
Abstract
In central Argentina, Serrano forest has a long history of fire disturbance; however, the impact of fire on avifauna remains unknown. We compared the avian–habitat relationships in forest patches with low, moderate, and high fire regimes using a community-level (species richness, abundance, ordination and guilds) and species-level (indicator species analysis) approach. In patches under each fire condition, we recorded bird community composition, richness and abundance, and different vegetation structure variables. The site under high-severity fire regime was structurally poor and had been converted from original forest to dense grassland. There, diversity of bird community was low, retaining approximately 30 % of the species present in the least impacted site. Avian assemblage was dominated by generalist and open area birds. Guilds were underrepresented, showing an important reduction of foliage granivorous, nectarivorous, omnivores, and foliage and bark insectivorous, and absence of fly-catchers. Moreover, low abundance of forest understory, midstory, and canopy species and of birds belonging to open and closed nesting guilds was detected. By contrast, under low and moderate-severity fire regimes highest bird diversity as well as highest representativeness of most guilds was observed. Forest bird species were strongly associated with low fire disturbance, whereas moderate fire disturbance was characterized by the presence of forest and generalist species. Given the critical conservation status of Serrano forest in Córdoba, Argentina, habitat restoration and protection of forest relicts could be suitable measures to promote avifauna preservation.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Disturbance plays a critical role in structuring habitat conditions for wild species because it alters vegetation heterogeneity and resource availability (Greenberg et al. 1995; Brotons et al. 2005; Smucker et al. 2005; Adeney et al. 2006). Fire is the major disturbance agent in many ecosystems because it represent an unpredictable threat that can spread across large areas over a short period (Barlow and Peres 2004). Natural ignitions have sparked wildfires from the origin of land plants to the present; however, human-caused ignitions have increased greatly in recent decades, altering fire regimes (e.g., frequency and destructiveness) and distribution (Bar Massada et al. 2009; Pausas and Keeley 2009). Different habitats undergo strong changes in vegetation, which become patches of structurally simple and similar open vegetation that will progressively tend to resume its former structure (Woinarski et al. 1999; Brotons et al. 2005; Pons and Wendenburg 2005; Adeney et al. 2006). This process results in a radical conversion of within-stand vertical structure, reducing niche space and resource availability for native fauna. The urgent nature of the problem makes it essential to know how structural changes in vegetation, resulting from different fire regimes, affect biodiversity; and the key to our knowledge is to have indicators that are measurable, precise, consistent, and sensitive to the phenomenon being tracked (Salafsky et al. 2001, 2002).
Birds are especially sensitive to habitat disturbances, such as fire, because community assemblage is strongly related to vertical and horizontal vegetation structure (MacArthur and MacArthur 1961; Moreira et al. 2003; Isaach et al. 2004; Slik and Van Balen 2006; Sevillano Ríos et al. 2011). The effects of fires on birds, which vary with the different time scales and disturbance regimes, have been widely documented. One of the effects of fire on avifauna is the reduction of nesting sites for cavity birds or for birds that build nests on trees or shrubs (Stribling and Barron 1995; Dwyer and Block 2000; Milesi et al. 2002). Repeated fires also increase the risk of nest predation and parasitism in ground nesting birds due to the reduction of understory vegetation cover (Recher 1997; Blake 2005; Camprodon and Brotons 2006). In addition, fires reduce food availability to avifauna, insectivorous species being one of the most affected groups, particularly those species that forage on the bark and foliage of trees or shrubs (Raphael et al. 1987; Marone 1991; Stribling and Barron 1995; López de Casenave et al. 1998; Milesi et al. 2002; Politi 2003). Conversely, aerial insectivorous and ground granivorous species are benefited by fires because the foliage of trees and shrubs is reduced in burned areas, favoring insect detection, and litter is removed, facilitating seed detection and availability (Marone 1990; Stribling and Barron 1995; Woinarski et al. 1999; Milesi et al. 2002; Politi 2003; Pons and Wendenburg 2005).
Habitat use is another factor frequently related to the effects of fire on birds. For instance, understory birds are one of the groups most vulnerable to forest disturbance, as they are usually sedentary and shy, showing intolerance of canopy gaps (Barlow et al. 2002). In a burned forest, closed canopy and understory are reduced, and niche space and resource availability for interior birds decrease, favoring species turnover to generalists, open forest and edge birds (Magurran 2004; Pons and Wendenburg 2005; Slik and Van Balen 2006; Ukmar et al. 2007). This effect can be exacerbated if post-fire salvage logging is performed in those places (Castro et al. 2010). On the other hand, at the landscape level, moderate fires can create a mosaic with vegetation communities of different successional stages, generating new habitats that can increase bird diversity (Marone 1990; Stribling and Barron 1995; Adeney et al. 2006).
Fire ecology and its effect on bird diversity has been well studied in Australia (e.g., Recher 1997; Woinarski et al. 1999; Leavesley et al. 2010) and the USA (e.g., Fontaine et al. 2009). However, in South America, these studies are comparatively scarce (e.g., Marone 1990; Barlow et al. 2002, 2006); and the avian responses to different fire regimes in forests remain poorly studied.
In central Argentina, the Chaco montane semi-arid forest in the Sierras Chicas of Córdoba (hereafter Serrano forest) provides an excellent opportunity to study the impact of fire on biodiversity. This area is very rich in fauna and flora and represents significant terrestrial ecosystems for biodiversity conservation by TNC (2005), and is a priority area for bird conservation (Beissinger et al. 1996). Serrano forest has a long history of fire disturbance. In the sixteenth century, after the Spanish conquest, new settlers started to disrupt the balance of the systems with increasing ignitions and deforestation to promote livestock raising and crop production (Gavier 2002). These activities have continued into the present with increasing intensity (Díaz et al. 1994; Zak et al. 2008), reducing Serrano forest biodiversity. At present, native forest is reduced to ~5 % of its original area, distributed as a collection of fragments embedded in a matrix of agriculture (Zak et al. 2004). Fragmentation and the resulting increment in forest edge leads to structural changes such as greater tree mortality, reduced living biomass and increased fuel loading. Hence, fire susceptibility poses a special risk to forest fragments (Uhl and Kauffman 1990; Cochrane 2001). The situation is even more serious if we consider that in these forest remnants at least one fire event occurs every year (Secretariat of Environment and Sustainable Development 2009).
Because it is essential to know how loss of forest structure affects avifauna, we assessed the response of bird communities in sites with different fire severity levels. Our objectives were to compare the avian–habitat relationships in forest patches with low, moderate and high-severity fire disturbance, and to identify the bird species characteristic of each fire disturbance condition. We used a community-level (species richness, abundance, ordination) and a species-level (indicator species analysis, ISA) approach to identify bird species associated with the vegetation structure of sites under each fire condition. We also used bird group or guild approach to evaluate the collective response of species to changes in habitat under each fire condition.
Materials and methods
Study area
The study area was located in the Serrano forest in central Argentina (Córdoba Province), which is a subtype of Chaco subtropical semi-arid forest (Zak et al. 2004; Appendix Fig. 1 in supplementary material). The climate is temperate semi-arid with a monsoonal precipitation regime. Average annual rainfall is 750 mm, and is mainly concentrated in the warm season (October–April), with mean maximum and minimum temperatures of 26 and 10 °C, respectively (Capitanelli 1979). This region is considered a semiarid environment because of the high potential for evapo-transpiration, which determines a water deficit during most of the year (Capitanelli 1979). Natural vegetation consists of four vegetation zones: lowlands Chaco forest (plains up to 750 m asl), Serrano forest (hills between 700 and 1,150 m asl), shrubland (between 1,150 and 1,200 m asl) and grassland (usually above 1,200 m asl; Luti et al. 1979; Cabido and Zak 1999; Gavier and Bucher 2004). Serrano forest was characterized by an overstory dominated by 8- to 10-m trees of Schinopsis haenkeana, Lithraea molleoides, Celtis ehrenbergiana, Zanthoxylum coco, and Ruprechtia apetala. The understory was dominated by shrublands of Acacia spp., Schinus spp. and Condalia montana; and ground cover (up to 1 m) composed of herbs, grasses and vines (Luti et al. 1979; Cabido et al. 1991; Gavier and Bucher 2004).
Fieldwork was performed in patches with different post-fire conditions, i.e., ‘natural treatments’. A burned patch was defined as a relatively homogeneous area of similar stand condition (e.g., basal area, species composition) that had the same fire history. Post-fire conditions were classified into low-severity fire regime (LF), moderate-severity fire regime (MF) and high-severity fire regime (HF). LF was an area (ca. 25 ha; 30°50′S; 64°29′W) that had not undergone fire events for more than 60 years. This area is part of an Important Bird Area, AR159 sensu Birdlife International, (Coconier and Di Giácomo 2009). Dominant trees were L. molleoides and S. haenkeana, with a shrub stratum dominated by Acacia caven, A. atramentaria and Schinus fasciculatus. The area under MF (ca. 25 ha; 30°49′S; 64°29′W) experienced three fire events in the last 60 years, two in 1992 and one in 2005. Here, the dominant vegetation structure was the shrub stratum of A. caven and A. atramentaria, and few isolated trees of S. haenkeana and L. molleoides. The HF area (ca. 25 ha; 30°54′S; 64°29′W) experienced many fire events in the last 60 years, with high incidence in the last decade and five fire events from 1996 to 2006. The last one in November 2006, some days before starting bird surveys. In this patch, vegetation was dominated by an herbaceous stratum of Clematis montevidensis, Melica macra, Bidens pilosa and Conyza bonariensis. The studied sites were all in mountainous areas only accessible by hiking or horse riding. None of the sites were subjected to salvage logging practices to our concern, or had significant exotic invasion; there were also no large native herbivorous species present or cattle overgrazing in any site in the last 60 years. Maximum separation between sites was 10 km; all of them share the same bird species pool (sensu Nores 1996). Sites were located in privately owned areas and none of them were legally protected.
Because most of the Serrano forest in the region is in a poor state of conservation (Gavier and Bucher 2004; Barchuk et al. 2009) and is seriously affected by exotic plant invasions (Hoyos et al. 2010; Tecco et al. 2010; Giorgis et al. 2011), overgrazing and urbanization (Gavier and Bucher 2004; Zak et al. 2008), we were unable to find other well preserved forest patches with similar post-fire conditions or habitat characteristics. Although our results could be considered difficult to extrapolate beyond the areas surveyed, the lack of relicts with similar ‘natural treatments’ to truly replicated plots is uncommon in studies on fire effects (e.g., van Mantgem et al. 2001; 2006; Talamo and Caziani 2003; Blake 2005; Pelegrin and Bucher 2010; Rush et al. 2012). Additionally, accumulating local studies is often the only approach for obtaining general patterns (Blake 2005).
Bird surveys
Data were collected during the austral spring and summer (November 10, 2006 to February 23, 2007), when bird richness peaks due to the presence of migrant and resident breeding species. Bird species richness and abundance were estimated using standard point counting techniques, by observers who recorded all birds seen and heard within a 25-m radius for 5 min (Ralph et al. 1993; Bibby et al. 2000). A total of 25 survey points (9 in LF, 9 in MF and 7 in HF) were located at least 150 m apart, to avoid double-counts of the same bird at neighboring points, and at 50 m from the patch edge to reduce its effect. At each point, the observer waited for a settling-down period of 5 min before the beginning of the count to minimize possible negative effects on bird behavior (Ralph et al. 1993; Bibby et al. 2000).
Point count stations were visited twice in each breeding season, always under favorable weather conditions (minimal wind and no rain), and between dawn and 11 a.m., totaling four counts at each point. Nocturnal species (Strigidae and Caprimulgidae) and species that only flew over the forest were not considered. Taxonomical classification follows Remsen et al. (2012). Point locations were recorded with a Garmin 12XL GPS (Garmin Etrex Legend, 2004).
Vegetation structure
Vegetation data were gathered on transects of 25 m in length, always facing east. The initial point on each transect was each bird point count. We sampled the following variables: ground cover, litter depth, understory vegetation structure, canopy cover and tree density. Percentage of ground cover and depth of litter (cm) were estimated using square plots (1 m2), which were situated directly adjacent to the transect line. Ground cover was recorded using the Braun-Blanquet method with six cover categories: (a) <1 %, (b) 1–5 %, (c) 5–25 %, (d) 25–50 %, (e) 50–75 %, and (f) 75–100 % (Kercher et al. 2003). Litter depth was measured with a conventional ruler. Vegetation structure of the understory was recorded with the interception point technique (Hays 1981); three strata were considered: 0–0.5, 0.5–1 and 1–1.5 m. Canopy cover was quantified using an ocular sighting tube (15 cm diameter, Van’t Hul et al. 1997). Tree and snag density (individuals/ha) was estimated with the line transect technique. We recorded the perpendicular distance with respect to the transect line of the ten closest trees (dbh ≥10 cm) or snags (Moreira et al. 2003; Rabinowitz 2003). We also recorded the percentage of fire scars on trees and shrubs.
Data analysis
We used species richness (S) and average individuals’ abundance (n) as measures of biodiversity. Changes in composition of avian communities among the different fire regimes (beta diversity) were tested with the Sorensen qualitative index (Magurran 2004).
Bird species were grouped into different guilds (Appendix Table 1 in supplementary material) according to the literature (Jácome 1996; Nores 1996; López de Casenave et al. 1998; Blendinger 2005; Dardanelli et al. 2006a, b). We refer to the term ‘guild’ as the group of birds that exploit the same resources in a similar way and belong to the same communities (Magurran 2004). Grouping bird species that are functionally similar is more effective for developing management guidelines than examining species individually (Canterbury et al. 2000; Mitchell et al. 2006), because intra-guild richness reflects integrity of an area (Bishop and Myers 2005). Birds were grouped into three groups of guilds: diet, nesting, and habitat use. Diet guilds included: terrestrial-insectivorous, TI (insectivorous birds that feed mainly on the ground); foliage and bark insectivorous, FI (birds that feed on insects mostly in shrub and tree foliage and on the bark); aerial insectivorous and fly-catchers, F (birds that catch insects in the air); terrestrial granivorous, TG (birds that feed mainly on seeds on the ground); foliage granivorous, FG (birds that feed mainly on seeds and fruits on the foliage); nectarivorous, N, and omnivores, O. Nesting guilds included: open-nesting birds (open nest above the ground), closed-nesting birds (cavities and domed nests above the ground) and ground-nesting birds (any nest on the ground). Habitat use guilds included: forest understory species, FU (species that use mainly the understorey or the ground of the forest); forest midstory and canopy species, FMC (species that use mainly the medium stratum and the canopy of the forest); generalist species, G (species that inhabit forests and open areas equally); and open areas species, OA.
Data normality and variance homogeneity were tested using Saphiro Wilks and Levene tests, respectively. Data expressed in percentages were previously arcsine transformed and species richness and abundance were square-root transformed to adjust normal distribution assumptions (Quinn and Keough 2002). We used one-way ANOVA to test for differences in overall bird richness and bird abundance among fire regimes (factor with 3 levels: HF, MF, LF) and Duncan’s post hoc test to assess differences among fire regimes (Quinn and Keough 2002). Spatial autocorrelation of ANOVA residuals was tested using the autocorrelation module provided by Past v 1.89 (Hammer et al. 2009) and no correlation was found (Appendix Fig. 2 in supplementary material for details). Moreover, we used one-way ANOVA-Duncan tests to check for differences in abundance of bird species per guild and vegetation characteristics among fire regimes (HF, MF, LF, Quinn and Keough 2002).
We also calculated the indicator value (IV) index (Dufrene and Legendre 1997) to find species significantly associated with a particular burn regime within the bird assemblage (i.e., LF, MF and HF) following the formula:
where A ij , is the mean abundance of species i in the treatment of group j compared to all groups in the study. B ij is the relative frequency of occurrence of the species i in the sites of groups j. Final multiplication by 100 produce percentages. An IV = 100 % shows that a species is a perfect indicator to a given habitat typology (for details, see Dufrene and Legendre 1997). Indicator values were tested for significance with a Monte Carlo randomization procedure (Quinn and Keough 2002).
Finally, we ran a canonical correspondence analysis (CCA) to relate forest structure and avifauna (measured as richness and abundance of birds per community and guilds) of each fire regime. The significance of the CCA was evaluated using a Monte Carlo permutation test; then, Spearman’s correlation test was used to assess the association among independent variables and the CCA axes selected (Conover 1999). Statistical analyses were performed with Infostat v.2010 (Di Rienzo et al. 2010).
Results
Habitat variables
Mean attributes of vegetation structure differed among sites under different fire regimes. Arboreal density, shrub cover in the 1–1.5 m understory, and litter depth were inversely related to the fire disturbance regimes (i.e., LF > MF > HF; Table 1), whereas percentages of fire scars were directly related (Table 1). The main differences were recorded in the HF site, which was structurally poor and dominated by grass species in the 0–0.5 m understory cover. Ground quality was also poor in the HF site, with low percentage of cover and lack of litter (Table 1). The remaining vegetation variables were similar in MF and LF sites, except for canopy cover, which was better represented in LF than in MF (Table 1). We did not find differences in snag density among fire regimes (Table 1).
Bird communities
We recorded 53 bird species; 41 species and 350 individuals were detected in the LF site, 47 species and 473 individuals in the MF site, and 17 bird species and 114 individuals in the HF site (Appendix Table 1 in supplementary material). Bird richness and abundance varied among sites (F = 19.25; p < 0.0001), the HF site showing the lowest values (Appendix Table 1 in supplementary material).
Sorensen’s similarity index showed that species turnover increased with increasing severity of the disturbance. LF and MF sites shared 89 % of the bird species, LF and HF shared 45 %, and HF and MF sites shared 44 % of species (Appendix Table 1 in supplementary material). Four species were found only in the LF site; eight were detected only in the MF and three were present only in the HF site. Thirteen species were ubiquitous (Appendix Table 1 in supplementary material).
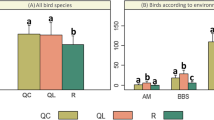
Birds per guild also showed variation among fire regimes (Fig. 1): abundance of terrestrial granivorous (F = 5.92; P = 0.009), foliage and bark insectivorous (F = 25.08; P < 0.0001), and ground-nesting (F = 9.88; P = 0.0009) species was highest in MF, with significant differences from the remaining sites (Fig. 1). The HF site was characterized by the low abundance of foliage granivorous (F = 13.23; P = 0.0002), nectarivorous (F = 20.31; P < 0.0001), omnivores (F = 29.14; P < 0.0001) foliage and bark insectivorous (F = 25.08; P < 0.0001), and by the absence of fly-catchers (Fig. 1). The HF site also had the lowest abundance of forest understory (F = 18.41; P < 0.0001), forest midstory and canopy species (F = 33.85; P < 0.0001) and of birds belonging to open (F = 53.04; P < 0.0001) and closed (F = 9.88; P = 0.0009) nesting guilds (Fig. 1). The LF site had the lowest richness of open area species (F = 19.39; P < 0.0001, Fig. 1). No differences were recorded in the guild of birds generalist in habitat use (F = 3.07; P = 0.07), and terrestrial insectivores (F = 1.66; P = 0.21, Fig. 1).

Bird mean abundance (±SE) by guilds in Serrano forest (central Argentina) sites with different fire regimes. Diet: TI terrestrial-insectivores, FI foliage insectivores, F fly-catchers, BI bark-insectivores, TG terrestrial granivorous, FG foliage granivorous, N nectarivorous and O omnivores. Habitat use: FU forest understory species, FMC forest midstory and canopy species, G generalist species and OA species from open areas and/or forest edge and nesting guilds. Black bars High fire regime, grey bars medium fire regimes and white bars low fire regimes. Asterisk indicates significant differences on fire regimes based on Duncan test (P < 0.05)
Species-habitat associations
Canonical correspondence analysis (CCA) showed significant correlation among bird species and habitat variables for two canonical axes (CCA axis 1: eigenvalue = 0.32; P = 0.02; CCA axis 2: eigenvalue = 0.27; r = 0.95, P = 0.01). Axis 1 was positively associated with the 0–0.5 m understory cover (Spearman r = 0.69; P < 0.05) and percentage of fire scars (r = 0.46; P < 0.05) and negatively with the 1–1.5 m understory cover (r = −0.77; P < 0.05), tree density (r = −0.8; P < 0.05) and percentage of ground cover (r = −0.57; P < 0.05). Axis 2 was positively associated with litter depth (r = 0.75; P < 0.05), 0.5–1 m understory cover (r = 0.56; P < 0.01), and canopy cover (r = 0.73; P < 0.05); in addition, axis 2 had a negative association with 0–0.5 m understory cover (r = 0.6; P < 0.05; Appendix Fig. 3 in supplementary material). Snag density had a low correlation (r = 0.22 and r = −0.15; P < 0.05) with axis 1 and 2 respectively.
Four bird species were positively associated with axis 1 (Appendix Fig. 4 in supplementary material), and hence 0–0.5 m understory cover and fire scars. Eleven bird species were negatively associated with axis 1 (Appendix Fig. 4 in supplementary material). Four species were positively related to CCA axis 2 (Appendix Fig. 4 in supplementary material), and consequently to litter depth, 0.5–1 understory cover and canopy cover; whereas seven bird species were negatively associated with axis 2 (Appendix Fig. 4 in supplementary material).
Indicator species analysis
Nine bird species were selected as significant (Monte Carlo test based on 1000 permutations; P < 0.05) indicators of (or species closely associated with) a particular habitat type. We considered six bird species as marginally significant (P < 0.10; Table 2) to the conservative level of significance used in the present work. One species was closely associated with the LF site, eight species with the MF site, and no species were significantly associated (P < 0.05) with the HF site; however, two species showed a marginal association with this site (Table 2).
Discussion
This work is a first approach to the understanding of the effects of fire regimes on birds of Serrano forest in central Argentina and serves as a starting point for further comprehensive studies. Our results show that local severity of fire, qualitatively measured as low, moderate and high severity regimes, had a strong effect on avifaunal composition. The highest contrast in bird diversity, species composition and guild representation was driven by the impoverishment of habitat attributes induced by repeated fires. High-severity fire disturbance (HF site) produced the conversion from original forest to dense grassland with the encroachment of the lower understory and reduction of tree density, litter depth and ground cover. In HF, avifauna was noticeably different, retaining only approximately 30 % of the species present in the least impacted site (LF), and with a significant turnover of birds to disturbance tolerant species or successional species (generalists and open area species).
Similar results were found in the dry forest of Madagascar, where recurrent fires turned a rich specialized forest avifauna, with some restricted range species, into one of opportunistic and open habitat birds, characteristic of savannas (Pons and Wendenburg 2005). Leavesley et al. (2010) found a similar pattern in dry Mulga woodlands in Australia, which was transformed into grassland by repeated burning, with loss of specialist birds, and generalist bird species dominating the assemblages.
The lack of significant differences in avian diversity between LF and MF sites could be related to the similarity in habitat structure between sites. Albeit the lack of differences in diversity, the small variations in tree density and, hence, canopy cover of the forest in the MF site led to the presence of eight species (most of them belonging to midstory and canopy species) that were not recorded in the LF site (Appendix Table 1 in supplementary material).
Structural changes of habitat attributes due to severity of fire events also produced variations in guild composition. Within feeding guilds, the oversimplification of habitat structure in the HF site notably reduced the diversity of food items, leading the reduction in numbers of foliage granivorous, omnivorous; nectarivorous and foliage and bark insectivorous. Similar results were found by Marone (1991) and Milesi et al. (2002) in Monte desert shrublands in western Argentina, and by López de Casenave et al. (1998) and Politi (2003) in lowland dry Chaco forest in northern and central Argentina. Fly-catchers were absent from the HF site possibly because of the lack of trees and shrubs used for perching, and the reduction of flying insect biomass available (Meehan and George 2003). By contrast, birds that forage on the foliage and the bark of trees and shrubs and terrestrial granivorous species were much more abundant at the moderately disturbed site (MF). Insectivorous birds were favored probably because of the increase in habitat heterogeneity; Leavesley et al. (2010) also found that insectivorous birds were dominant in the intermediate disturbance situation in burned Mulga woodlands in Australia. In turn, the terrestrial granivorous birds increased due to better accessibility and detection of grains on the ground due to reduced litter depth, lower tree density and lower upper understory cover. This phenomenon has also been suggested for other similar studies in pine forests in the USA (Raphael et al. 1987; Stribling and Barron 1995), in temperate spruce and pine forest in Sweden (Edenius 2011), in savannas in northern Australia (Woinarski et al. 1999) and in Chaco lowland forest in central Argentina (Politi 2003). The lack of differences in terrestrial insectivorous among sites could be related to the low density of these birds in the entire study area.
Regarding habitat use, the loss of structural complexity of vegetation in the HF site reduced the occurrence and abundance of forest bird species, favoring the proliferation of habitat generalists and open area species, which are probably evolutionarily adapted to disturbances caused by fire (Edenius 2011). Similar results were detected by Ukmar et al. (2007) and Battisti et al. (2008) in pinewoods in Italy, by Slik and Van Balen (2006) in a lowland tropical rainforest of eastern Borneo and by Pons and Wendenburg (2005) in dry forests in Madagascar.
In general, nesting guilds show diverse responses to fire regimes (Saab and Powell 2005; Rush et al. 2012). In our study, LF and MF sites presented the largest number of closed and open nesting birds that use trees and shrubs to nest. Both groups were notably underrepresented in the HF site due to the absence of canopy and understory nesting substrate for these species. The abundance of ground nesting birds was higher in the MF site, probably because of a greater heterogeneity of micro-sites than in the LF site and, in turn, greater availability of resources than in the HF site. These findings are in agreement with observations in Alabama piedmont (Stribling and Barron 1995) and pine forests of northern Arizona, USA (Dwyer and Block 2000), and the Monte desert shrublands in western Argentina (Milesi et al. 2002).
In agreement with avian diversity and guild patterns, indicator bird species of each site were also strongly related to the degree of habitat disturbance. The species characteristic or indicator of low disturbance by fire (LF) was Elaenia parvirostris, a forest species that uses the upper understory, midstory and canopy strata. This species has also been found to be sensitive to other types of habitat alteration, like forest exploitation (selective logging and understory alteration by livestock breeding) in coastal woodlands of eastern-central Argentina (Cueto and Lopez de Casenave 2000), and to shrub removal in Chaco dry woodlands in northern Argentina (Codesido et al. 2009). Additionally, E. parvirostris also prefers native forest without exotic species to forest severely invaded by introduced trees and shrubs in central-eastern Argentina (Fandiño et al. 2010).
Bird species that were best associated with the habitat type of the MF site included a generalist species, like Sappho sparganura, a species that uses the forest understory, such as Synallaxis frontalis, but also species that use more than one forest strata, such as Chlorostilbon lucidus, Asthenes baeri, Myiophobus fasciatus, Saltatricula multicolor, Poospiza nigrorufa, and Geothlypis aequinoctialis. Some other studies in Argentina have shown similar results. S. multicolor disappeared or its density was severely reduced in sites affected by fires, where tree and shrub cover were poor or absent, in desert scrublands in central-western Argentina (Milesi et al. 2002). A. baeri has been found to be sensitive to shrub removal in Chaco dry forests in northern Argentina (Codesido et al. 2009). S. frontalis was reported as sensitive to forest exploitation (selective logging and understory alteration by livestock grazing) in coastal woodlands of eastern-central Argentina (Cueto and Lopez de Casenave 2000) and to shrub removal in Chaco dry forests in northern Argentina (Codesido et al. 2009). G. aequinoctialis was also found to be associated with intermediate disturbance (a fire occurred 8 years earlier) in the Chaco lowland forest in central Argentina (Politi 2003). On the other hand, C. lucidus preferred unburned Chaco lowland forest in central Argentina (Politi 2003) but showed tolerance to shrub removal in the same type of forest in northern Argentina (Codesido et al. 2009) and can live in exotic pine plantations with no shrubs (Dardanelli 2003). Additionally, most of these species were resilient to forest fragmentation in lowland woodlands in central Argentina (Dardanelli et al. 2006a).
No species were selected as significant indicators of high disturbance by fires. However, if we relax the significance level (to p < 0.1) one generalist species, Troglodytes aedon, and one open area species, Embernagra platensis, appear as indicators. T. aedon seems to be a species adapted to successional conditions, as it is considered a very common and generalist species in central Argentina (Nores 1996) and was found to prefer burned forests (Politi 2003) and areas where the shrub stratum was experimentally removed in Chaco lowland dry forests in northern Argentina (Codesido et al. 2009). E. platensis also seems to be adapted to successional conditions. It is considered a common open area species in central Argentina (Nores 1996) and behaves as a forest edge species in highly fragmented lowland dry forests (Dardanelli et al. 2006b).
In conclusion, the loss of forest bird diversity and simplification of assemblage structure was directly related to the local severity of fire disturbance, as has been observed in similar studies (e.g., Ukmar et al. 2007, Battisti et al. 2008). A moderate-severity fire regime, in turn, was not detrimental to avian diversity; in fact, foliage and bark insectivorous, terrestrial granivorous and ground nesting birds were best represented under these conditions. However, some forest species were not able to cope with this moderate level of forest disturbance, and needed a less disturbed forest to persist. Moreover, recurrent fires in an already relictual ecoregion like the Serrano forest which has gone through a dramatic deforestation and fragmentation process (Gavier and Bucher 2004; Zak et al. 2004, 2008), leave very few habitats available to forest bird species, especially for those sensitive to area loss (Dardanelli et al. 2006a). More severe effects on the avifauna will probably be evident in the future because of the extinction debt (Tilman et al. 1994). In this scenario, restoration of Serrano forest appears to be an appropriate tool for biodiversity recovery; hence, further research is needed to better understand habitat successional dynamics in an ecosystem that has experienced different fire regimes. The creation of more protected areas in the region should also be a high priority action for biodiversity conservation, especially considering that this ecosystem is one of the richest in Córdoba Province.
References
Adeney JM, Ginsberg JR, Russell GJ, Kinnaird MF (2006) Effect of an ENSO-related fire on birds of a lowland tropical forest in Sumatra. Anim Conserv 9:292–301
Bar Massada A, Radeloff VC, Stewart SI, Hawbaker TJ (2009) Wildfire risk in the wildland–urban interface: a simulation study in northwestern Wisconsin. For Ecol Manag 258:1990–1999
Barchuk AH, Britos AH, De la Mata E (2009) Diagnosis of native forest cover in the province of Cordoba in Geographic Information System. Research Group Participatory Land report, Universidad Nacional of Cordoba, Cordoba, Argentina
Barlow J, Peres CA (2004) Avifaunal response to single and recurrent wildfires in Amazonian forests. Ecol Appl 14:1358–1373
Barlow J, Haugaasen T, Peres CA (2002) Effects of surface wildfires on understorey bird assemblage in Amazonian forests. Biol Conserv 105:157–169
Barlow J, Peres CA, Henriques LMP, Stouffer PC, Wunderle JM (2006) The responses of understorey birds to forest fragmentation, logging and wildfires: an Amazonian synthesis. Biol Conserv 128:182–192
Battisti C, Ukmar E, Luiselli L, Bologna MA (2008) Diversity/dominance diagrams show that fire disrupts the evenness in Mediterranean pinewood forest bird assemblages. Community Ecol 9:107–113
Beissinger SR, Steadman EC, Wohlgenant T, Blake JG, Zack S (1996) Null models for assessing ecosystem conservation priorities: threatened birds as titers of threatened ecosystems in South America. Conserv Biol 10:1343–1352
Bibby CJ, Burgess ND, Hill DA, Mustoe S (2000) Bird census techniques. Academic Press, London
Bishop JA, Myers WL (2005) Associations between avian functional guild response and regional landscape properties for conservation planning. Ecol Indic 5:33–48
Blake JG (2005) Effects of prescribed burning on distribution and abundance of birds in a closed-canopy oak-dominated forest, Missouri, USA. Biol Conserv 121:519–531
Blendinger PG (2005) Abundance and diversity of small-bird assemblages in the Monte desert, Argentina. J Arid Environ 61:567–587
Brotons L, Pons P, Herrando S (2005) Colonization of dynamic: where do birds come from after fire? J Biogeogr 32:789–798
Cabido M, Zak M (1999) Vegetation of north of Cordoba. Report of the Secretariat of Agriculture, Livestock and Natural Resources of Cordoba
Cabido M, Carranza ML, Costa A, Páez S (1991) Contribution to the phytosociological knowledge of the Chaco Belt forest in Córdoba Province, Argentina. Phytocoenologia 19:547–566
Camprodon J, Brotons L (2006) Effects of undergrowth clearing on the bird communities of the Northwestern Mediterranean Coppice Holm oak forest. For Ecol Manag 221:72–82
Canterbury GE, Martin TE, Petit DR, Petit LJ, Bradford DF (2000) Bird communities and habitat as ecological indicators of forest condition in regional monitoring. Conserv Biol 14:544–558
Capitanelli RC (1979) Climate. In: Vazquez JB, Miatello RA, Roque ME (eds) Physical geography of Córdoba province. Boldt, Buenos Aires, pp 144–203
Castro J, Moreno-Rueda G, Hódar JA (2010) Experimental test of postfire management in pine forests: impact of salvage logging versus partial cutting and nonintervention on bird species assemblages. Conserv Biol 24:810–819
Cochrane MA (2001) Synergistic interactions between habitat fragmentation and fire in evergreen tropical forest. Conserv Biol 15:1515–1521
Coconier EG, Di Giácomo AS (2009) Argentina. In Devenish C, Díaz Fernández DF, Clay RP, Davidson I, Yépez Zabala I (eds) Important bird areas Americas: priority sites for biodiversity conservation. BirdLife Conservation Series No. 16. BirdLife International, Quito, pp 59–70
Codesido M, Drozd AA, Gado PA, Bilenca D (2009) Responses of a bird assemblage to manual shrub removal in a Chacoan subtropical semiarid forest, Argentina. Ornitol Neotrop 20:47–60
Conover WJ (1999) Practical Nonparametric Statistics. Wiley, New York
Cueto VR, Lopez de Casenave J (2000) Bird assemblages of protected and exploited coastal woodlands in east-central Argentina. Wilson Bull 112:396–403
Dardanelli S (2003) The role of pine plantations as habitat for birds and the impact on natural environments of Cordoba, Argentina. Master Thesis. Universidad Nacional of Cordoba, Cordoba, Argentina (in Spanish)
Dardanelli S, Nores ML, Nores M (2006a) Minimum area requirements of breeding birds in fragmented woodland of central Argentina. Divers Distrib 12:687–693
Dardanelli S, Serra DA, Nores M (2006b) Composition and abundance of the avifauna of woodland fragments in Córdoba, Argentina. Acta Zool Lilloana 50:71–83
Di Rienzo JA, Casanoves F, Balzarini MG, Gonzalez L, Tablada M, Robledo CW (2010) InfoStat version 2010. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina
Díaz S, Acosta A, Cabido M (1994) Community structure in montane grasslands of central Argentina in relation to land use. J Veg Sci 5:483–488
Dufrene M, Legendre P (1997) Species assemblage and indicator species: the need for a flexible asymmetrical approach. Ecol Monogr 67:345–366
Dwyer JK, Block VW (2000) Effects of wildfire on densities of secondary cavity nesting birds in ponderosa pine forests of northern Arizona. In: Kieth M, Moser CF (eds) Fire and forest ecology: innovative silviculture and vegetation managements. Tall Timbers Fire Ecology Conference Proceedings, No. 21. Tall Timbers Research Station, Tallahassee, pp 151–156
Edenius L (2011) Short-term effects of wildfire on bird assemblages in old pine- and spruce-dominated forests in northern Sweden. Ornis Fenn 88:71–79
Fandiño B, Berduc AJ, Beltzer AH (2010) Avian assemblages in native and exotic forests in the reproductive season in a protected area of El Espinal de Entre Ríos, Argentina. Ornitol Neotrop 21:1–16
Fontaine JB, Donato DC, Robinson WD, Lae BE, Kauffman JB (2009) Bird communities following high-severity fire: response to single and repeat fires in a mixed-evergreen forest, Oregon, USA. For Ecol Manag 257:496–1504
Gavier GI (2002) Deforestation and forest fragmentation in the Sierras Chicas of Cordoba, Argentina. Master Thesis. Universidad Nacional of Cordoba, Cordoba, Argentina (in Spanish)
Gavier GI, Bucher EH (2004) Cordoba montane forest loss and fragmentation between 1970 and 1997. Acad Nac Cs (Cordoba, Argentina) Miscelánea N° 101
Giorgis M, Tecco P, Cingolani AM, Renison D, Marcora P, Paiaro V (2011) Factors associated with woody alien species distribution in a newly invaded mountain system of central Argentina. Biol Invasions 13:1423–1434
Greenberg CH, Lawrence DH, Neary DG (1995) A comparison of bird communities in burned and salvage-logged, clearcut, and forested Florida sand pine scrub. Wilson J Ornithol 107:40–50
Hammer O, Harper DAT, Ryan PD (2009) PAST v.1.89. Palaeontological Statistics. Free download http://www.nhm.uio.no/norges/past/download.htm
Hays RL (1981) Estimating wildlife habitat variables. Fish and Wildlife Service. Biological Services Program FWS/OBS-81/47
Hoyos L, Gavier-Pizarro GI, Kuemmerle T, Bucher EH, Radeloff VC, Tecco PA (2010) Invasion of glossy privet (Ligustrum lucidum) and native forest loss in the Sierras Chicas of Córdoba, Argentina. Biol Invasions 12:3261–3275
Isaach JP, Holz SL, Ricci L, Martínez MM (2004) Post-fire vegetation change and bird use of a salt marsh in coastal Argentina. Wetlands 24:235–243
Jácome MR (1996) Avifauna of humid ravines of the Sierras Chicas of Cordoba: current status and guidelines for management. Master Thesis. Universidad Nacional of Cordoba, Córdoba, Argentina (in Spanish)
Kercher SM, Frieswyk CB, Zedler JB (2003) Effects of sampling teams and estimation methods on the assessment of plant cover. J Veg Sci 14:899–906
Leavesley AJ, Cary GJ, Edwards GP, Gill AM (2010) The effect of fire on birds of Mulga woodland in arid central Australia. Int J Wildl Fire 19:949–960
López de Casenave J, Pelloto JP, Caziani SM, Mermoz M, Protomastro J (1998) Responses of avian assemblages to a natural edge in a Chaco semiarid forest in Argentina. Auk 115:425–435
Luti R, Solis M, Galera FM, Muller N, Berzal M, Nores M, Herrera M, Barrera JC (1979) Vegetation. In: Vasquez J, Miatello R, Roque M (eds) Physical geography of the province of Córdoba. Boldt, Buenos Aires, pp 297–368
MacArthur RH, MacArthur JW (1961) On bird species diversity. Ecology 42:594–598
Magurran A (2004) Measuring biological diversity. Blackwell Publishing, Malden
Marone L (1990) Modifications of local and regional bird diversity after a fire in the Monte Desert, Argentina. Rev Chil Hist Nat 63:187–195
Marone L (1991) Habitat features affecting bird species distribution in the Monte desert, Argentina. Ecol Aust 1:77–86
Meehan TD, George TL (2003) Short-term effects of moderate- to high-severity wildfire on a disturbance-dependent flycatcher in Northwest California. Auk 120:1102–1113
Milesi FA, Marone L, López De Casenave J, Cueto VR, Mezquida ET (2002) Management guilds as indicators of environmental conditions: a case study with birds and habitat disturbances in the central Monte desert, Argentina. Ecol Aust 12:149–161
Mitchell MS, Rutzmoser SH, Wigley TB, Loehle C, Gerwin JA, Keyser PD, Lancia RA, Perry RW, Reynolds C, Thill RE, Weih R, White D, Wood PB (2006) Relationships between avian richness and landscape structure at multiple scales using multiple landscapes. For Ecol Manag 221:155–169
Moreira F, Delgado A, Ferreira S, Borralho R, Oliveira N, Inácio M, Silva JS, Rego F (2003) Effect of prescribed fire on vegetation structure and breeding birds in young Pinus pinaster stands of northern Portugal. For Ecol Manag 184:225–237
Nores M (1996) Avifauna of the province of Cordoba. In: di Tada E, Bucher EH (eds) Biodiversity of the province of Córdoba. Universidad Nacional of Río Cuarto, Cordoba, pp 255–337
Pausas JG, Keeley JE (2009) A burning story: the role of fire in the history of life. Bioscience 59:593–601
Pelegrin N, Bucher EH (2010) Long-term effects of a wildfire on a lizard assemblage in the arid Chaco forest. J Arid Environ 74:368–372
Politi N (2003) Degradation and fire in the arid Chaco: implications for bird assemblage. Master Thesis. Universidad Nacional of Cordoba, Cordoba, Argentina (in Spanish)
Pons P, Wendenburg C (2005) The impact of fire and forest into savanna on the bird communities of West Madagascan dry forests. Anim Conserv 8:183–193
Quinn GP, Keough MJ (2002) Experimental design and data analysis for biologists. Cambridge University Press, London
Rabinowitz AR (2003) Wildlife field research and conservation training manual. Wildlife Conservation Society, New York
Ralph CJ, Geupel GR, Pyle P, Martin TE, Desante DF (1993) Handbook of field methods for monitoring landbirds. USDA Forest Service General Technical Report PSW-144. Albany, CA
Raphael MG, Morrison ML, Yoder-Williams MP (1987) Breeding bird populations during twenty-five years of postfire succession in the Sierra Nevada. Condor 89:614–626
Recher HF (1997) Impact of wildfire on the avifauna of Kings Park, Perth, Western Australia. Wildl Res 24:745–761
Remsen JV, Cadena JR, Jaramillo CD, Nores M, Pacheco JF, Robbins MB, Schulenberg, TS, Stiles FG, Stotz DF, Zimmer KJ (2011) A classification of the bird species of South America. American Ornithologists’ Union. http://www.museum.lsu.edu/~Remsen/SACCBaseline.htm.. Accessed 1 Aug 2012
Rush S, Klaus N, Keyes T, Petrick J, Cooper R (2012) Fire severity has mixed benefits to breeding bird species in the southern Appalachians. For Ecol Manag 263:94–100
Saab V, Powell DW (2005) Fire and avian ecology in North America: process influencing pattern. Stud Avian Biol 30:1–13
Salafsky N, Cauley H, Balachander G, Cordes B, Parks J, Margoluis C, Bhatt S, Encarnacion C, Russell D, Margoluis R (2001) A systematic test of an enterprise strategy for community-based biodiversity conservation. Conserv Biol 15:1585–1595
Salafsky N, Margoluis R, Redford KH, Robinson JG (2002) Improving the practice of conservation: a conceptual framework and research agenda for conservation science. Conserv Biol 16:1469–1479
Secretariat of Environment and Sustainable Development (2009) Forest fire statistics 2001–2009. http://www.ambiente.gov.ar. Accessed 15 June 2011
Sevillano Ríos SS, Lloyd H, Valdéz-Velásquez A (2011) Bird species richness, diversity and abundance in Polylepis woodlands, Huascaran biosphere reserve, Peru. Stud Neotrop Fauna E 46:69–76
Slik JWF, Van Balen S (2006) Bird community changes in response to single and repeated fires in a lowland tropical rainforest of eastern Borneo. Biodivers Conserv 15:4425–4451
Smucker KM, Hutto RL, Steele BM (2005) Changes in birds abundance after wildfire: importance of severity and time since fire. Ecol Appl 15:1535–1549
Stribling HL, Barron MG (1995) Short-term effects of cool and hot prescribed burning on breeding songbird populations in the Alabama piedmont. Sout J Appl For 19:18–22
Talamo A, Caziani SM (2003) Variation in woody vegetation among sites with different disturbance histories in the Argentine Chaco. For Ecol Manag 184:79–92
Tecco PA, Díaz S, Cabido M, Urcelay C (2010) Functional traits of alien plants across contrasting climatic and land-use regimes: do aliens join the locals or try harder than them? J Ecol 98:17–27
Tilman D, May RM, Lehman CL, Nowak MA (1994) Habitat destruction and the extinction debt. Nature 371:65–66
TNC (2005) Gran Chaco americano ecoregional assessment. Fundación Vida Silvestre Argentina, Fundación para el Desarrollo Sustentable del Chaco (DeSdel Chaco) and Wildife Conservation Society Bolivia (WCS). 1st edn. The Nature Conservancy, Buenos Aires
Uhl C, Kauffman JB (1990) Deforestation, fire susceptibility, and potential tree responses to fire in the eastern Amazon. Ecology 71:437–449
Ukmar E, Battisti C, Luiselli L, Bologna MA (2007) The effect of fire on communities, guilds and species of breeding birds in burnt and control pinewoods in central Italy. Biodivers Conserv 16:3287–3300
Van Mantgem PJ, Schwartz M, Keifer M (2001) Monitoring fire effects for managed burns and wildfires: coming to terms with pseudoreplication. Nat Area J 21:266–273
Van Mantgem PJ, Stephenson NL, Keeley JE (2006) Forest reproduction along a climatic gradient in the Sierra Nevada, California. For Ecol Manag 225:391–399
Van’t Hul JT, Lutz RS, Mathews NE (1997) Impact of prescribed burning on vegetation and bird abundance at Matagorda Island, Texas. J Range Manag 50:346–350
Woinarski JCZ, Brock C, Fisher A, Milne D, Oliver B (1999) Response of birds and reptiles to fire regimes on pastoral land in the Victoria River District, Northern territory. Rangel J 21:24–38
Zak MR, Cabido M, Hodgson JG (2004) Do subtropical seasonal forests in the Gran Chaco, Argentina, have a future? Biol Conserv 120:589–598
Zak MR, Cabido M, Cáceres D, Díaz S (2008) What drives accelerated land cover change in central Argentina? Synergistic consequences of climatic, socioeconomic, and technological factors. Environ Manag 42:181–189
Acknowledgments
We acknowledge financial support from the Agencia Córdoba Ambiente, CONICET (PIP N° 11220090100263) and Secyt-UNC. We are grateful to the Tessi family for allowing us to conduct fieldwork on their properties. We also thank J. Heredia and L. Volkmann for their field assistance; C. Trujillo and J. Brasca for helping us with the English edition and the JFR editor and two anonymous reviewers for providing helpful comments that improved the manuscript. L. M. B. is a researcher from CONICET and S. D. had a fellowship from CONICET.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
About this article
Cite this article
Albanesi, S., Dardanelli, S. & Bellis, L.M. Effects of fire disturbance on bird communities and species of mountain Serrano forest in central Argentina. J For Res 19, 105–114 (2014). https://doi.org/10.1007/s10310-012-0388-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10310-012-0388-4