
Abstract
The effect of fruit availability on chimpanzee party size was investigated in the montane forest of Kahuzi. Seasonal variation in both fruit availability and party size was examined. Fruit abundance per se does not affect chimpanzee party size. However, seasonality and distribution patterns of fruits are both determinant ecological factors that control the size of chimpanzee parties at Kahuzi. There was no correlation between fruit abundance and the spatial distribution of fruits. When fruits were clumped and available in large amounts for a long period, chimpanzee party size increased, or otherwise decreased when fruits were highly available for only a limited period. Tree species that produced only a small amount of ripe fruit throughout the year did not affect the foraging party size of chimpanzees. Temporal and spatial variability in fruit abundance seems to constrain grouping patterns of chimpanzees at Kahuzi more so than in other chimpanzee habitats previously described.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Earlier studies on the social organization of chimpanzees revealed flexibility both in size and composition of party (individuals gathering together), known as a fission–fusion social system (Nishida 1968; Nishida and Kawanaka 1972). More recently, the focal point of numerous empirical researches has aimed to better understand the factors that influence this social structure. Ecological factors, in particular food availability, have been frequently tested in the field to assess how this affects chimpanzee foraging party size.
The ecological constraint model predicts that the size of primate social groups should have an upper limit that is determined by the abundance and distribution of food (Chapman and Chapman 2000).
Findings from several field studies have revealed that chimpanzees tend to increase their foraging party size when fruits are abundant or have a clumped distribution (Goodall 1986; Chapman et al. 1995; Boesch 1996; Doran 1997; Matsumoto-Oda et al. 1998; Wrangham et al. 1992). Conversely, fruit abundance has not been found to affect chimpanzee party size at Kalinzu (Hashimoto et al. 2001). There was no relationship between these two factors at Kibale when multiple sources of important foods were available (Isabirye-Basuta 1988). Comparison between chimpanzee party size and fruit abundance yielded only a weak relationship at Budongo forest (Newton-Fisher et al. 2000). Stanford et al. (1994) reported the presence of estrous females as the main factor predicting larger chimpanzee parties and promoting the males’ decision to hunt at Gombe. They found that chimpanzee parties were largest during the dry season and early wet season when their food supply was restricted, suggesting that food availability was not the major factor setting the upper limit of chimpanzee party size at Gombe. Therefore, effects of food supply on chimpanzee grouping patterns differ greatly across habitats. Furthermore, only a few studies have measured fruit abundance quantitatively (Newton-Fisher et al. 2000; Wrangham 2000; Anderson et al. 2002; Mitani et al. 2002), among them only the study by Newton-Fisher et al. (2000) analyzed the effect of particular fruit tree species’ phenology on chimpanzee group size. Newton-Fisher et al. (2000) suggested that both abundance and dispersal should be considered when investigating the impact of food supply on grouping patterns of chimpanzees. There are a number of potential variables of food availability (such as abundance, distribution, seasonality, degree of preference, etc.) that may influence party size in chimpanzees.
This paper aims to investigate how chimpanzee group size varies in relation to temporal and spatial changes in the availability of particular food fruit species. I contrast the effect (on chimpanzee party size) of tree species that produce fruits for only a short period of the year versus those that produce fruits year-round. Kahuzi chimpanzees have been found up to 2,600 m above sea level (a.s.l.), the highest ever reported altitude for Pan troglodytes schweinfurthii distribution. A previous botanical survey (Yamagiwa et al. 1993, 1996) indicated that in comparison to lowland and mid-altitude forests, the Kahuzi montane forest is characterized by lower fruit diversity and productivity. Therefore, unlike their conspecifics in other forested habitats where fruits are abundant, Kahuzi chimpanzees may face a more limited range of fruits, and this may affect their grouping patterns.
Methods
Study site
The study area covers approximately 30 km2 of montane forest and is located along the eastern border of Kahuzi-Biega National Park, at an altitude ranging between 2,050 and 2,350 m a.s.l
The vegetation is predominantly montane forest characterized by a low canopy, which includes bamboo (Arundinaria alpina) forest, primary forest, secondary forest, and Cyperus latifolius swamps. Casimir (1975), Goodall (1977), and Yumoto et al. (1994) described in detail the compositions of the different types of vegetation. Dominant species of trees include Podocarpus usambarensis (Podocarpaceae), Symphonia globulifera (Guttiferae), and Carapa grandiflora (Meliaceae) in the primary forest; Hagenia abyssinica (Rosaceae), Myrianthus holstii (Moraceae), and Vernonia spp. (Compositae) in the secondary forest; Hypericum revolutum (Guttiferae) and Rapanea melanophloeos (Myrsinaceae) in the Cyperus swamp; and S. globulifera (Guttiferae) and Syzygium parvifolium (Myrtaceae) in and around swamp areas. Herbs, vines, and ferns (Urera hypselodendron, Basella alba, Lactuca sp., Pteridium aquilinum, etc.) constitute the dense terrestrial vegetation of the secondary forest (Yumoto et al. 1994).
The climate is characterized by a clear dry season (June–August) and a distinct long rainy season, which may be divided into two wet periods according to the rainfall pattern: the early wet season from September to December and the late wet season from January to May. A mean annual rainfall of 1,619 mm/year has been recorded for a period of 14 years (1982–1995). The monthly mean temperature was 19.7°C and was constant over those 14 years (Basabose and Yamagiwa 2002).
Party size sampling
Counts of party size were performed on a unit group of chimpanzees showing semi-habituation that numbered 22 individuals comprising 8 adult and adolescent males (estimated to be more than 10 years old), 6 adult and adolescent females (estimated to be more than 10 years old), and 8 immature individuals including 2 infants (estimated to be 0–4 years old). The observations took place opportunistically from August 1994 to October 1996.
Chimpanzees were located by following their fresh trails in the forest, by localizing calls and searching where calls were heard, and by visiting fruiting trees known to be frequently visited by chimpanzees.
It was often difficult to count the chimpanzees within a group when animals were found dispersed throughout a dense habitat where visibility was limited. Counts were possible most of the time when the chimpanzees were up on trees feeding or resting. As only semi-habituated animals were observed, I kept a distance (20–30 m from which they often tolerated my presence) to count all the individuals within a given group. I then counted all the individuals in the party, endeavoring to count all the subjects remaining on the ground nearby. Parties analyzed in this study were mainly those located during foraging episodes on fruiting trees (n=71 parties), for which reliable individual counts were possible. They represent only about one third of all the parties encountered. On average, reliable chimpanzee counts were achieved for only six foraging parties (range: 0–15) per month. I excluded data for some months (February, May, and October 1995 and February 1996) during which reliable counts could not be achieved.
In a party, I counted all the individuals including adults, immatures, and infants. No fission–fusion events of parties were witnessed during the observations. The sex and age class of all individuals were identified to the extent possible. Due to incomplete habituation of the study group, and the dense vegetation that limits visibility on the ground layer within the forest, only foraging episodes offered a good opportunity to quantify chimpanzee parties. Therefore, only foraging parties were analyzed. The size of parties varied significantly between months (ANOVA, F=4.172; df=70; P<0.0002).
Anderson et al. (2002) reported that foraging parties are expected to be larger than traveling parties. Although there is no detailed data on the comparison between sizes of foraging and traveling parties of Kahuzi chimpanzees, I assumed from previous study group observations that there is no significant difference between both party types.
The study group has been regularly monitored since 1991 when the habituation program without artificial provisioning started. To assess whether the tolerance level of chimpanzees changed with group size, I recorded the duration that chimpanzee parties tolerated the presence of observers (generally three persons) from a distance of 20–30 m. This was done after counting all the chimpanzees present in a party encountered in a hiding site. Tolerance to observers was measured for 57 foraging parties for a total of 848 min. Lone individuals and mother–infant parties were generally shy and fled within 1 min after they noticed the presence of observers. I started to record the time when there were signs that the chimpanzees noticed our presence. I stopped recording the time when they left and when the departure was motivated by the presence of observers (this generally occurred when chimpanzees stopped eating, glanced toward observers for a brief moment, then fled one by one).
Fruit availability sampling
In order to estimate fruit food abundance in the study area, a vegetation survey was conducted using a belt transect 5,000 m long and 20 m wide providing a total area of 10 ha. The transect ran for 3,170 m (6.34 ha) in secondary forest, 1,212 m (2.42 ha) in primary forest and 618 m (1.24 ha) in C. latifolius dominated swamp. To quantify the distribution of fruit, this transect was divided into 20 contiguous sections, each 250×20 m (0.5 ha). I counted the number of stems and measured the basal area (m2) of all trees (DBH≥10 cm) present in all 20 sections along this transect. The presence of fruit was monitored twice a month from late August 1994 to early July 1996. A monthly fruit abundance index (Fm) of all tree species, the fruits of which are eaten by chimpanzees, was then calculated using the following formula:
in which P km denotes the proportion of fruiting trees for species k in month m, and B k denotes the total basal area per hectare for species k. Basal area [(1/2 DBH)2× π] is used as a good estimation of fruit abundance in tropical forests (Chapman et al. 1992).
Monthly distribution of fruit was quantified by calculating the degree of clumping as the ratio of the variance to mean: Coefficient of dispersion (CD, Zar 1999). The CD was calculated using data on fruit abundance in different sections of the transect, indexed as the sum of the DBHs of trees providing fruit food (Malenky et al. 1993; Chapman et al. 1995). In this study, I use the term “fruit abundance” to indicate the amount of fruit produced at a given time; whereas the term “fruit availability” indicates both fruit amount and distribution (in clumped or uniform patterns) within the habitat.
Fruit diet sampling
Because of poor visibility in the forest and the low level of habituation of the chimpanzee study group, I used an indirect method (fecal analysis) to describe the fruit diet of chimpanzees at Kahuzi. Fecal analysis as a method to describe the diet of the ape has a number of drawbacks, but is sometimes the most feasible way of studying the diet of gorillas and chimpanzees living in dense vegetation of tropical forests where visibility is limited, particularly at ground level. While fecal analysis alone cannot give a complete picture of diet, it is a valuable tool for studying poorly habituated ape populations. In particular, fruit remains may be easily identified in ape dung and therefore the method, although not quantitatively precise, can report what fruits are eaten and at what frequency.
Fresh feces (up to 1 day old) of chimpanzees were collected. Each dung sample was sluiced in 1-mm mesh sieves and dried in sunlight. For each dried fecal sample, the seeds were separated from other materials (such as green leaf fragments, fibers, animal items, soil, etc.). For fruit species with large seeds (>2 mm), I counted the exact number of seeds in the dung. Small seeds were classified as rare, common, or abundant with respect to the total mass of the fecal sample. I identified seeds to the species level in comparison with the seed samples identified by Yumoto et al. (1994) at the National Botanical garden in Belgium and those kept at our herbarium.
Data analysis
I conducted a series of regression analyses to determine which factor from among fruit abundance, fruit distribution, or fruit consumption most strongly influenced party size of chimpanzees. I analyzed the relationship between chimpanzee party sizes and fruit availability indices (abundance and distribution pattern) of different important fruit species in the chimpanzee’s diet. For fruit availability data, I used only fruits that are most preferred by chimpanzees at Kahuzi and for which trees were represented by at least ten individuals in the transect. Those species included 158 trees of B. bridelifolia, 64 trees of Allophyllus sp., 36 trees of Maesa lanceolata, 34 trees of S. parvifolium, 17 trees of Myrianthus holstii, and 10 trees of Ficus spp.
Ekebergia capensis, although among the preferred fruit species of Kahuzi chimpanzees and eaten more than Allophyllus sp. and M. lanceolata fruits (Basabose 2002), was not chosen. Trees of E. capensis were very rare in the transect (four individuals). I also regressed party size to the proportion of fruit consumption by chimpanzees (assessed by fecal analysis) to find out whether or not there was a relationship between these two factors.
Results
Party size and composition
Over the 23 months of observation, chimpanzee party size varied substantially (mean 4.43 individuals per party; range=1–13; SD=±2.50; n=71 parties). Analysis of 848 min of observations spent with foraging parties showed a significant positive correlation (r=0.38; P<0.005; n=57) between party size and the duration of observation, suggesting that chimpanzees tended to tolerate the presence of human observers best when in large parties.
Because of poor visibility, it was often difficult to distinguish the sex of immature and adolescent individuals in a party. Taking only the adults, parties contained slightly more females (mean=1.625; range: 0–6) than males (mean=1.291; range: 0–5) on average, but the difference was not statistically significant (Mann–Whitney U-test; U′=1,875.5; P=0.209, NS). That is, although males tended to range alone in solitary (14.08%) or all-male parties (5.63%), they often joined mixed parties including individuals of both sexes and all age classes. All-female parties were not observed throughout the study, while observations of consort pairs (including an adult male and an estrous female) occurred six times (8.45%).
Party size and fruit availability
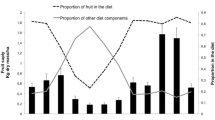
Figure 1 presents the monthly variation in fruit abundance and chimpanzee party size in the Kahuzi montane forest. Despite the variations observed in fruiting patterns among tree species, overall fruit abundance was negatively correlated with rainfall (r=−0.581, P<0.003, df=22). There was a statistically consistent seasonal difference in fruit availability. Dry months generally corresponded with periods of high fruit abundance (F 2,42=7.79, P<0.002, Fig. 2a). The fruit distribution patterns also varied seasonally. However, there was no relationship between fruit abundance and fruit distribution (P=0.285, NS). In contrast to fruit abundance, fruits were more clumped in the wet months than in dry ones (F 2,42=6.05, P<0.005), though no statistically significant difference was found between the dry season and early wet seasons (Fig. 2b). The seasonal difference observed between fruit abundance and fruit distribution patterns may be explained by the different phenological patterns of each fruit species (Fig. 1). Considering the relatively longer wet period at Kahuzi (9 months/year), this may have led to an overall clumped fruit distribution during the wet months.

Monthly changes in party size and fruit abundance for the six common woody species, the fruits of which are eaten by Kahuzi chimpanzees. The horizontal line crossing the histograms is the overall monthly fruiting index (Fm) mean. (Each month was divided into two periods: first and last 15 days. The number of trees the fruits of which were monitored during the study period are 10 Ficus spp., 36 Maesa lanceolata, 17 Myrianthus holstii, 34 Syzygium parvifolium, 158 Bridelia bridelifolia, and 64 Allophyllus sp.). Fm indicates the fruit abundance index. Phenological data were collected from late August 1994 to early July 1996, whereas party size data came from early August 1994 to late October 1996

Seasonal change in a fruit abundance, b fruit distribution, and c party size. The monthly fruit availability index (Fm) and coefficient of dispersion (CD) were calculated for six common fruit species eaten by chimpanzees: Ficus spp., Myrianthus holstii, B. bridelifolia, Maesa lanceolata, Allophyllus sp., and S. parvifolium
There was a seasonal difference in party size, with larger parties in the late wet season than in any other period of the year (Fig. 2c).
Fruit availability and fruit consumption effects on chimpanzee party size
An overall Fm median of 2 was found for all six fruit species. The median of Fm calculated for 24 months was used as a cut-off criteria for fruit abundance. Fm≥2 was considered as a small amount of fruit, whereas Fm<2 was considered as a small amount of fruit. Species for which a large amount of fruit was available year-round were those for which Fm was ≥2 for more than half of the months during the study period. Those species were Ficus spp. (17 months with sometimes over 7 consecutive months of a large amount of fruit) and M. lanceolata (19 months with up to 9 consecutive months of a large amount of fruit).
Syzygium parvifolium produced a large amount of fruit for only 10 months, with a gap of several months of a small amount of fruit. This species was considered as producing a large amount of fruit for a limited period (only 2–4 consecutive months). Other fruit species (M. holstii, B. bridelifolia and Allophylus sp.) produced a small amount of fruit throughout the study period.
The potential influence of fruit availability on chimpanzee grouping patterns indicated that party size was positively affected by overall fruit distribution (CD) of the six fruit species commonly eaten by chimpanzees at Kahuzi (r=0.38, P<0.05, df=28). Conversely, fruit abundance (Fm) showed no significant relationship with chimpanzee party size (Fig. 3). Separate analysis performed on each fruit species showed diverse effects of Fm on chimpanzee grouping patterns. The party size was positively correlated with Fm of species producing a large amount of fruit year-round (e.g., M. lanceolata: r=0.44, df=26, P<0.05). Party size was negatively related to the Fm of S. parvifolium r=−0.41, df=27, P<0.05), for which relatively large amounts of fruit were available only during a limited period of the year. Fruits of both M. lanceolata and S. parvifolium were dispersed in a clumped pattern during the study period (Fig. 4). However, though available in relatively large amount and distributed in a clumped pattern over the study period, fruit of Ficus spp. did not affect chimpanzee party size.

Relationship between party size and a fruit abundance index, and b coefficient of dispersion of six common fruit species eaten by chimpanzees

Mean coefficient of the distribution of six common fruit species eaten by chimpanzees at Kahuzi
Species that produced only a small amount of ripe fruit year-round (M. holstii, Allophyllus sp. and B. bridelifolia) and throughout the study period did not affect the foraging party size of chimpanzees. Fruits of all three species were evenly distributed with a mean CD<1 (Fig. 4).
Over 26 months from August 1994 to September 1996, a total of 3,377 chimpanzee fecal samples were collected, averaging 130 samples per month (range 28–427). Among the six most preferred fruit species, consumption of M. holstii, M. lanceolata, S. parvifolium, and Allophylus sp. varied seasonally, while no seasonal effect was found in the fruit consumption of Ficus spp. and B. bridelifolia (Table 1). Fruit consumption was not related to fruit availability (abundance and distribution) throughout the study (Spearman correlation tests, P>0.05). Moreover, fruit consumption was not related to the chimpanzee party size (Spearman correlation tests, P>0.05). These results suggest that Kahuzi chimpanzees may not adjust foraging party size to either fruit abundance or to the degree of fruit consumption, but to the distribution of preferred fruits.
Discussion
Comparison of party size between habitats is difficult due to the different methodologies used by observers. Definitions of party membership also vary between studies. Some studies have considered a party as all the individuals present therein (e.g., Tutin et al. 1983; Boesch 1996); whereas others exclude infants from the count (e.g., Chapman et al. 1994; Doran 1997). In this study, a party means all individuals present and parties reported were mostly foraging parties. Kahuzi chimpanzees tended to forage in parties containing a mean number of 4.43 individuals. Even when excluding the two infants of the community from the count (mean=4.10), the two means were not significantly different. The size of parties observed during this study ranged from lone individuals (mostly males) up to 13 individuals with more parties of 4 and 5 individuals (excluding lone individuals). The mean party size (4.43) was about 20% of the community size at Kahuzi, which was proportionally similar to those reported for other small communities at Bossou (Sakura 1994), Mt Assirik (Tutin et al. 1983) and Mahale (Nishida 1968). The difference emerged when compared to the larger communities of Tai, Gombe, Budongo, and Kibale (Table 2). There was a positive correlation between the mean party size and the community size (Spearman correlation test; r=0.8833; n=9; P<0.002). The small size of the study community (22 individuals) reflected a low density (0.13 individual/km2) in the Kahuzi montane forest (Yamagiwa et al. 1992). It has been suggested that this low density among forest dwelling chimpanzee populations may be due to the low production and diversity of fruit trees (Yamagiwa et al. 1996).
Overall, the relationship between fruit abundance and chimpanzee party size appeared to be weak at Kahuzi, suggesting that factors other than fruit abundance alone are responsible for the size of chimpanzee parties. Previous studies reported the presence of estrous females (Goodall 1986; Stanford et al. 1994; Boesch 1996; Matsumoto-Oda 1999; Hashimoto et al. 2001) and monkey-hunting season (Stanford et al. 1994; Boesch 1996) as factors that increase the size of chimpanzee parties. Anderson et al. (2002) suggested that when estrous females were present, Tai chimpanzee parties tended to remain large regardless of fruit availability. Given that estrous females and males gathering or dispersing may be driven by fruit abundance, both factors (fruit availability and presence of estrous females) might be considered when evaluating their effect on chimpanzee party size, since they may affect each other. Food availability and the number of estrous females were both found positively correlated with monthly party size of the Ngongo chimpanzee in the Kibale National Park (Mitani et al. 2002).
At Kahuzi, the constraints on visibility due to dense vegetation limited the investigation of estrous females on chimpanzee party size. Presence of estrous females was commonly seen in bisexual, mixed, and consort parties throughout the study. However, I did not observe more than two estrous females in a party. Their potential effect on the size of chimpanzee parties has not yet been studied at Kahuzi.
Other studies (Stanford et al. 1994; Boesch 1996) suggested that monkey-hunting is a factor that increases the size of chimpanzee parties. Observations of carcasses and fecal specimens indicated that predation on mammals occurred during periods of high fruit diversity at Kahuzi (Basabose and Yamagiwa 1997). Foraging parties presented in this study may be different in size and composition from hunting parties. Party sizes in chimpanzees vary with different behavioral activities (Boesch 1996). It is still unknown whether hunting at Kahuzi occurs when chimpanzee parties are large. Moreover, the absence the of red colobus monkey at Kahuzi, the most frequent prey at Gombe (Stanford et al. 1994), Mahale (Uehara et al. 1992), and Tai (Boesch and Boesch 1989), may result in different chimpanzee hunting strategies between Kahuzi and these habitats. Further observations are needed to elucidate the relationship between hunting, fruit availability and prey vulnerability at Kahuzi.
Fruit diet did not affect Kahuzi chimpanzee party size. A previous study (Basabose 2002) reported that chimpanzees did not eat fruits according to their abundance at Kahuzi. They actively seek a particular fruit species rather than simply consume it opportunistically. Because the quantity of fruits actually eaten by chimpanzees may not be exactly known from fecal analysis, performing a correlation test between fruit availability and fruit consumption resulting from indirect observation (fecal analysis) may produce biases on the actual relationship between both factors. While this comparison may not report the precise situation, it serves however to draw a tendency which may be verified by further direct observations. Further observations recording feeding time actually spent by chimpanzee parties in individual fruiting tree species are needed to assess the degree of influence of fruit consumption on chimpanzee party size.
Given the low fruit productivity that characterized Kahuzi montane forest, chimpanzees may be expected to increase party size when fruits are abundant. In contrast, the study shows that party size is not affected by fruit abundance, whether they are preferred food items or not. The absence of the expected relationship may in part be due to the difference in the phenological pattern of major chimpanzee food fruit species at Kahuzi.
Kahuzi chimpanzee party size was larger in the wet months, when fruits were less abundant. In comparison with other habitats, where party size was reported to vary seasonally, the inverse pattern of party size versus fruit abundance was not consistent with both Tai and Mahale (Doran 1997; Matsumoto-Oda 2002). Doran (1997) reported smaller parties in July (dry season), when preferred fleshy fruits were less available at Tai, suggesting a positive correlation between both factors. Accordingly, at Mahale, where chimpanzee party size was positively correlated with fruit abundance (Itoh et al. 1999; Matsumoto-Oda 2002), party size was significantly larger in the early dry season, when fruits were more abundant. Why were the sizes of chimpanzee parties at Kahuzi larger in the wet months when fruit abundance was the lowest? The diversity and abundance of fruits in Kahuzi montane forest are relatively smaller than those in other habitats. Kahuzi chimpanzees tended to increase folivorous diet such as pith, leaves, and terrestrial herbaceous vegetation (THV) during fruit scarcity (Basabose 2002). Frugivory in Kahuzi chimpanzees may increase intragroup competition over fruits and limit their party size during the dry season, while folivorous diet may mitigate their competition, allowing them to form larger parties in the late wet season, when fruit abundance was the lowest during the study period at Kahuzi.
In an attempt to explain the effect of fruit on Kahuzi chimpanzee grouping patterns, this study suggests that foraging party size is affected by the distribution pattern of fruits. Chimpanzees increase foraging party size when fruits have a clumped dispersion in the habitat. Furthermore, intra-group competition may arise during periods of high availability of depleting fruits (available for only a limited period), which may therefore constrain the size of the foraging party. The distribution of S. parvifolium fruit was clumped, but ripe fruit was abundant for only a limited period (2–4 consecutive months). Chimpanzee party size was inversely correlated to fruit abundance of S. parvifolium, suggesting that the presence of such fleeting food may negatively affect the grouping pattern of chimpanzees at Kahuzi. It has been suggested that when animals rely on food items found in depleting patches, group size will be constrained by the size, density, and distribution of food patches (Chapman et al. 1995). Competition on ephemeral, patchy food items may force chimpanzees into small parties (Wrangham 1977, 1986). At Kahuzi, it seems likely that depleting fruits (e.g., S. parvifolium) may limit the party size of chimpanzees by increasing competition for fruits within foraging groups, leading to the formation of small groups. When large amount of fruits are available for a relatively longer period of 7–11 consecutive months and are distributed in a clustered pattern (e.g., M. lanceolata), Kahuzi chimpanzees may increase the size of foraging parties. Newton-Fisher et al. (2000), suggested that low food abundance may force Sonso chimpanzees into parties of one or two individuals, but as food increases, the constraint on party size is relaxed, until food supply becomes unimportant in determining the sizes of parties.
Species producing a small amount of fruit throughout the study period (M. holstii, Allophyllus sp. and B. bridelifolia) did not affect the grouping pattern of chimpanzees at Kahuzi.
However, fruits of Ficus spp. were produced in relatively large amounts throughout the study period and distributed in a clumped pattern, but did not affect party size. Fig fruit is available year-round at Kahuzi, though the amount varies between months. Hashimoto et al. (2001) reported that the constant availability of Musanga leo-errerae and Ficus spp. in the Kalinzu forest led to a general decrease in effect of fruit on chimpanzee party size. Fig fruit is staple food for Kahuzi chimpanzees. They eat fig fruit throughout the year, regardless of the period of scarcity or abundance of other pulp fruits (Basabose 2002). The absence of seasonality in both availability and consumption of fig fruit at Kahuzi may be partially responsible for the lack of or small influence of fig fruit on chimpanzee party size.
This study suggests that food abundance per se does not affect Kahuzi chimpanzee party size, but that seasonal variation observed in the size of parties is mostly driven by the distribution pattern of preferred ripe fruits. Fruit species, the trees of which produce a large amount of ripe fruit in a clumped distribution, with the exception of Ficus spp., are expected to affect chimpanzee party size at Kahuzi. When a large amount of these fruits were available for a relatively long period, party size increased or otherwise decreased when they were available for only a limited period. This grouping strategy of chimpanzee foraging parties may reduce intra-group competition over fruit in the montane forest of Kahuzi, where fruit is considered a limited food resource. The effect of temporal and spatial variability of fruit, which seems to constrain grouping patterns of chimpanzees at Kahuzi more than in other habitats, should be described by a long-term study.
References
Anderson DP, Nordheim EV, Boesch C, Moermond TC (2002) Factors influencing fission–fusion grouping in chimpanzees in the Tai National Park, Côte d’Ivoire. In: Boesch C, Hohmann G, Marchant LF (eds) Behavioural diversity in chimpanzees and bonobos. Cambridge University Press, Cambridge, pp 90–1001
Basabose AK (2002) Diet composition of chimpanzees inhabiting the montane forest of Kahuzi, Democratic Republic of Congo. Am J Primatol 58:1–21
Basabose K, Yamagiwa J (1997) Predation on mammals by chimpanzees in the montane forest of Kahuzi, Zaire. Primates 38:45–55
Basabose AK, Yamagiwa J (2002) Factors affecting nesting site choice in chimpanzee at Tshibati, Kahuzi-Biega National Park: influence of sympatric gorillas. Int J Primatol 23:263–282
Boesch C (1996) Social grouping in Tai chimpanzees. In: McGrew WC, Marchant L, Nishida T (eds) Great ape society. Cambridge University Press, Cambridge, pp 101–113
Boesch C, Boesch H (1989) Hunting behaviour of wild chimpanzees in the Tai National Park. Am J Phys Anthropol 78:547–573
Boesch C, Boesch-A H (2000) The chimpanzee of the Tai Forest: Behavioural ecology and evolution. Oxford University Press, Oxford
Casimir MJ (1975) Feeding ecology and nutrition of an eastern gorilla group in the Mt. Kahuzi region (République du Zaíre). Folia Primatol 24:81–136
Chapman CA, Chapman LJ (2000) Determinants of group size in primates: the importance of travel costs. In: Boinski S, Garber PA (eds) On the move: how and why animals travel in group. University of Chicago Press, Chicago, pp 24–42
Chapman CA, Chapman LJ, Wrangham RW, Hunt K, Gebo D, Gardner L (1992) Estimators of fruit abundance of tropical trees. Biotropica 24:527–531
Chapman CA, White FJ, Wrangham, RW (1994) Party size in chimpanzees and bonobos. In: Wrangham RW, McGrew WC, de Waal FBM, Heltne PG (eds) Chimpanzee culture. Harvard University Press, Cambridge, pp 41–57
Chapman CA, Wrangham RW, Chapman LJ (1995) Ecological constraints on group size: an analysis of spider monkey and chimpanzee subgroups. Behav Ecol Sociobiol 36:59–70
Clark AP, Wrangham RW (1994) Chimpanzee arrival pant hoots: Do they signify food or status? Int J Primatol 15:185–205
Doran D (1997) Influence of seasonality on activity patterns, feeding behavior, ranging and grouping patterns in Taíchimpanzees. Int J Primatol 18:183–206
Goodall AJ (1977) Feeding and ranging behavior of a mountain gorilla group (Gorilla gorilla beringei) in the Tshibinda–Kahuzi region (Zaire). In: Clutton-Brock TH (ed) Primate ecology: studies of feeding and ranging behaviour in lemurs, monkeys and apes. Academic Press, London, pp 450–479
Goodall J (1986) The chimpanzees of Gombe: patterns of behavior. Harvard University Press, Cambridge, MA
Hashimoto C, Furuichi T, Tashiro Y (2001) What factors affect the size of chimpanzee parties in the Kalinzu forest, Uganda? Examination of fruit abundance and number of estrous females. Int J Primatol 22:947–959
Isabirye-Basuta G (1988) Food competition among individuals in a free-ranging chimpanzee community in Kibale forest, Uganda. Behaviour 105:135–147
Itoh N, Nakamura M, Nishida T (1999) Fission–fusion pattern of M group chimpanzees of Mahale: fruit food availability and party size (Abstract). Primate Res 15:436
Malenky RK, Wrangham RW, Chapman CA, Vineberg EO (1993) Measuring chimpanzee food abundance. Tropics 2:231–244
Matsumoto-Oda A (1999) Mahale chimpanzees: grouping patterns and cycling females. Am J Primatol 47:197–207
Matsumoto-Oda A (2002) Behavioral seasonality in Mahale chimpanzees. Primates 43:103–117
Matsumoto-Oda A, Hosaka K, Huffman MA, Kawanaka K (1998) Factors affecting party size in chimpanzees of the Mahale Mountains. Int J Primatol 19:999–1011
Mitani JC, Watts DP, Lwanga JS (2002) Ecological and social correlates of chimpanzee party size and composition. In: Boesch C, Hohmann G, Marchant LF (eds) Behavioural diversity in chimpanzees and bonobos. Cambridge University Press, Cambridge, pp 102–111
Newton-Fisher NE, Reynolds V, Plumptre AJ (2000) Food supply and chimpanzee (Pan troglodytes schweinfurthii) party size in the Budongo forest reserve, Uganda. Int J Primatol 21:613–628
Nishida T (1968) The social group of wild chimpanzees in the Mahali Mountains. Primates 9:167–224
Nishida T, Kawanaka K (1972) Inter-unit-group relationships among wild chimpanzees of the Mahali Mountains. Kyoto University. Afr Stud 7:131–169
Sakura O (1994) Factors affecting party size and composition of chimpanzees (Pan troglodytes verus) at Bossou, Guinea. Int J Primatol 15:167–183
Stanford C, Wallis J, Mpongo E, Goodall J (1994) Hunting decisions in wild chimpanzees. Behaviour 131:1–18
Tutin CEG, McGrew WC, Baldwin PJ (1983) Social organization of savanna-dwelling chimpanzees, Pan troglodytes verus, at Mt. Assirik, Senegal. Primates 24:154–173
Uehara S, Nishida T, Hamai M, Hasegawa T, Hayaki H, Huffman MA, Kawanaka K, Kobayashi S, Mitani JC, Takahata Y, Takasaki H, Tsukahara T (1992) Characteristics of predation by the chimpanzees in the Mahale Mountains National Park, Tanzania. In: Nishida T, McGrew WC, Marler P, Pickford M, de Waal FBM (eds) Tropics in primatology, vol 1. Human origins. University of Tokyo Press, Tokyo, pp 143–158
Wrangham RW (1977) Feeding behavior of chimpanzees in Gombe National Park, Tanzania. In: Clutton-Brock TH (ed) Primate ecology: studies of feeding and ranging behaviour in lemurs, monkeys and apes. Academic Press, London, pp 503–538
Wrangham RW (1986) Ecology and social relationships in two species of chimpanzees. In: Rubenstein DI, Wrangham RW (eds) Ecological aspects of social evolution: birds and mammals. Princeton University Press, Princeton, pp 352–378
Wrangham R (2000) Why are male chimpanzees more gregarious than mothers? A scramble competition hypothesis. In: Kappeler P (ed) Primate males. Causes and consequences of variation in group composition. Cambridge University Press, Cambridge
Wrangham RW, Clark AP, Isabirye-Basuta G (1992) Female social relationships and social organization of Kibale chimpanzees. In: Nishida T, McGrew WC, Marler P, Pickford M, de Waal FBM (eds) Topics in primatology, vol 1. Human origins. University of Tokyo Press, Tokyo, pp 81–98
Yamagiwa J, Mwanza N, Spangenberg A, Maruhashi T, Yumoto T, Fischer A, Steinhauer BB, Refisch J (1992) Population density and ranging pattern of chimpanzees in Kahuzi-Biega National Park, Zaire: a comparison with a sympatric population of gorillas. Afr Study Monogr 13:217–230
Yamagiwa J, Mwanza N, Spangenberg A, Maruhashi T, Yumoto T, Fischer A, Steinhauer BB (1993) A census of the eastern lowland gorillas Gorilla gorilla graueri in Kahuzi-Biega National Park with reference to mountain gorillas G.g. beringei in the Virunga region, Zaire. Biol Conserv 64:83–89
Yamagiwa J, Kaleme K, Mwanga M, Basabose K (1996) Food density and ranging patterns of gorillas and chimpanzees in the Kahuzi-Biega National Park, Zaire. Tropics 6:65–77
Yumoto T, Yamagiwa J, Mwanza N, Maruhashi T (1994) List of plant species identified in Kahuzi-Biega National Park, Zaire. Tropics 3:295–308
Zar JH (1999) Biostatistical analysis, 4th edn. Prentice-Hall, Upper Saddle River, New Jersey
Acknowledgements
This study was financed by the International Scientific Research Program of Japan (No. 08041146 to Prof. J. Yamagiwa) in cooperation with CRSN (Centre de Recherche en Sciences Naturelles) in the Democratic Republic of Congo. I thank Prof. J. Yamagiwa for his supervision during my field research at Kahuzi. I would like to thank ICCN (Institut Congolais pour la Conservation de la Nature) for granting permission to carry out this study in its National Park. Thanks are also due to Prof. S. Bashwira, Dr. M. Nshombo, Dr. B. Baluku, Mr. B. Kasereka, Mr. M.O. Mankoto, and Ms. S. Mbake for their administrative help. I am grateful to Mr. K. Kaleme, Mr. M. Bitsibu, Mr. S. Kamungu and all of the guides and guards of Kahuzi-Biega National Park for their technical help. Thanks are also extended to Prof. M.A. Huffman and two anonymous reviewers for helpful comments on earlier versions of this manuscript.
Author information
Authors and Affiliations
Corresponding author
About this article
Cite this article
Basabose, A.K. Fruit availability and chimpanzee party size at Kahuzi montane forest, Democratic Republic of Congo. Primates 45, 211–219 (2004). https://doi.org/10.1007/s10329-004-0087-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10329-004-0087-7