
Abstract
The growing populations of sika deer (Cervus nippon yesoensis) in Hokkaido, Japan, are having increasingly negative impacts on forests. Although the percentage or frequency of browsed twigs and seedlings is a useful index of the impact of deer on forests, differences in the browse preferences of deer among tree species should also be considered. We evaluated the woody browse preferences of sika deer and examined a method for evaluating deer impacts based on the occurrence of browsing scars on seedlings, together with browse preferences. We also validated the percentages of browsed seedlings irrespective of browse preferences as an index of deer impact. We measured the heights of seedlings 30–200 cm tall in five 5 × 20 m plots located in Abies sachalinensis plantations in each of seven tracts and recorded browsing damage that occurred on leader shoots of seedlings. Browse species preference was evaluated using a generalized linear model (GLM) that included the seven tracts, species, and seedling height as explanatory variables. The results of the GLM suggest that differences in browse preferences among species are continuous and cannot be categorized discretely. The tract coefficients are considered to be an index of deer impact levels and can be estimated from the percentage of browsed seedlings for a species if the species coefficient is given. Simple percentages of browsed seedlings can also be used as an index of impact level, regardless of browse preferences, though these may be inaccurate, especially when the site is dominated by significantly preferred or avoided species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Growing deer populations cause serious impacts on many forests throughout temperate regions (Côté et al. 2004; Takatsuki 2009; Reimoser and Putman 2011). The impacts of sika deer (Cervus nippon yesoensis) on natural forests and forest plantations in Hokkaido, Japan, are increasing as deer populations increase (Akashi 2009b; Akashi et al. 2013), and deer population control has become a critical issue for forest management. Estimations of population size and trends are essential for effective population management. Spotlight surveys, catch per unit effort, and sighting per unit effort (SPUE) by hunters have been used to monitor deer population trends in Hokkaido (Uno et al. 2006), and deer abundance in eastern and western Hokkaido has been estimated using harvest-based Bayesian techniques (Yamamura et al. 2008). However, local population densities have not yet been estimated, and given that one of the primary objectives of deer population management is to mitigate deer impacts on natural vegetation, the development of reliable indices of both local deer abundance and impacts on vegetation is a major priority.
Various indices have been developed to evaluate the impacts of deer on vegetation. Projective cover (Fujiki et al. 2010), species composition (Sweetapple and Nugent 2004), browsing on woody seedlings (Frelich and Lorimer 1985; Castleberry et al. 1999; Morellet et al. 2001; Rooney and Waller 2003), and the height of indicator herb species (Anderson 1994; Augustine and de Calesta 2003; Koh et al. 2010) in the understory vegetation have been considered. The forest floor of many forests in Hokkaido is dominated by dwarf bamboo (Sasa spp. and Sasamorpha borealis; Toyooka et al. 1983), which prevents seedling establishment (Umeki and Kikuzawa 1999; Noguchi and Yoshida 2004). This means that the abundance and species composition of seedlings and herbaceous plants depends largely on bamboo cover. Seedling density, projective cover, and species composition reflect the cumulative impact of deer browsing, while current impact levels are needed for deer population management. New browsing scars represent the impact of the current deer population; our previous study evaluated the effects of sika deer using percentages of seedlings browsed in the current year for all broad-leaf species (Akashi et al. 2011).
However, deer browsing preferences vary among tree species (Castleberry et al. 1999; Forsyth et al. 2002; Boulanger et al. 2009; Mosbacher and Williams 2009; Koda and Fujita 2011). While indicative species have been used to obtain a reliable index in other studies (e.g., Morellet et al. 2001; Rooney and Waller 2003; Koh et al. 2010), selecting indicative species that cover most of Hokkaido is not feasible because the flora often differ among sites and no species may be common among all sites (Akashi et al. 2011). Thus, one possible approach is to determine deer preferences for major tree species and to estimate deer impacts at each site from browsing on dominant species, combined with browse preferences. Several studies in Hokkaido have quantitatively identified the tree preferences of deer for bark stripping since scars persist on trunks for a considerable time afterwards (Takayanagi et al. 1991; Takahashi et al. 1997; Sakabe et al. 1998; Kojima et al. 2006; Suzuki et al. 2011), but few have described the browse preference of deer.
The other approach is to use the percentage of browsed seedlings of all species regardless of browse preference as an index. This may be less precise than approaches incorporating browse preference, but in forests with a low seedling density or low abundance of dominant species, field surveys evaluating individual species are time-consuming. Thus, the limitations of simple percentages of browsed seedlings should be recognized based on validation through comparisons with evaluated impact levels combined with browse preferences.
In this study, we reanalyzed data from Akashi et al. (2011). Our objectives were: (1) to determine the woody browse preferences of sika deer, (2) to advance an approach for evaluating deer impacts on forests based on the percentage of browsed seedlings combined with browse preferences, and (3) to examine the validity of the percentage of browsed seedlings of all species regardless of browse preferences as an index of deer impact on forests.
Methods
Study sites and sampling methods
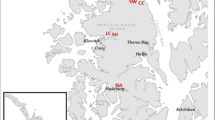
Surveys were conducted from August 31 to October 8, 2009 (Akashi et al. 2011). Our study sites were located in seven tracts within the Hokkaido Prefectural Forests (Akkeshi: AKK; Urahoro: URA; Niikappu: NII; Nishiokoppe: NIS; Mukawa: MUK; Toyokoro: TOY; Tsukigata: TSU; Fig. 1). Within each of the seven tracts, five sites were selected within Sakhalin fir (Abies sachalinensis) plantations aged 40–60 years. These sites were managed according to standard Prefectural Forests procedures; ca. 2500–3000 seedlings ha−1 were planted after clear-cutting and managed with several thinning operations until harvesting at a stand age of 60–80 years. We arbitrarily located a plot measuring 5 × 20 m at each site. We measured the heights of seedlings 30–200 cm tall in each plot, identified their species, and recorded any browsing damage that occurred on leader shoots in the survey year.

Locations of the study tracts. Five sites were established in each of the seven tracts
Sika deer are the only wild ungulate species in Hokkaido. SPUE (the number of deer sighted per hunter-day) is the only index of sika deer abundance that covers nearly all of Hokkaido; it is tallied annually using grid squares of 2′30″ latitude by 3′45″ longitude based on reports from hunters. SPUE values (averaged for 3 × 3 grids) differed among the tracts, from 2.1 to 10.2 deer/hunter-day (2006–2008 hunting seasons; Akashi et al. 2011).
Analysis
We used a generalized linear model (GLM) with a binomial distribution and logit link function to examine browse species preferences based on the presence of browsing scars on seedlings. The GLM included broad-leaf species with ten or more seedlings in total that occurred in two or more tracts and with at least one seedling browsed by deer; seedlings of species that did not meet these criteria were excluded from the analysis. The explanatory variables included tract, species, and seedling height, and variable selection was conducted using Akaike’s information criterion (AIC). The analysis was conducted in R 3.1.2 (R Core Team 2014). The estimated coefficients for each species indicated their browse preference value. Multiple comparisons among the estimated species coefficients were conducted using Tukey’s honest significant difference (HSD) test in the multcomp package in R (Hothorn et al. 2013).
Among the species analyzed in the GLM, the percentages of browsed seedlings of species with five or more seedlings in each tract were plotted against their corresponding preference coefficients to examine the difference in preference for a given species among tracts. The percentages estimated by the GLM at each tract can be calculated as a function of species coefficients if the seedling height is given. The estimates with seedling height of 63 cm (the average height of seedlings analyzed) were overlaid for each tract.
The estimated coefficients for tracts can be considered as indices of deer impact levels. These coefficients and the percentages of browsed seedlings of all species in each tract were compared with the SPUE values by Spearman’s rank correlation coefficient.
Results
Species composition of seedlings
There were 64 species of seedlings in the seven study tracts, including 60 broad-leaf species and 4 conifers. Species composition differed among tracts, and no species occurred in all tracts (Table 1). Dominant seedling species at a given tract were often uncommon or absent in the other tracts. Both the frequency and species richness of seedlings were low at URA and AKK.
The number of seedlings of 2 conifer species and 28 broad-leaf species were less than 10. No browsing was detected for two conifers (A. sachalinensis and Cephalotaxus harringtonia var. nana) and one broad-leaf (Aralia elata) species that had more than 10 seedlings. Five species with 10 or more seedlings occurred only in one tract (Betula ermanii in NIS, Symplocos sawafutagi in TSU, Zanthoxylum piperitum in NII, Sorbaria sorbifolia var. stellipila in AKK, and Staphylea bumalda in TOY).
Browse preference of deer
Browse preferences were analyzed using the GLM for 26 species. The AIC score for the full model containing tract, species, and seedling height was 1187.8 (the lowest); in comparison, the AIC score for the model without seedling height was 1193.2. Thus, all variables were included in the model. Though seedling height was included in the model, 86 % of analyzed seedlings were shorter than 100 cm, and the range of coefficients for seedling height (30–200 cm) was smaller than the coefficients for the tracts and species. The estimated coefficients for each species from the GLM were continuously distributed (Table 2). Tukey’s HSD comparisons showed that the differences between some of the most and least preferred species were significant, but no significant differences were detected among most species (Table 2).
An index of deer impacts on the forest
The percentages of browsed seedlings for each species in each tract were distributed along lines estimated from the GLM and average seedling height, with a few exceptions (e.g., Morus australis in NIS; Fig. 2). The percentages of browsed seedlings for a given species were higher in MUK and NIS than in NII, and few seedlings of less preferred species (species coefficients <0) were browsed in TOY and TSU (Fig. 2).

Percentages of browsed seedlings for species with five or more seedlings in each tract, plotted against the estimated generalized linear model coefficient of species for the occurrence of browsing damage on each seedling. Dashed lines indicate the estimated percentages for 63-cm tall seedlings in each tract. The numbers in parentheses are the numbers of species represented in each tract
The order of coefficients in the GLM for each tract was consistent with the percentages of browsed seedlings for all species, except for the inversion of NII and NIS (Fig. 3). Spearman’s rank correlation coefficient in tracts with SPUE was 0.71 (P = 0.088) for coefficients of the GLM for each tract, and 0.75 (P = 0.066) for percentages of browsed seedlings for all species.

Percentages of browsed seedlings and the estimated coefficient for each tract
Discussion
The browse preferences of plants are often categorized as either preferred, not selected, or avoided (Forsyth et al. 2002; Boulanger et al. 2009; Koda and Fujita 2011), but the classification of a given species can vary among sites (e.g., Koda and Fujita 2011). Some species that were not consumed by deer at one site could be preferred at other sites where more palatable plants were unavailable. The GLM results suggest that the differences in woody browse preferences among species were continuous and could not be categorized discretely, but the rank order of species was generally consistent among tracts (Fig. 2). The browsing probability of seedlings also depended on seedling height, and the GLM evaluated the browse preferences of species while considering this factor.
Among the species with small estimated coefficients, the aboveground parts of Hydrangea serrata var. yesoensis and Rubus idaeus often wizen in winter. Deer typically only browse the leaf blades and petioles of Kalopanax septemlobus and A. elata, which have leaves with long petioles or compound leaves. In these cases, no browsing scar is left on the twigs or stems, resulting in underestimation of their browsing damage. However, species coefficients (Table 2) represent relative preferences for major broad-leaf species in Hokkaido, though the differences in browse preferences among species were not significant for most species. Further data from other areas and more species should be included in the analysis to increase the availability of evaluated preferences.
The estimated GLM coefficients for the tracts (Table 2) represent the impact level combined with browse preferences. The tract coefficient of the GLM with average seedling height can be estimated from the percentage of browsed seedlings for a species if the coefficient of the species is given (Fig. 2). For example, 20 % for species with a coefficient of −1 means that the impact level is the same as at NII, and is equivalent to 70 % for species with a coefficient of 1. Detecting deer impacts at an early stage and controlling their abundance are necessary to maintain forest health (Akashi 2009a), and the percentage of browsed seedlings for preferred species can be used as an indicator of early-stage browsing impacts. Few seedlings of preferred species can be found in tracts with high impact levels, but less preferred species can be used to estimate the impact levels at these sites.
Strong correlations with the SPUE values were not detected, both for the tract coefficients and percentages of browsed seedlings (P > 0.05). This may partly stem from the fact that the data used to determine the SPUE value are restricted to hunting seasons (in most areas, October–March). Sika deer in Hokkaido often exhibit seasonal migration between their summer and winter ranges (Uno and Kaji 2000; Sakuragi et al. 2003; Igota et al. 2004). It is possible that deer impact levels differ between summer and winter.
The difficulty in evaluating impact level from the percentage of browsed seedlings of a species stems from the low density of seedlings in forests dominated by dwarf bamboo and/or in forests with high deer impact levels. Low abundances of dominant species also complicate evaluations based on single species. According to our results, the percentages of browsed seedlings of all species, not considering preferences, were generally consistent with the estimated GLM coefficients for the tracts, which represent the impact level in relation to browse preferences (Fig. 3). This indicates that simple percentages of browsed seedlings can also be used as an effective index of deer impact levels in Hokkaido, regardless of browse preferences. However, because the seedling species composition differed among tracts (Table 1), the results may be inaccurate, especially for sites dominated by significantly preferred or avoided species.
Observations by many involved persons are ideal to evaluate deer impacts at many sites, but identifying plant species is often difficult despite the fact that considering species preferences is preferable when evaluating deer impacts. Monitoring the percentage of browsed seedlings is a simple and affordable method for detecting the early warning signs of negative deer impacts on forests. In conclusion, there is a continuum of species from preferred to avoided, and intensive surveys considering browse preferences are needed to demonstrate the impact of deer on forests; however, extensive surveys using simple methods are also beneficial for understanding the extent of deer impacts, provided that the methods’ limitation are recognized.
References
Akashi N (2009a) Simulation of the effects of deer browsing on forest dynamics. Ecol Res 24:247–255
Akashi N (2009b) Browsing damage by sika deer on trees in young plantations and its relation to relative deer density indices in Hokkaido, Japan. J Jpn For Soc 91:178–183 (in Japanese with English abstract)
Akashi N, Unno A, Terazawa K (2011) Effects of deer abundance on broad-leaf tree seedling establishment in the understory of Abies sachalinensis plantations. J For Res 16:500–508
Akashi N, Fujita M, Watanabe O, Uno H, Ogiwara H (2013) An evaluation of the impact levels of sika deer (Cervus nippon) on natural forests using a simple check list. J Jpn For Soc 95:259–266 (in Japanese with English abstract)
Anderson RC (1994) Height of white-flowered trillium (Trillium grandiflorum) as an index of deer browsing intensity. Ecol Appl 4:104–109
Augustine DJ, de Calesta D (2003) Defining deer overabundance and threats to forest communities: from individual plants to landscape structure. Ecoscience 10:472–486
Boulanger V, Baltzinger C, Saïd S, Ballon P, Picard J-F, Dupouey J-L (2009) Ranking temperate woody species along a gradient of browsing by deer. For Ecol Manag 258:1397–1406
Castleberry SB, Ford WM, Miller KV, Smith WP (1999) White-tailed deer browse preferences in a southern bottomland hardwood forest. South J Appl For 23:78–82
Côté SD, Rooney TP, Tremblay JP, Dussault C, Waller DM (2004) Ecological impacts of deer overabundance. Annu Rev Ecol Evol Syst 35:113–147
Forsyth DM, Coomes DA, Nugent G, Hall GMJ (2002) Diet and diet preferences of introduced ungulates (Order: Artiodactyla) in New Zealand. NZ J Zool 29:323–343
Frelich LE, Lorimer CG (1985) Current and predicted long-term effects of deer browsing in hemlock forests in Michigan, USA. Biol Conserv 34:99–120
Fujiki D, Kishimoto Y, Sakata H (2010) Assessing decline in physical structure of deciduous hardwood forest stands under sika deer grazing using shrub-layer vegetation. J For Res 15:140–144
Hothorn T, Bretz F, Westfall P, Heiberger RM, Schuetzenmeister A (2013) Multcomp: simultaneous inference in general parametric models. Version 1.3-1. http://cran.r-project.org/web/packages/multcomp/
Igota H, Sakuragi M, Uno H, Kaji K, Kaneko M, Akamatsu R, Maekawa K (2004) Seasonal migration patterns of female sika deer in eastern Hokkaido, Japan. Ecol Res 19:169–178
Koda R, Fujita N (2011) Is deer herbivory directly proportional to deer population density? Comparison of deer feeding frequencies among six forests with different deer density. For Ecol Manag 262:432–439
Koh S, Bazely DR, Tanentzap AJ, Voigt DR, Da Silva E (2010) Trillium grandiflorum height is an indicator of white-tailed deer density at local and regional scales. For Ecol Manag 259:1472–1479
Kojima Y, Yasui Y, Orihashi K, Terazawa M, Kamoda S, Kasahara H, Takahashi Y (2006) Relationship between bark preferences of Cervus nippon yesoensis and inner bark components of small-diameter tree trunks. J Jpn For Soc 88:337–341 (in Japanese with English abstract)
Morellet N, Champely S, Gaillard J-M, Ballon P, Boscardin Y (2001) The browsing index: new tool uses browsing pressure to monitor deer populations. Wildl Soc Bull 29:1243–1252
Mosbacher EV, Williams CE (2009) Browse preference and browsing intensity of white-tailed deer (Odocoileus virginianus) in Allegheny high plateau riparian forests, USA. Wildl Biol Pract 5:11–21
Noguchi M, Yoshida T (2004) Tree regeneration in partially cut conifer-hardwood mixed forests in northern Japan: roles of establishment substrate and dwarf bamboo. For Ecol Manag 190:335–344
R Core Team (2014) R: A language and environment for statistical computing. R Foundation for statistical computing, Vienna, Austria. http://www.R-project.org/
Reimoser F, Putman R (2011) Impacts of wild ungulates on vegetation: costs and benefits. In: Putman R, Apollonio M, Andersen R (eds) Ungulate management in Europe: problems and practices. Cambridge University Press, Cambridge, pp 144–191
Rooney TP, Waller DM (2003) Direct and indirect effects of white-tailed deer in forest ecosystems. For Ecol Manag 181:165–176
Sakabe T, Yabe T, Yajima T, Shibuya M, Takahashi K (1998) Tree damage by sika deer in a wintering area in the Iwaobetsu district on the Shiretoko peninsula. Res Bull Hokkaido Univ For 55:113–122 (in Japanese with English abstract)
Sakuragi M, Igota H, Uno H, Kaji K, Kaneko M, Akamatsu R, Maekawa K (2003) Seasonal habitat selection of an expanding sika deer Cervus nippon population in eastern Hokkaido, Japan. Wildl Biol 9:141–153
Suzuki M, Fujimaki A, Kamoda S, Maehara T, Saito H, Matsui M, Iguchi K, Kaji M, Kamata N (2011) Factors affecting the probability of bark-stripping by deer in a natural forest: an analysis based on tree characteristics, deer habitat utilization and environmental conditions. J Jpn For Soc 93:213–219 (in Japanese with English abstract)
Sweetapple PJ, Nugent G (2004) Seedling ratios: a simple method for assessing ungulate impacts on forest understories. Wildl Soc Bull 32:137–147
Takahashi Y, Inukai M, Iguchi K, Takahashi I, Yamamoto H (1997) A case of forest damage caused by sika deer (Cervus nippon yesoensis Heude) in Iwanazawa experimental plot of the Tokyo University Forest in Hokkaido. Trans Mtg Hokkaido Br Jap For Soc 45:84–86 (in Japanese)
Takatsuki S (2009) Effects of sika deer on vegetation in Japan: a review. Biol Conserv 142:1922–1929
Takayanagi A, Furumoto H, Watanabe Y, Sato S, Matsushita K, Ito T (1991) Deer barking in Shiranuka district of Kyoto University Forest in Hokkaido. Rep Kyoto Univ For 22:13–27 (in Japanese)
Toyooka K, Sato A, Ishizuka S (1983) The distribution map of Sasa group in Hokkaido. Hokkaido Branch, Forest Products Research Institute, Sapporo, Japan (in Japanese)
Umeki K, Kikuzawa K (1999) Long-term growth dynamics of natural forests in Hokkaido, northern Japan. J Veg Sci 10:815–824
Uno H, Kaji K (2000) Seasonal movements of female sika deer in eastern Hokkaido, Japan. Mammal Study 25:45–57
Uno H, Kaji K, Saitoh T, Matsuda H, Hirakawa H, Yamamura K, Tamada K (2006) Evaluation of relative density indices for sika deer in eastern Hokkaido, Japan. Ecol Res 21:624–632
Yamamura H, Matsuda H, Yokomizo H, Kaji K, Uno H, Tamada K, Kurumada T, Saitoh T, Hirakawa H (2008) Harvest-based bayesian estimation of sika deer populations using state-space models. Popul Ecol 50:131–144
Acknowledgments
This work was supported by JSPS KAKENHI Grant Number 25450222 and the Mitsui & Co., Ltd. Environment Fund for the project “Development of assessment method on impacts by sika deer to natural vegetation for ecosystem management”.
Author information
Authors and Affiliations
Corresponding author
About this article

Cite this article
Akashi, N., Unno, A. & Terazawa, K. Significance of woody browse preferences in evaluating the impact of sika deer browsing on tree seedlings. J For Res 20, 396–402 (2015). https://doi.org/10.1007/s10310-015-0492-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10310-015-0492-3