantations for use as shelter (Wada and Tokida 1981), as indicated by the observation that the KH troop selected this habitat type when it was snowing but not when it was not snowing.
In contrast, the NB troop selected adjacent zones and avoided conifer plantations, pine forests, and even deciduous broad-leaved forests outside the adjacent zones. Most of the adjacent zones consisted of conifer plantations and pine forests (Table 2), and these were avoided habitat types (Table 1). Because those habitat types may provide fewer wild food resources during the study period, monkeys of the NB troop do not seem to have used adjacent zone as feeding sites. Thus, the high score of Manly’s α for adjacent zones indicated that the NB troop may stay longer around human settlements to raid crop foods, irrespective of habitat types around them. These results are consistent with the prediction that monkeys living in a habitat with fewer wild food resources are more likely to select human settlements and/or adjacent zones. Monkeys of the NB troop consumed various cultivated foods such as crops, fruits, and ornamental plants in gardens. The dependence on such cultivated foods by this troop would lead to frequent utilization of areas around human settlements.
As predicted, travel speed of both troops was slow in selected habitat types and quick in avoided habitat types, with some exceptions. However, no significant correlation between Manly’s α and travel speed was found. This may be partly because the degree of habitat type selection was not correlated strictly with travel speed and/or because number of habitat types in this study was rather small for statistics (KH troop, n = 6; NB troop, n = 5).
Travel speed in human settlements was the lowest for all habitat types with one exception of rice fields for the KH troop. This suggests that human settlements are large food patches for crop-raiding monkeys, and the monkeys stay for a period of time or move slowly when they feed, even if human settlements are not positively selected. The KH troop moved slowly in rice fields, although this habitat type was avoided. Monkeys of the KH troop were observed frequently to use the boundary between forests and rice fields to feed on plants such as Rumex acetosa and Taraxacum officinale on the edge and to rest there. This boundary utilization as a feeding site may lead to monkeys’ slow travel in rice fields. Although human settlements and the boundary between forests and rice fields were used as feeding site, central parts of those open areas were less likely to be used by monkeys, probably because of the high risk of being chased away by humans and dogs (Cowlishaw 1997). This may lead to low scores of Manly’s α in human settlements and rice fields, because areas substantially used by monkeys were much limited.
We have reported the results for the analyses of travel speeds when monkeys traveled in single or multiple habitat types between two location points. We have also analyzed travel speeds only when monkeys traveled completely within the same habitat type between two location points (KH troop, n = 78; NB troop, n = 69), but the results were unclear because five of the seven habitat types for the KH troop and four of the six habitat types for the NB troop had small sample sizes (N ≤ 5). Such small sample sizes in these habitats may result from the following possible reasons that are not mutually exclusive. One reason is that monkeys traveled longer in a habitat type and then travel lines are more likely to include multiple habitat types. Another is that when the area of a habitat type was relatively small, travel lines were less likely to be contained within the habitat type. We need many more sample sizes in each habitat type to enable more detailed analysis of travel speeds.
Data in this study were collected for two troops with different troop size during a limited period in 1 year only, and thus some important factors affecting habitat use by monkey troops were not examined. For example, this study did not analyze the effect of annual variations in availability of food resources in a habitat on use of the habitat by monkeys. Previous studies reported that home range use by Japanese macaques was affected by annual variations in food resources in the habitat (Hill and Agetsuma 1995; Tsuji and Takatsuki 2004, 2009). In these study areas a super-abundant harvest of acorns in autumn might lead to high availability of fallen acorns in winter, and could thereby affect use of broad-leaved forests by monkeys.
Difference in the sizes of the study troops could also affect the results of this study, because troop sizes are related to the area of broad-leaved forests in the home range (Takasaki 1981; Muroyama 2008). In fact, the KH troop had a smaller troop size (n = 18) and a much larger area of broad-leaved forests per animal (9.88 ha/animal, not including the forests within adjacent zones) than the NB troop (n = 62, 0.00 ha/animal). This means that the effects of different habitat types between the KH and NB troops were emphasized by the different troop sizes. Furthermore the area of broad-leaved forest per animal for the KH troop was larger than the average (1.9 ha/animal) predicted by Takasaki (1981).
The results of this study suggest that crop-raiding monkeys which have a home range involving habitat types with ample wild food resources (e.g., broad-leaved forests) are more likely to select these habitat types and avoid habitat types around human settlements. In contrast, monkeys with fewer wild food resources in their home range, i.e. a very small area of broad-leaved forests, are more likely to select habitat types around human settlements such as adjacent zones, but not broad-leaved forests. These findings suggest that some proportion of broad-leaved forests in the home range may be essential to prevent monkeys from depending on crop foods in human settlements, though we need more studies on this issue under various types of vegetation for conclusive proof.
References
Altmann J, Muruthi P (1988) Differences in daily life between semi-provisioned and wild-feeding baboons. Am J Primatol 15:213–221
Börger L, Franconi N, De Michele G, Gantz A, Meschi F, Manica A, Lovari S, Coulson T (2006) Effects of sampling regime on the mean and variance of home range size estimates. J Anim Ecol 75:1393–1405
Chijiiwa A (2002) Movement patterns and habitat use of Japanese macaque (Macaca fuscata) troops in a conifer-highly planted region (in Japanese with English summary). Master’s thesis. Doshisha University, Kyoto, Japan, p 65
Cowlishaw G (1997) Trade-offs between foraging and predation risk determine habitat use in a desert baboon population. Anim Behav 53:667–686
Forthman-Quick DL (1986) Activity budgets and the consumption of human foods in two troops of baboons, Papio anubis, at Gilgil, Kenya. In: Else JC, Lee PC (eds) Primate ecology and conservation. Cambridge University Press, Cambridge, pp 221–228
Furuichi T, Takasaki H, Sprague DS (1982) Winter range utilization of a Japanese macaque troop in a snowy habitat. Folia Primatol (Basel) 37:77–94
Hasegawa T, Hasegawa M, Tsuchiya S (1977) Takagoyama shuhen no nihonzaru no genjou Hogo no kanten kara (in Japanese). Nihonzaru 3:50–61
Hill CM (2000) Conflict of interest between people and baboons: crop raiding in Uganda. Int J Primatol 21:299–315
Hill DA, Agetsuma N (1995) Supra-annual variation in the influence of Myrica rubra fruit on the behavior of a troop of Japanese macaques in Yakushima. Am J Primatol 35:241–250
Hooge PN, Eichenlaub B (1997) Animal movement extension to ArcView. Ver.1.1: Alaska Biological Science Center. U.S. Geological Survey, Anchorage
Horrocks JA, Baulu J (1994) Food competition between vervets (Cercopithecus aethiops sabaeus) and farmers in Barbados: implications for management. Rev Ecol (Terre et Vie) 49:281–294
Imaki H, Koganezawa M, Okumura T, Maruyama N (2000) Home range and seasonal migration of Japanese monkeys in Nikko and Imaichi, central Honshu, Japan. Biosph Conserv 3:1–16
Imaki H, Koganezawa M, Maruyama N (2006) Habitat selection and forest edge use by Japanese monkeys in the Nikko and Imaichi area, central Honshu, Japan. Biosph Conserv 17:87–96
Izumiyama S, Mochizuki T, Shiraishi T (2003) Troop size, home range area and seasonal range use of the Japanese macaque in the Northern Japan Alps. Ecol Res 18:465–474
Kanamori H, Inoue J (1993) Shimaneken ni okeru nihonzaru no seisoku, higai, oyobi taisaku no jittai (in Japanese). For Pests 42:6–13
Manly B, McDonald L, Thomas D, McDonald T, Erickson W (2002) Resource selection by animals: statistical design and analysis for field studies. Kluwer, Dordrecht
Muroyama Y (2008) Conservation of Satoyama and damage management: Japanese macaques (in Japanese). In: Takatsuki S, Yamagiwa J (eds) Japanese mammalogy: medium- and large-sized mammals, and primates. University of Tokyo Press, Tokyo, pp 427–452
Muroyama Y, Torii H, Maekawa S (1999) The distribution and management status of wild Japanese macaques (Macaca fuscata fuscata) in Kinki district, Japan (in Japanese). Wildl Forum 5:1–15
Nakagawa N, Iwamoto T, Yokota N, Soumah AG (1996) Inter- regional and inter-seasonal variations of food quality in Japanese macaques: constraints of digestive volume and feeding time. In: Fa JE, Lindburg DG (eds) Evolution and ecology of macaque societies. Cambridge University Press, Cambridge, pp 207–234
Newmark WD, Manyanza DN, Gamassa DGM, Sariko HI (1994) The Conflict between Wildlife and local people living adjacent to protected areas in Tanzania—human density as a predictor. Conserv Biol 8(1):249–255
Saj TL, Sicotte P, Paterson JD (1999) Influence of human food consumption on the time budget of vervets. Int J Primatol 20:977–994
Strum SC (1994) Prospects for management of primate pests. Rev Ecol (Terre et Vie) 49:295–306
Takasaki H (1981) Troop size, habitat quality, and home range area in Japanese macaques. Behav Ecol Sociobiol 9:277–281
Toda H (2007) Crop damages caused by spatial movements of Japanese macaques in the Kameyama Hills of Mie Prefecture, Japan (in Japanese with English summary). Geogr Rev J 80:614–634
Tsuji Y, Takatsuki S (2004) Food habits and home range use of Japanese macaques on an island inhabited by deer. Ecol Res 19:381–388
Tsuji Y, Takatsuki S (2009) Effects of yearly change in nut fruiting on autumn home-range use by Macaca fuscata on Kinkazan Island, northern Japan. Int J Primatol 30:169–181
Wada K, Tokida E (1981) Habitat utilization by wintering Japanese monkeys (Macaca fuscata) in the Shiga Heights. Primates 22:330–348
Watanabe K, Muroyama Y (2005) Recent expansion of the range of Japanese macaques, and associated management problems. In: Paterson JD, Wallis J (eds) Commensalism and conflict: The primate-human interface. Special topics in primatology 4. American Society of Primatologists, Norman, pp 313–331
Yamada A (2004) Effects of food availability in croplands and forests on habitat selection of crop-raiding macaques (in Japanese). Master’s thesis. Kyoto University, Kyoto, Japan, p 30
Yamagiwa J, Hill DA (1998) Intraspecific variation in the social organization of Japanese macaques: past and present scope of field studies in natural habitats. Primates 39:257–273
Acknowledgments
We are grateful to the city offices of Kameyama, Suzuka, Nabari, and Uda, and to the local residents who cooperated with our study. We thank the members of Seminar on Ecology and Social Behavior at Primate Research Institute for their valuable comments. We are very thankful to David Sprague, Naoki Agetsuma, and one anonymous reviewer for their invaluable comments to improve the manuscript. The research and care of monkeys followed the Guideline for Field Research of Non-human Primates of Primate Research Institute, Kyoto University. This study was financed by the Twenty-first Century COE Program, Kyoto University and Grand-in-Aid for Scientific Research (C) 18580268.
Author information
Authors and Affiliations
Corresponding author
About this article
Cite this article
Yamada, A., Muroyama, Y. Effects of vegetation type on habitat use by crop-raiding Japanese macaques during a food-scarce season. Primates 51, 159–166 (2010). https://doi.org/10.1007/s10329-009-0183-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10329-009-0183-9