
Abstract
We presented two chimpanzees with a task in which they were required to pull each end of a rope simultaneously to drag blocks supporting food into reach. The chimpanzees did not succeed in initial tests. They did not immediately understand the necessity for cooperation, and they did not adjust their behavior to work with the partner. However, the frequency of success gradually increased as the number of sessions increased and the task was varied. They began to look at the partner frequently, wait if the partner was not holding the rope, and pull the rope in synchrony with the partner. However, they did not use interactive behaviors or eye contact to synchronize their behavior. One chimpanzee was then paired with a human partner in the same situation. After initial failures, the chimpanzee began to solicit the human partner for cooperation: looking up at his face, vocalizing, and taking the partner’s hand. When this chimpanzee was again paired with the chimpanzee partner, no soliciting behavior was observed. Thus, the chimpanzees could learn to coordinate their behavior through trial and error. Communicative behavior emerged during the task, but the communication differed according to the identity of the partner.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Cooperative behavior by nonhuman primates, including cooperative hunting by wild chimpanzees (Boesch and Boesch 1989), cooperative parenting by some species of New World monkeys (Garber 1997), and reciprocal grooming in many species of monkeys (Muroyama 1991), has been documented in the wild. In contrast, only a few experimental studies have examined cooperative behavior in more controlled situations.
A pioneering study was conducted by Crawford (1937). A task was introduced in which a chimpanzee had to pull a rope to drag a box that contained food. The box was too heavy for one chimpanzee to pull; thus, two chimpanzees had to cooperate by each pulling a rope to drag the box. In early trials, the two chimpanzees did not cooperate. Success occurred after active teaching by the experimenter, through which each chimpanzee learned to pull the rope upon a verbal cue from the experimenter. Once cooperation was established in this way, the chimpanzees cooperated even without verbal cues. The chimpanzees even began to solicit the partner when necessary, by touching, placing an arm over the partner’s body, or vocalizing.
Decades later, Chalmeau and colleagues carried out experimental studies of cooperation in three nonhuman primate species. They first examined the chimpanzee (Chalmeau 1994; Chalmeau and Gallo 1996a, b). A specially constructed fruit distributor was presented to a group of captive chimpanzees; two individuals had to pull a handle connected to the device simultaneously to make a fruit fall into the enclosure. The dominant male and an infant produced most of the pulling responses, and the male obtained nearly all the fruit. This male glanced increasingly frequently toward the partner, suggesting that he learned to cooperate with the infant. On the other hand, the infant did not reliably check the partner; the authors interpreted the infant’s behavior as partially linked to play activities.
Chalmeau et al. (1997a) then tested orangutans. Two orangutans pulled the handles simultaneously, but as in the chimpanzees, only one individual did all the monitoring and coordinating. The more passive partner was a young female, lacking complete sensorimotor development or possibly presenting a psychomotor retardation, according to the authors. Thus, there was asymmetry in the pairs of individuals who performed the cooperative responses.
The third species tested was the capuchin monkey (Cebus apella) (Chalmeau et al. 1997b). The monkeys sometimes succeeded in the task, but the successes appeared to be the fortuitous outcome of the monkeys pulling the handle randomly. However, Mendres and de Waal (2000) utilized the paradigm pioneered by Crawford (1937) and showed that capuchin monkeys understood the role of the partner. Therefore, the nature of the task may influence the animals’ performance.
The method used by Crawford was re-introduced to chimpanzees by Povinelli and O’Neill (2000). They trained a pair of chimpanzees to cooperatively pull a rope attached to very heavy boxes to obtain food. The learning process was not described in detail; rather, attention was focused on the behavior of a trained chimpanzee paired with a naïve partner. They found no evidence of the use of gestures by the experienced individual to instruct the naïve partner.
Melis et al. (2006) used our method, described in this paper, to test cooperation in chimpanzees. They showed that chimpanzees could recruit a partner to collaborate when necessary, and that they recruited the more effective of two partners on the basis of their past experience with each partner. Again, the learning process was not the focus of study.
Following these lines of research, further tests with well-matched chimpanzee pairs in another type of test are of interest for three reasons. First, additional tests are required to observe the process of spontaneous learning, without human intervention. Second, changes in the difficulty of a task may elicit different behavioral characteristics from the subjects. The tasks used by Crawford (1937) and Chalmeau (1994) did not require complete understanding of how to cooperate with another individual, because the subjects could solve the task by randomly pulling the rope. If the task requires true synchronization of behavior between partners, they may be more likely to show more communicative behavior. Third, communicative behavior may vary across different pairs. We are especially interested in whether a subject’s behavior changes when paired with a human whose communicative behavior differs from that of conspecifics.
Here, we describe a task designed to test chimpanzee cooperative behavior. Two chimpanzees of similar size and age participated, and we examined how they learned to coordinate and synchronize their behavior. In addition, to investigate the emergence of spontaneous soliciting behavior, one chimpanzee of the pair and a human were tested in the same experimental situation.
Methods
Subjects and housing conditions
The subjects were two young female chimpanzees (Pan troglodytes) cared for at the Great Ape Research Institute (GARI) of Hayashibara Biochemical Laboratories, Inc. Tsubaki and Mizuki, were 7 years and 1 month old and 6 years and 3 months old, respectively, at the start of the first test in March 2003. They were born in captivity at another institution. Tsubaki was mother-reared until a few months before she was moved to GARI, and Mizuki was hand-reared from a few days after birth. They arrived at GARI in January 1999 and lived together with three other individuals as a group in a large outdoor compound of approximately 7,400 m2. The compound contained natural forest, a pond, and a 13-m-high climbing structure with attached indoor rooms and a smaller compound (Idani and Hirata 2006). The chimpanzees were fed various fruits and vegetables three to ten times a day. Water was freely available, and the chimpanzees were never food-deprived.
Apparatus
Two identical styrene blocks, a vinyl chloride pipe, and a cotton rope were used as materials for the apparatus. The styrene block was 19 cm wide, 19 cm long, and 11 cm tall. A 6 × 10-cm hole, through which the rope passed, was bored from the right to left side of the block. The pipe, 2 cm in diameter and 160 cm long, connected the two blocks. The pipe was longer than the arm span of the subjects with arms outstretched horizontally to the side. The rope, 5 mm in diameter, varied in length according to the test conditions.
All training and test sessions took place in an experimental booth. The booth was heptagonal in cross-section, 7.6 m2 in area, and 5 m tall. The walls of the booth had a 12-cm-high opening at the bottom that was divided into two 5-cm-high openings by a horizontal metal bar, 2 cm in diameter. The chimpanzees in the booth could reach with a hand through either of these two openings.
Training
Each chimpanzee was individually trained to pull both ends of the rope by herself. One block was placed outside the booth at a distance of 70 cm from the opening; this was beyond the reach of a chimpanzee when introducing her hand through the 5-cm opening. Food (a piece of sugarcane or sweet potato) was placed on the block. A rope was passed through the hole in the block, and the two ends were fed into the booth through the opening. The chimpanzee could drag the block within reach to obtain the food by pulling both ends of the rope. If she pulled only one end of the rope, she would obtain only the rope, and the block would remain out of reach. One training session consisted of five to ten trials; both chimpanzees succeeded on 100% of trials from the second session on, until all ten sessions were completed.
Initial test
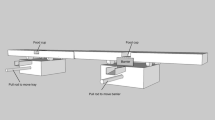
Six days after the training sessions ended, the first test with the two chimpanzees together was conducted. Two blocks identical to that used in training were connected with the 160 cm-long pipe and placed outside the booth, 70 cm from the opening. Food (a piece of sugarcane or sweet potato) was placed on each block. A rope was passed through each block and each end was fed into the booth through the opening so that 10 cm of rope was inside the booth (Fig. 1).

A picture of the experimental apparatus
The two chimpanzees were brought to the booth together before testing began. While one experimenter (SH) prepared the apparatus, another experimenter (KF) stayed in the booth with the chimpanzees to keep them occupied. KF called the chimpanzees to an area about 2.5 m from where the apparatus was being set up, and fed them small pieces of fruit while remaining in front of them blocking their access. When the apparatus was ready, the second experimenter stopped feeding the chimpanzees and moved aside to allow them to behave freely. A trial ended when the chimpanzees obtained food from the blocks or obtained only the rope. At the end of a trial, the first experimenter again prepared the apparatus for use while the second experimenter kept the chimpanzees occupied. Each session consisted of ten trials, and only one session was conducted each day. A total of three sessions was conducted.
Learning phase
Because the chimpanzees were never successful in the initial test, modifications were introduced by changing the length of the rope ends inside the booth: long (130 cm), medium (80 cm), slightly short (40 cm), and short (10 cm, same as in the initial test). All other conditions were the same as in the initial test. In the first three treatments, one end of the rope could be pulled before the other without the other end disappearing from the booth. In addition, one chimpanzee could pull both ends of the rope by bringing one end closer to the other and therefore succeed individually. We ran 30, 10, 10, and 10 sessions with the long-, medium-, slightly short-, and short-rope treatments, respectively, in succession, at a frequency of 1 session per day, and 0–2 sessions per week. Each session consisted of ten trials.
Waiting room test
To reduce any effect of the experimenter remaining in the booth with the chimpanzees, another treatment was added after the completion of the short-rope treatment described above. The chimpanzees were kept in a room adjacent to the experimental booth while the apparatus was prepared. This waiting room was 4.3 m2 in area and 2.5 m high, and separated from the booth by an electrically controlled door. When the apparatus was ready, the experimenter opened the door to allow the chimpanzees to enter the booth. This treatment was inserted as probe trials among “normal” short-rope treatment trials that served as baseline trials. Thus, after two or three trials during which the experimenter stayed in the booth, the experimenter exited the booth, called the chimpanzees into the waiting room, and a probe trial ensued. A total of 30 sessions consisting of 10–12 trials were conducted at a frequency of 1 session per day and 0–2 sessions per week. During the first five sessions, there were three waiting room probe trials; during the subsequent 25 sessions, there were 4 waiting room probe trials.
Test with a human partner
Nine months after the waiting room tests, another test was introduced in which an experimenter (SH) acted as a cooperative partner with a chimpanzee. Only Mizuki was used as a subject because Tsubaki had become pregnant and her motivation to participate was variable. SH had extensive prior contact with Mizuki over 3 years, including feeding, playing, body checks, and some training for studies. However, Mizuki had never been trained in a cooperative situation with a human prior to this test.
The test procedure was similar to that of the initial test. Between trials, the experimenter (partner) and Mizuki remained in the booth away from the apparatus, and the second experimenter stood in front of them while another assistant prepared the apparatus. When the apparatus was ready, the second experimenter moved aside so that the partner and Mizuki could move freely.
There were two behavioral conditions for the partner. In the first condition, immediately after the second experimenter moved aside, the partner moved toward the apparatus simultaneously with Mizuki, and adjusted his timing to pull the rope with Mizuki. In the second condition, the partner remained still for 2 s after the second experimenter moved aside, unless Mizuki showed any soliciting behavior toward him. The partner started to approach the rope after 2 s had elapsed. In both conditions, the partner did not initiate any interactions with Mizuki. The two conditions were performed in alternate trials and one session consisted of ten trials. If they succeeded in dragging the blocks to the booth, the partner took the food from the block in front of him and placed it in his pocket. Four sessions were conducted, at intervals of 1–4 days.
On the same day the fourth session ended, a final session was conducted. The first six trials were conducted with Mizuki paired with Tsubaki. The next three trials were conducted with Mizuki paired with the human partner (SH). There then followed another three trials of Mizuki paired with Tsubaki, before the session was terminated due to the unstable behavior of Tsubaki.
Scoring
All test sessions were recorded using three video cameras, and several performance indicators were scored from the videotapes. A pull was defined as when a subject took the rope and moved it towards herself. Glances were defined as abrupt head movements in the direction of the partner from the time the experimenter moved aside. A wait was defined as when a subject took the rope, glanced at the partner who had not yet arrived at the rope, and did not pull the rope herself.
There were four possible sequences of behaviors associated with glancing. The first was glancing at the partner who held the rope and then pulling the rope herself. The second was glancing at the partner who did not hold the rope and then pulling the rope herself. The third was glancing at the partner who did not hold the rope and then waiting. The fourth was glancing at the partner who did not hold the rope and then approaching the other end of the rope to attempt the task alone.
There were also four possible outcomes of a trial. The first was complete success, where both subjects pulled the rope simultaneously, dragged the two blocks within reach, and obtained the foods on the blocks. The second was partial success, where both of the subjects pulled the rope, but only one block was dragged within reach because of mistimed pulling. The third was monopolization, where one of the subjects pulled both ends of the rope alone and obtained the food from both blocks. The fourth was failure, where only one subject pulled only one end of the rope, so that the other end was pulled outside the booth, and neither block was brought within reach.
Any interactions by the subject (e.g., taking the partner’s hand) were noted. In addition, the latency from the time the experimenter moved aside until the time the chimpanzees started to move was measured to an accuracy of 0.5 s. The latency to arrival at the end of the rope was also measured. The first author (SH) scored all of the sessions, and an assistant unfamiliar with the purpose of the study analyzed 33% of the data to assess reliability from a second-by-second scoring of the occurrence of glancing, pulling and waiting, along with the latency data. The observed agreement was 95.6% (Cohen’s kappa of 0.83) for glances and 99.2% (Cohen’s kappa of 0.97) for pulling and waiting. The agreement for latency was 94.7%.
Results
Initial test
The chimpanzees were never successful in the initial test. In all 30 trials, only one end of the rope was pulled by one of the two chimpanzees. Tsubaki pulled the rope and failed in the first, second, fourth, seventh, and ninth trials of the first session. Mizuki pulled the rope and failed in all other trials in the first session and all trials in the second and third sessions. Mizuki showed no glancing behavior during these trials. Tsubaki glanced at Mizuki who was pulling the rope twice, in the third and tenth trials.
Learning phase
The chimpanzees initially failed, but gradually became successful during the course of the learning phase. In trials 1 and 2 of the first session with the long rope, Mizuki pulled both rope ends alone and was successful; Tsubaki did not approach the rope. In the third trial, Tsubaki approached the rope and pulled one end while Mizuki pulled the other end, resulting in the first successful trial. Outcomes were variable after this trial. The change in frequency of complete successes in each session with the long rope is shown in Fig. 2. Complete successes increased as the number of sessions increased (Spearman r s = 0.60; P < 0.001).

Change in the frequency of complete successes across sessions in the long rope-length treatment
Table 1 shows the frequency of the glancing sequences during the learning phase. Mizuki showed more glancing than Tsubaki (Mann–Whitney z = 7.37; P < 0.001) in the long-rope condition. Tsubaki rarely showed glancing behavior in the long-rope trials, but began to do so in the second half of the medium-rope condition (Table 1). Mizuki glanced at her partner at least once on every session across all treatments (Table 1). In terms of the behavioral sequences associated with glancing, Mizuki pulled the rope after glancing at the partner who did not hold the rope less frequently with the medium, slightly short, and short ropes than with the long rope (Kruskal–Wallis H = 22.3; P < 0.01). Mizuki was first observed to wait in the 13th session with the long rope (Fig. 3). The frequency of waiting by Mizuki was significantly higher in the second half of the long-rope treatment than the first half (Mann–Whitney z = 3.00; P < 0.01).

Mizuki looking at her partner Tsubaki and waiting
The average occurrence of complete success during the long-, medium-, slightly short-, and short-rope treatments was 71, 82, 88, and 92%, respectively. Thus, the chimpanzees succeeded in most trials even in the short-rope treatment, which was identical to the initial test.
The percentage of trials in which both subjects started to approach the rope within 0.5 s after the experimenter moved aside was 121/300 (40%), 88/100 (88%), 94/100 (94%), and 99/100 (99%) in the long-, medium-, slightly short-, and short-rope treatments, respectively. Thus, latency to approach the rope decreased across conditions. Following a similar pattern, the percentage of trials in which both subjects approached the rope within 0.5 s of each other was 106/300 (35%), 74/100 (74%), 94/100 (83%), and 86/100 (86%) in the long-, medium-, slightly short-, and short-rope treatments, respectively.
Waiting room test
The chimpanzees were seldom successful in early sessions, but the incidence of success increased in later sessions. Mizuki showed waiting behavior from the 9th to 16th sessions, for a total of eight times (Table 2), but waiting did not result in success at the time. In these trials, Mizuki waited for 2–19 s (mean: 7.6 s), but Tsubaki did not approach the rope; eventually, Mizuki pulled only one end of the rope, resulting in a failed trial. Tsubaki began to show waiting behavior from the 17th session, and waited consistently thereafter (Table 2). In most of these trials, Mizuki approached the rope while Tsubaki was waiting, and the frequency of complete success increased accordingly (Spearman r s = 0.79, P < 0.01; Fig. 4). The percentage of trials in which both subjects approached the rope within 0.5 s of each other was 13/115 (11%). Thus, many of the successes occurred as a result of Tsubaki’s waiting behavior.

Change in the frequency of complete successes across sessions in the waiting room test in which the chimpanzees were released into the booth from a waiting room
Soliciting behavior
No interactions were observed between the two chimpanzees during any sessions of the initial test, learning phase, or waiting room test. In the long-, medium-, and slightly short-rope treatments during the learning phase, it was possible for one subject to succeed alone, i.e., it was not necessary to work with the partner. However, the partner was required for success during the initial test, short-rope treatment of the learning phase, and the waiting room test. In 106 failed trials in these tests, the subjects could have succeeded by interacting with the partner before pulling the rope (30 trials in the initial test, 5 in the short-rope treatment of the learning phase, and 71 in the waiting room test). However, no such behavior was observed. In addition, the two chimpanzees never showed eye-to-eye contact.
Test with a human partner
The chimpanzees showed soliciting behavior toward a human partner but not toward the conspecific partner. In the first session with the human partner, success was achieved in all five trials in which the partner adjusted to Mizuki, but all trials in which the partner approached the rope 2 s later than Mizuki resulted in failure because Mizuki ignored the partner when pulling the rope. In the second session the first four trials (trials 1, 3, 5, and 7) in which the partner adjusted to Mizuki were successful, whereas trials 2 and 6, in which the partner approached the rope 2 s later than Mizuki, resulted in failure. In the fourth trial, Mizuki waited for the partner without approaching the rope, resulting in success. In the eighth trial, when the partner did not approach the rope, Mizuki looked up at his face, whimpered, and took the partner’s hand (Fig. 5). Note that the partner was unaware that Mizuki looked at his face because he always looked forward to avoid cuing Mizuki. When Mizuki took the partner’s hand, both approached the rope and successfully completed the task. Soliciting by Mizuki, i.e., taking the partner’s hand when approaching the rope, occurred in all later trials except for trial 1 of the third session (21 trials in total), regardless of whether the partner approached the rope immediately or after a delay.

Mizuki looking up at the face of the human partner, whimpering, and taking the partner’s hand
In the fifth session, Mizuki was paired with Tsubaki for the first six trials. Mizuki never showed soliciting behavior to Tsubaki. Tsubaki did not pull the rope in trials 1, 2, 5, or 6. In trial 3, she pulled the rope before Mizuki, and failed. In the fourth trial, both pulled the rope simultaneously and dragged the blocks within reach, but Tsubaki did not take the food. In the subsequent three trials, Mizuki was again paired with the human partner. She took the partner’s hand before approaching the rope in all three trials. In the final three trials, Mizuki was again paired with Tsubaki, but she never showed soliciting behavior. Tsubaki did not approach the rope, and all of the trials ended in failure.
Discussion
The chimpanzees were never successful in the initial test, as they did not adjust their behavior to work with their partner. However, the frequency of success gradually increased across sessions. The emergence of waiting behavior was a factor in the rise in successful performances. Thus, the two chimpanzees learned to coordinate their own behavior with that of the partner, after trial and error.
Although the chimpanzees synchronized their pulling with that of the partner by checking and waiting for the partner, they seemed to use another tactic. In most of the later trials of the learning phase, both chimpanzees approached the rope immediately after the experimenter moved aside. Because the distance to the rope was the same for both chimpanzees, this synchronized start usually resulted in the synchronized arrival at each end of the rope. Thus, success could be achieved without checking the partner’s behavior; the chimpanzees used the behavior of the experimenter as a cue and established their own routine.
This explains why the frequency of success was low at the beginning of the waiting room test, in which no experimenter was present. However, the chimpanzees eventually learned to wait for the partner and synchronize timing of pulling the rope, again through establishing their own routine. In all successful trials, Tsubaki entered the room first, approached the same end of the rope, and waited for Mizuki, who approached the other end of the rope. They then pulled the ends of the rope simultaneously.
Mutual eye contact was never observed between the chimpanzees. Either one of the two individuals could coordinate her behavior to that of the partner, but they did not use mutual eye contact to do this. Chalmeau and colleagues also found that coordination was one-directional (Chalmeau 1994; Chalmeau et al. 1997a). However, the pairs that they tested were asymmetric: one member was either very young or was suspected to have mental retardation. In contrast, Mizuki and Tsubaki were similar in size and age, had no known physical or mental handicaps, and both of them became experienced in this task. However, they did not interact with each other to facilitate solution of the task.
The absence of interactive or soliciting behavior toward the conspecific partner is consistent with the results of Povinelli and O’Neill (2000), although the task that we presented was more difficult in terms of the necessity to synchronize the timing of pulling. The only behavior shown to the partner by the two chimpanzees was glancing and waiting. Tomasello et al. (2005) reviewed the experimental studies of cooperation in chimpanzees and concluded that almost no communication between partners was observed. These authors suggested that chimpanzees lack shared intentionality, which can be defined as collaborative interactions in which participants have a shared goal (shared commitment) and coordinated action roles to pursue that goal. The absence of interactions during this study may be ascribed to the absence of shared intentionality as suggested by Tomasello et al. (2005).
In contrast to the absence of soliciting behavior toward the conspecific partner, however, Mizuki spontaneously solicited help from the human partner. The dominance relationship with the partner may affect the likelihood of such behavior, but this does not account for the absence of soliciting behavior toward Tsubaki. Mizuki sometimes monopolized both food items when Tsubaki also approached the food. Therefore, it is unlikely that subordinate status caused Mizuki not to interact with her conspecific partner. Soliciting behavior by a gorilla was previously described by Gómez (1990). When a female infant lowland gorilla was placed in a room with humans, she approached a human, took his hand, and led him to the door, which was locked by a latch. The gorilla then placed the human’s hand on the latch and looked at his eyes. These behaviors were considered to be the gorilla communicating to the human that he should open the latch.
Such cases are often discussed in terms of enculturation, when raising an ape in a human cultural environment encourages it to engage in collaborative and communicative behaviors. This may result in more human-like communicative behaviors than if the ape was raised in conspecific society (Call and Tomasello 1996; Gómez 2004). Although the chimpanzees in Crawford’s (1937) study displayed soliciting behaviors toward conspecific partners, Povinelli and O’Neill (2000) point out that these behaviors were observed only after extensive shaping and postulated that solicitation may emerge through a process of conventionalization. However, our results indicate the need for more careful attention to such cases. This is the first clear demonstration that a chimpanzee raised by humans communicates differently with humans and conspecifics in the same situation. These behavioral differences do not support the idea that enculturation alters the general understanding of how to act with others and that gestures are used in a conventionalized way toward different partners. Rather, experience with others, probably through trial and error in various types of interaction, including play, may lead to an understanding of how specific individuals respond to one’s own behaviors. Thus, in the course of daily interactions, Mizuki may have learned that it was fruitless to show soliciting behavior to Tsubaki, whereas it worked with humans. However, our interpretation is tentative, because we have data from only one chimpanzee–chimpanzee pair and one chimpanzee–human pair.
The methods used here are easy to replicate, in contrast to Crawford’s (1937) procedure. Melis et al. (2006) used our method and reported that several chimpanzees spontaneously solved this type of task. The apparatus used by Melis et al. (2006) was slightly different from ours: the ends of the rope extending into the room were longer, meaning that the chimpanzees did not have to completely synchronize pulling. It is likely that coordinating timing with the partner was the difficult point for the chimpanzees in our study, which required a learning phase. Further studies, such as cross-species comparisons using the same methodology, will clarify the evolutionary roots of cooperation, which is fundamental to human social behavior.
References
Boesch C, Boesch H (1989) Hunting behavior of wild chimpanzees in the Tai National Park. Am J Phys Anthropol 78: 547–573
Call J, Tomasello M (1996) The effect of humans on the cognitive development of apes. In: Russon AE, Bard KA, Parker ST (eds) Reaching into thought: the minds of the great apes. Cambridge University Press, Cambridge, pp 371–403
Chalmeau R (1994) Do chimpanzees cooperate in a learning task? Primates 35:385–392
Chalmeau R, Gallo A (1996a) What chimpanzees (Pan troglodytes) learn in a cooperative task. Primates 37:39–47
Chalmeau R, Gallo A (1996b) Cooperation in primates: critical analysis of behavioral criteria. Behav Proc 35:101–111
Chalmeau R, Lardeux K, Brandibas P, Gallo A (1997a) Cooperative problem solving by orangutans (Pongo pygmaeus). Int J Primatol 18:23–32
Chalmeau R, Visalberghi E, Gallo A (1997b) Capuchin monkeys, Cebus apella, fail to understand a cooperation task. Anim Behav 54:1215–1225
Crawford MP (1937) The cooperative solving of problems by young chimpanzees. Comp Psychol Monogr 14:1–88
Garber P (1997) One for all and breeding for one: cooperation and competition as a tamarin reproductive strategy. Evol Anthropol 5:187–199
Gómez JC (1990) The emergence of intentional communication as a problem-solving strategy in the gorilla. In: Parker ST, Gibson KR (eds) “Language” and intelligence in monkeys and apes: comparative developmental perspectives. Cambridge University Press, New York, pp 333–355
Gómez JC (2004) Apes, monkeys, children, and growth of mind. Harvard University Press, Cambridge
Idani G, Hirata S (2006) Studies at the Great Ape Research Institute. In: Washburn DA (ed) Primate perspectives on behavior and cognition. American Psychological Association, Washington, pp 29–36
Melis AP, Hare B, Tomasello M (2006) Chimpanzees recruit the best collaborators. Science 311:1297–1300
Mendres KA, de Waal FBM (2000) Capuchins do cooperate: the advantage of an intuitive task. Anim Behav 60:523–529
Muroyama Y (1991) Mutual reciprocity of grooming in female Japanese macaques (Macaca fuscata). Behaviour 119:161–170
Povinelli DJ, O’Neill D (2000) Do chimpanzees use their gestures to instruct each other? In: Baron-Cohen S, Tager-Flusberg H, Cohen D (eds) Understanding other minds: perspectives from developmental cognitive neuroscience, 2nd edn. Oxford University Press, Oxford, pp 459–487
Tomasello M, Carpenter M, Call J, Behne T, Moll H (2005) Understanding and sharing intentions: the origins of cultural cognition. Behav Brain Sci 28:675–690
Acknowledgments
This study was financially supported by the Ministry of Education, Culture, Sports, Science and Technology, Japan (grant 1870266 to S. Hirata), the Sasagawa Scientific Research Grant from the Japan Science Society (grant 16–308 to S. Hirata), and the Core-to-Core Program HOPE by the Japan Society for the Promotion of Science. We thank S. Sekine, K. Sugama, and K. Kusunoki for assistance with data collection, and N. Morimura and G. Idani for support and suggestions. Thanks are also due to N. Sato, F. Kawashima, and T. Nanba for support in conducting the experiment and for caring for the chimpanzees. The chimpanzees were cared for according to the “Guide for the Care and Use of Laboratory Animals” of Hayashibara Biochemical Laboratories, Inc., and the guidelines laid down by the Primate Society of Japan.
Author information
Authors and Affiliations
Corresponding author
About this article
Cite this article
Hirata, S., Fuwa, K. Chimpanzees (Pan troglodytes) learn to act with other individuals in a cooperative task. Primates 48, 13–21 (2007). https://doi.org/10.1007/s10329-006-0022-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10329-006-0022-1