
Abstract
Sudden oak death is a disease affecting coastal forests in California and southern Oregon. The spatial pattern of disease dynamics is important for forest and landscape pathology; in this work we investigated the interaction across landscape scales of disease symptomology in coast live oaks, Quercus agrifolia, (trunk bleeding, presence of beetles, and presence of the fungus Hypoxylon thouarsianum) and tree mortality through time. We used two-dimensional spatial analysis tools with data gathered in point-centered-quarter format in 2001 and 2004 to quantify the population density of the disease through time; to examine the spatial pattern of tree mortality across scales through time; and to examine the spatial co-occurrence of disease symptoms with crown mortality through time. Early in the study period dead trees were strongly clustered at smaller scales (~300 m) and after three years this clustering was less pronounced. Bleeding on trees occurred in clusters away from dead trees, particularly in 2004, likely indicating a new cohort of infected trees. The presence of H. thouarsianum was strongly related to overstory mortality through time. Beetle-infested trees co-occurred with mortality in 2001. By 2004, they occurred throughout the forest, and were less strongly correlated with overstory tree mortality, suggesting a future peak of tree mortality.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
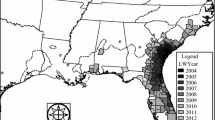
The pathogen Phytophthora ramorum is the causal agent of “sudden oak death” (SOD), a forest disease that has reached epidemic levels in hardwood and mixed hardwood forests in 14 counties in central coastal California (CA) and one county in southern Oregon (OR) (Fig. 1). The disease has two forms, a frequently fatal tree disease which has killed tens of thousands of oak and tanoak trees (coast live oak, Quercus agrifolia, California black oak, Q. kelloggii, Shreve oak, Q. parvula var. shrevei, and tanoak, Lithocarpus densiflorus) and non-fatal diseases commonly called “foliar and twig dieback” which affect more than 25 foliar hosts including tree and shrub species (Rizzo 2003; Rizzo and Garbelotto 2003; Davidson et al. 2005; McPherson et al. 2005). The foliar hosts play an important role in short-range pathogen dispersal within forests, with their leaves serving as reservoirs for the pathogen spores which can be spread to soil and leaf litter via rain, and short distances through a forest by wind-driven rain (Davidson et al. 2002, 2005; Davidson and Shaw 2003; Rizzo and Garbelotto 2003). The primary symptom of sudden oak death on affected trees is the production of a viscous red to black exudate from bark cankers, typically on the lower trunk, often referred to as bleeding. The bleeding areas on such trees are often colonized by bark and ambrosia beetles (Svihra 1999), and may exhibit the fruiting bodies (sporophores) of the native fungus, Hypoxylon thouarsianum (McPherson et al. 2000 McPherson et al. 2005). This symptom progression was established by McPherson et al. (2005) who monitored disease symptoms and associated organisms on SOD host trees (Quercus kelloggii, Q. agrifolia and Lithocarpus densiflorus) in Marin County, CA, USA, in 20 plots quarterly over four years.

a Distribution of sudden oak death in California and b China Camp State Park study area in Marin County, CA, USA, showing transect locations
Like other recent forest pathogen outbreaks globally, sudden oak death has occurred over regional rather than local scales, displaying striking patchiness across the landscape (Kelly and Meentemeyer 2002; Rizzo and Garbelotto 2003). The study of this disease within forest systems thus necessarily involves larger-scale approaches to complement traditional individual tree-based forest pathological research (Holdenrieder et al. 2004). To this end, there have been studies examining the progression of disease symptoms through time across a gradient of the disease (McPherson et al. 2005), and examining the spatial pattern of disease-related mortality at large scales (Kelly and Meentemeyer 2002), but there has been little work examining the interplay of disease symptomology and mortality across landscape scales through time. Quantification of the interaction of disease symptoms and mortality across time and space is an important addition to our knowledge of disease establishment and spread, and can be performed using an array of spatial analytical techniques.
Ecologists and biogeographers often use randomly located plots or arrays of one-dimensional samples to analyze patterns. Such data can be analyzed using spatial statistics tools such as Ripley’s K statistic, which is defined as the expected number of individuals within a distance of a randomly chosen individual in a population (Ripley 1976; Kenkel 1988, 1994; Cressie 1993). The method has been used to study vegetation mortality patterns and tree interactions in numerous systems (Andersen 1992; Szwagrzyk and Czerwczak 1993; Vacek and Lepš 1996; Cole and Syms 1999; Eccles et al. 1999). The method is not ideal in all situations, as it requires complete enumeration of the target (e.g., all trees in a study area must be analyzed). This is not always practical over large scales. In such cases, researchers can use either a remote sensing approach or a more comprehensive field-based approach to more completely enumerate a target prior to analysis. For example, Kelly and Meentemeyer (2002) and Liu et al. (2006) used remote sensing and spatial analysis to quantify the clustering of overstory mortality associated with sudden oak death across large scales. A remote sensing approach cannot capture understory or tree-specific symptomology, and thus two-dimensional transect data are commonly used because researchers can cover large areas while capturing vegetation detail and spatial location, thus adequately capturing landscape-scale patterns in vegetation distribution (Wilson and Gurevitch 1995; Akashi 1996; Ma et al. 2001; Bryant et al. 2005).
This work uses data gathered from “point-centered-quarter” (Engeman et al. 1994) transects to first quantify the population density of trees with different disease symptoms across the study area, and second to analyze the spatial patterning of the disease to determine the scale of clustering of symptoms across space, the degree to which disease symptoms tended to co-occur in the forest, and the extent to which these patterns changed through time.
Materials and methods
Study area
China Camp State Park (CCSP) is located on the southwestern shore of San Pablo Bay (latitude = 38.00059°, longitude = 122.48514°) near the city of San Rafael in California (Fig. 1). Elevation varies from near sea level to 290 m. The dominant overstory forest trees are coast live oak, black oak, bay laurel (Umbellularia californica), and madrone (Arbutus menziesii). Additional overstory species include valley oak (Q. lobata), blue oak (Q. douglasii), California buckeye (Aesculus californica), Douglas-fir (Pseudotsuga menziesii), big leaf maple (Acer macrophyllum), Eucalyptus spp., and coast redwood (Sequoia sempervirens). The vegetation cover varies considerably, comprising open fields, chaparral, dense shrub thickets, and relatively open, oak-dominated closed canopy forests. Common shrubs, small trees, and vines include manzanita (Arctostaphylos manzanita), toyon (Heteromeles arbutifolia), hazel (Corylus cornuta), buckeye (Aesculus californica), coffeeberry (Rhamnus californica), huckleberry (Vaccinium ovatum), rhododendron (Rhododendron macrophyllum), poison oak (Toxicodendron diversilobum), and honeysuckle (Lonicera hispidula). With the exception of the white oaks, Q. douglasii and Q. lobata, and Eucalyptus spp., these woody plants have all been confirmed as hosts of P. ramorum (Davidson et al. 2003).
Point-centered quarter sampling
The point-centered quarter (PCQ) method of transect data collection is one of the most popular vegetation sampling strategies (Cottam and Curtis 1956). We collected data using the PCQ method in CCSP in the summer of 2001, and each tree was re-evaluated in 2004. Parallel linear transects were established approximately 500 m apart, aligned in a southerly direction from North San Pedro Road (see Fig. 1), which served as the anchor point for the seven transects. Sampling center points (nodes) were located every 100 m on each transect, with the first node starting 100 m from the anchor. Transects ranged from 2 to 28 nodes in length. One short transect was offset due to inaccessible terrain and not anchored to the road. The location of each center point was recorded with a global positioning system (GPS) device. At each node, cardinal compass directions defined four quadrants. The closest coast live oak or black oak (>5 cm dbh) in each quadrant was identified to species, labeled with a unique tag number, and dbh and disease status were determined. The distance from node center and azimuth of each tree were recorded. Dead trees were included unless they showed evidence of significant decay and significant loss of limbs, indicating they had been dead for more than about five years. Only trees that were within a maximum distance of 30 m from a node center were recorded.
Symptom evaluation
Visual disease symptoms for each of the trees at each node were evaluated on a scale of one to five as: 1, asymptomatic; 2, bleeding only; 3, bleeding plus beetles; 4, bleeding plus beetles plus Hypoxylon; 5, dead (following McPherson et al. 2005). These are referred to in the paper as “symptom levels”. We analyzed the data from the PCQ method in two ways: we first estimated the population density of trees across the study area, and then we examined the spatial pattern of crown mortality and associated symptoms. The data from the PCQ nodes were formatted in slightly different ways in both cases.
Population density estimation
The population density of oak trees at each of five different symptom levels was estimated from the PCQ data in three steps. First, the mean of all node center-to-plant distances (the mean distance from the center node to all four sample trees) was calculated for all sample points, regardless of symptom, using the following method: \( \left( {\bar d} \right){\text{m}} \) (the mean point-to-plant distance) is formulated as: \( \bar d = \frac{1}{n}\sum\nolimits_{{i = 1}}^{n} {d_{i} } , \) where n is the overall sample size. The squared value of this mean distance gives the mean grid area occupied by one tree. Second, the population density of oak trees at all symptom levels can be estimated as the ratio between a unit area (1 ha) and the mean grid area. The overall population density of oak trees ρ (tree/ha) was calculated as: \( \rho = \frac{{10,000\;{\text{m}}^{ 2} / {\text{ha}}}}{{\left( {\bar dm} \right)^{2} / {\rm{tree}}}} \) given the units of mean distance and area are meters and square meters, respectively. Finally, the population density of oak trees at a particular symptom level is calculated as the product of the overall population density ρ and the percentage of trees at that symptom level. Thus, the population density of symptom level i is denoted by ρ i , and ρ i = ρ × p i , where \( p_{i} = \frac{{n_{i} }}{n} \times 100\% , \) n i , and n are the sample size of trees at symptom level i (i = 1, 2, 3, 4, 5) and the overall sample size, respectively.
Spatial pattern analysis of transect data
Quadrat variance methods such as blocked quadrat variance (BQV), local quadrat variances (LQV), paired quadrat variances (PQV), and new local variances (NLV) are commonly applied to detect spatial patterns in plant communities from transect data (some examples include: Hill 1973; Usher 1983; Carter and O’Connor 1991; Schaefer 1993; Ver Hoef et al. 1993; Schaefer and Messier 1994; Edwards et al. 1996; Dai and van der Marrel 1997; Dale 1999; Ribeiro and Fernandes 2000; Guo and Kelly 2004). These methods essentially analyze the spatial pattern from one-dimensional data and require data to be recorded on a series of quadrats along a transect. Our original transect data were collected using a PCQ sampling method, where each transect consists of a number of nodes with four observations recorded at each node’s four quarters. In order to apply the quadrat variance methods to the transect data for detection of the scale of disease clustering, we adjusted the PCQ transect data in two steps: first we treated the nodes along each transect as the centers of the quadrats, and the width of quadrats is determined as the maximum point-to-plant distance, and, second, the quarter-wise observations were converted to quadrat-wise observations by replacing the four-quarter records with the mean or maximum value for each node.
We applied two quadrat variance analysis methods including paired quadrat variances (PQV) and two-term local quadrat variances (TTLQV), to the adjusted transect data. PQV calculates the mean squared difference of two paired quadrats at varying distance apart whereas TTLQV calculates the mean squared difference of adjacent blocks of quadrats with varying block sizes. The two methods are defined in the following formulas (Dale and MacIsaac 1989; Dale 1999):
where x j is the observation on the j-th quadrat; n is total number of quadrats; r is the scale parameter, which is the integer-times of the quadrat size. The peaks of the variance in V PQV and V TTLQV can be interpreted as the scale of the pattern. In our case, the peak in each graph represents the size of the clustering, and the height of the peak indicates strength of clustering.
Paired quadrat variances is mathematically identical to a one-dimensional variogram analysis, detecting the scale of the pattern under investigation (Bryant et al. 2005). Just as the variogram can be extended to a cross-variogram for co-variance analysis, the PQV can also be extended to a paired quadrat covariance (PQC) in order to study the co-variance among oak trees at different symptom levels. To do this, the PCQ transect data were adjusted as follows: for each symptom level, the quadrat value at each node was recorded as 1 or 0 (presence or absence of a symptom). This generated five binary transects of quadrats, each of which corresponded to one symptom. The PQC is similarly defined in the formula:
where x i,k and x j,k are the presence/absence values on the k-th quadrat for symptom level i and j, respectively; n is total number of quadrats; r is the scale parameter, which is the integer-times of the quadrat size.
We used the formulae above to analyze data along the longest transect in our study area. This transect is 2,800 m in length. Since we were examining spatial patterning across landscape scales, we decided to use only the longest transect for this portion of the work; analysis on the other transects would yield less useful patterns, as they are shorter.
Results
Population density estimation
The mean node center-to-plant distance (for all coast live oaks) is 10.13 m, and the population density of all oak trees in the study area is 97.48 trees/ha. The tree density characteristics and the mean disease state of coast live oaks are reported for transects that were at least ten nodes in length in Table 1. Although the mean tree density of the transect that was analyzed here was greater than the mean of the other transects, the incidence of sudden oak death symptoms was similar to that in the other transects. The population density of trees at different symptom levels in both years is reported in Table 2. The number, percentage and density of trees with all symptoms except crown mortality declined between 2001 and 2004. In other words, trees with no symptoms, trees with bleeding, trees with bleeding and beetles, and trees with bleeding, beetles, and Hypoxylon were less frequently found in 2004 than in 2001. The number of nodes with dead crowns increased between 2001 and 2004, from 46 to 86.
Spatial pattern analysis of transect data
The results from both the PQV and TTLQV methods of transect analysis are very similar. Both the PQV and TTLQV plots show strong clustering of overstory mortality in 2001, and less pronounced clustering in 2004 (Fig. 2). PQV and TTLQV plots of both mean symptom number of 5 (trees from all four quadrants at a node had dead crowns) and maximum of 5 (at least one of the trees in the four quadrants at a node had a dead crown) in 2001 showed strong clustering at 300 m. Both plots also reveal a weaker larger-scale clustering in overstory mortality in 2001; the PQV shows clustering of overstory mortality (for nodes with one or more dead trees) at 1000 m and the TTLQV shows this clustering at 1,100 m. In 2004, there was substantially less spatial pattern in overstory mortality despite the increase in numbers of dead trees.

Result of transect analysis for clustering of overstory mortality: a PQV analysis of the longest transect using the mean symptom from each node in both years, and the maximum symptom from each node in both years; b TTQLV analysis of the longest transect using the mean symptom from each node in both years, and the maximum symptom from each node in both years
We compared co-occurrence in space for trees with the symptoms: bleeding and crown mortality (2 and 5), beetles and crown mortality (3 and 5), beetles plus Hypoxylon and crown mortality (4 and 5) by using PQC analysis (Fig. 3), and we review the results symptom-by-symptom. For both years, we see an increasingly strong association of symptom state with mortality as tree transition progresses from bleeding, to beetle infestation, to the presence of Hypoxylon. The strength of the association with mortality is in relation to the “0” variance x-axis, where a strong association is positive and a weak association is negative. Bleeding: Earlier in the epidemic in 2001, bleeding did not co-occur with dead trees; bleeding trees existed in small-scale clusters (250 and 600 m in size) away from major clusters of dead trees. By 2004 this relationship was even more pronounced: bleeding occurred away from dead trees, on new clusters of trees across all spatial scales, indicating perhaps a new cohort of infected trees in the forest. Beetles: In 2001, beetle-infested trees did not co-occur with mortality at small scales, but were associated with dead trees in clusters from 500–700 m, and over 1,000 m. In 2004, beetle-infested trees did not display much spatial patterning, and were found throughout the forest, except at larger scales (over 1,200 m), where beetle-infested trees did co-occur with dead trees. The presence of beetles on trees is a stronger predictor of large-scale mortality than bleeding alone. Hypoxylon: Hypoxylon on trees was strongly correlated with dead trees across all scales early in the infestation, and again in 2004, although slightly less strongly.

Results of PQC: a data comparing nodes with bleeding and mortality, symptom 5, and symptoms 4 and 5 for 2001; b data comparing nodes with symptoms 2 and 5, symptoms 3 and 5, and symptoms 4 and 5 for 2004
Discussion
This work provides an estimate of the density of symptomatic trees at different levels of disease across a forested landscape affected by sudden oak death. Earlier in the epidemic, dead trees were strongly clustered at smaller scales (~300 m) and, despite an increase in dead crowns later in the epidemic, this clustering was less pronounced in 2004. These patterns support those found using remotely sensed data across the park. For example, we have been mapping overstory crown mortality through time in this study area, and have used the data to quantify spatial pattern of mortality across landscape scales (Kelly and Meentemeyer 2002; Kelly et al. 2004; Liu et al. 2006; Guo et al. 2007). We found in those studies clusters of overstory tree mortality of 200–400 m in size.
The relationship between disease symptom classes and oak mortality across several spatial scales, examined using the PQC plots, may indicate future waves of mortality. The data show that the symptom progression described in McPherson et al. (2005) of bleeding, followed by beetle attack, followed by Hypoxylon infestation, also exists spatially, with some differences. Bleeding trees occurred in clusters away from existing dead trees; beetles-infested trees were associated with mortality in the earlier (2001) evaluation but in the later (2004) evaluation these were more evenly distributed throughout the forest and were less correlated with mortality. The association of beetle-infested trees with dead trees in 2001 likely reflects the outcome of earlier clusters of infected trees. Because of the temporal variability associated with the median survival estimates, this association implies that these trees represent the remnant survivors from an earlier infection episode. The presence of Hypoxylon was strongly related to mortality consistently through the study period. This association is supported by other studies showing that trees exhibiting H. thouarsianum fruiting bodies represent the final disease stage before death. Because bleeding is the first visual symptom of infection the PQC analysis provides evidence of a potential new cohort of infected trees. Survival models (McPherson et al. 2005) found that median expected survival declined significantly once infected trees were attacked by beetles. Thus, the spatial distribution of beetle-infested trees should be strongly correlated with trees exhibiting H. thouarsianum, and subsequently, with mortality. This result supports the importance of long-term monitoring of forest diseases. While the initial striking pattern of dead trees was less pronounced in 2004 than in 2001, the distribution of trees in advanced disease stages insures there will be considerable additional mortality in the future (McPherson et al. 2005).
The use of transects to capture spatial patterning of vegetation across landscapes is common in ecology. Many have used transect data to analyze spatial patterning of heterogeneous vegetation, but such data can be misinterpreted. To help minimize these problems, Guo and Kelly (2004) recommend doing more than one quadrat variance test to insure correct interpretation of patch or gap size, and we have done so. While the transect method allowed us to capture much of the spatial heterogeneity across the forest here, we were constrained in our spatial analysis by the orientation of the park: we covered the forests with north-south oriented transects, and only one of the transects had sufficient length to perform all of the analyses that we performed here. Still, we are confident that these analyses capture the important spatial patterns across the park.
As with any forest pathogen, P. ramorum exists in the context of biotic and abiotic factors that may affect its distribution. In structurally heterogeneous forests, humidity and temperature may influence local abundance and infectivity of this pathogen (Garbelotto et al. 2003; Condeso and Meentemeyer 2007). The distribution of host species may also constrain or facilitate the spread of P. ramorum. Bay laurel foliar infections may provide a major source of inoculum in these forests (Davidson et al. 2002; Kelly and Meentemeyer 2002; Swiecki and Bernhardt 2002; Davidson et al. 2005; Condeso and Meentemeyer 2007). However, the broad host range of P. ramorum suggests that a number of other species may serve as sources of spore production. Further, bay laurel has not been consistently found associated with infected oaks (Maloney et al. 2005; Condeso and Meentemeyer 2007).
In conclusion, this paper reports on spatial and temporal patterning of symptoms associated with sudden oak death across landscape scales (hundreds of meters) in China Camp State Park, CA, USA. We quantify the clustering of dead trees across the landscape, and then investigate the relationship between disease symptoms and dead trees through space and time. We show that earlier in the course of the epidemic, overstory mortality shows a strong clustered pattern at about 300 m. Three years later there are more dead trees, but the clustering is less pronounced. We suggest that the temporal symptom progression documented elsewhere (e.g., in McPherson et al. 2005) has important spatial implications: we show that bleeding occurs early, separate from dead trees, that beetle-attacked trees exist throughout the forest, and that Hypoxylon is strongly related to tree mortality in space. This conclusion is consistent with a pattern of aggregated disease establishment followed by outward expansion of new infections. The fact that the spatial distribution of trees displaying bleeding shows more variation in 2004 than in 2001 suggests that continuing infections of new trees is occurring in sites where infected trees were less abundant.
References
Akashi N (1996) The spatial pattern and canopy-understory association of trees in a cool temperate, mixed forest in western Japan. Ecol Res 11:311–319
Andersen M (1992) Spatial analysis of two-species interactions. Oecologia 91:134–140
Bryant DM, Ducey MJ, Innes JC, Lee TD, Eckert RT, Zarin DJ (2005) Forest community analysis and the point-centered quarter method. Plant Ecol 175:193–203
Carter AJ, O’Connor TG (1991) A two-phase mosaic in a savanna grassland. J Veg Sci 2:231–236
Cole RG, Syms C (1999) Using spatial pattern analysis to distinguish causes of mortality: an example from kelp in north-eastern New Zealand. J Ecol 87:963–972
Condeso TE, Meentemeyer RK (2007) Effects of landscape heterogeneity on the emerging forest disease sudden oak death. J Ecol 95:364–375
Cottam G, Curtis JT (1956) The use of distance measures in phytosociological sampling. Ecology 37:451–460
Cressie NAC (1993) Statistics for spatial data. Wiley, New York
Dai X, van der Marrel E (1997) Transect-based patch size frequency analysis. J Veg Sci 8:865–872
Dale MRT (1999) Spatial pattern analysis in plant ecology. Cambridge University Press, Cambridge
Dale MRT, MacIsaac DA (1989) New methods for the analysis of spatial pattern in vegetation. J Ecol 77:78–91
Davidson JM, Shaw CG (2003) Pathways of movement for Phytophthora ramorum, the causal agent of Sudden Oak Death. In: Proceedings of sudden oak death online symposium. American Phytopathological Society, http://www.apsnet.org/online/SOD
Davidson JM, Rizzo DM, Garbelotto M, Tjosvold S, Slaughter GW (2002) Phytophthora ramorum and sudden oak death in California: II. transmission and survival. In: Proceedings of fifth symposium on oak woodlands: oak woodlands in California’s changing landscape. USDA Forest Service, San Diego, pp 741–749
Davidson JM, Werres S, Garbelotto M, Hansen EM, Rizzo DM (2003) Sudden Oak Death and associated diseases caused by Phytophthora ramorum. Plant Health Progress. doi:10.1094/PHP-2003-0707-01-DG
Davidson JM, Wickland AC, Patterson HA, Falk KR, Rizzo DM (2005) Transmission of Phytophthora ramorum in mixed-evergreen forest in California. Phytopathology 95:587–596
Eccles NS, Esler KJ, Cowling RM (1999) Spatial pattern analysis in Namaqualand desert plant communities: evidence for general positive interactions. Plant Ecol 142:71–85
Edwards GR, Parsons AJ, Newman JA, Wright IA (1996) The spatial pattern of vegetation in cut and grazed grass/white clover pastures. Grass Forage Sci 51:219–231
Engeman RM, Sugihara RT, Pank LF, Dusenberry WE (1994) A comparison of plotless density estimators using Monte Carlo simulation. Ecology 75:1769–1779
Garbelotto M, Davidson JM, Ivors K, Maloney PE, Hüberli D, Koike ST, Rizzo DM (2003) Non-oak native plants are main hosts for sudden oak death pathogen in California. Calif Agric 57:18–23
Guo QH, Kelly M (2004) Interpretation of scale in paired quadrat variance methods. J Veg Sci 15:763–770
Guo Q, Kelly M, Gong P, Liu D (2007) An object-based classification approach in mapping tree mortality using high spatial resolution imagery. GISci Remote Sens 44:24–47
Hill MO (1973) The intensity of spatial pattern in plant communities. J Ecol 61:225–235
Holdenrieder O, Pautasso M, Weisberg PJ, Lonsdale D (2004) Tree diseases and landscape processes: the challenge of landscape pathology. Trends Ecol Evol 19:446–452
Kelly M, Meentemeyer RK (2002) Landscape dynamics of the spread of sudden oak death. Photogram Eng Remote Sens 68:1001–1009
Kelly M, Shaari D, Guo QH, Liu DS (2004) A comparison of standard and hybrid classifier methods for mapping hardwood mortality in areas affected by “sudden oak death”. Photogram Eng Remote Sens 70:1229–1239
Kenkel NC (1988) Pattern of self-thinning in jack pine: testing the random mortality hypothesis. Ecology 69:1017–1024
Kenkel NC (1994) Bivariate pattern analysis of jack pine-trembling aspen interaction. Abstr Bot 18:49–55
Liu D, Kelly M, Gong P (2006) A spatial-temporal approach to monitoring forest disease spread using multi-temporal high spatial resolution imagery. Remote Sens Environ 101:167–180
Ma KM, Zu YG, Godron M (2001) Scale correlation between vegetation and soil in larch forest, NE China. J Environ Sci (China) 13:51–57
Maloney PE, Lynch SC, Kane SF, Jensen CE, Rizzo DM (2005) Establishment of an emerging generalist pathogen in redwood forest communities. J Ecol 93:899–905
McPherson BA, Wood DL, Storer AJ, Svihra P, Rizzo DM, Kelly NM, Standiford RB (2000) Oak mortality syndrome: sudden death of oaks and tanoaks. In: Tree notes, no 26. California Department of Forestry and Fire Protection, Sacramento, pp1–6
McPherson BA, Mori SR, Wood DL, Storer AJ, Svihra P, Kelly NM, Standiford RB (2005) Sudden oak death in California: disease progression in oaks and tanoaks. For Ecol Manage 213:71–89
Ribeiro KT, Fernandes GW (2000) Patterns of abundance of a narrow endemic species in a tropical and infertile montane habitat. Plant Ecol 147:205–217
Ripley BD (1976) The second-order analysis of stationary point processes. J Appl Probab 13:255–266
Rizzo DM (2003) Sudden Oak Death: host plants in forest ecosystems in California and Oregon. In: Proceedings of sudden oak death online symposium. American Phytopathological Society, http://www.apsnet.org/online/SOD
Rizzo DM, Garbelotto M (2003) Sudden oak death: endangering California and Oregon forest ecosystems. Front Ecol Environ 1:197–204
Schaefer JA (1993) Spatial patterns in taiga plant communities following fire. Can J Bot 71:1568–1573
Schaefer JA, Messier F (1994) A paired-quadrat method for use in multiscale ordination. Plant Ecol 113:9–11
Svihra P (1999) Sudden death of tanoak, Lithocarpus densiflorus. In: Pest alert, no 1. University of California Cooperative Extension, Novato, pp 1–2
Swiecki T, Bernhardt E (2002) Evaluation of stem water potential and other tree and stand variables as risk factors for Phytophthora ramorum canker development in coast live oak. In: Proceedings of fifth symposium on oak woodlands: oak woodlands in California’s changing landscape. USDA Forest Service, San Diego, pp 787–798
Szwagrzyk J, Czerwczak M (1993) Spatial patterns of trees in natural forests of East-Central Europe. J Veg Sci 4:469–476
Usher MB (1983) Pattern in the simple moss-turf communities of the sub-Antarctic and maritime Antarctic. J Ecol 71:945–958
Vacek S, Lepš J (1996) Spatial dynamics of forest decline: the role of neighbouring trees. J Veg Sci 7:789–798
Ver Hoef JM, Cressie NAC, Glenn-Lewin DC (1993) Spatial models for spatial statistics-some unification. J Veg Sci 4:441–452
Wilson C, Gurevitch J (1995) Plant size and spatial pattern in a natural population of Myosotis micrantha. J Veg Sci 6:847–852
Acknowledgments
We would like to acknowledge financial support from University of California Division of Agriculture and Natural Resources, USDA Exotic pests and Disease Research Program (fund number: 03XN020), and USDA Cooperative State Research, Education, and Extension Service (fund number: 34519–14392).
Author information
Authors and Affiliations
Corresponding author
About this article
Cite this article
Kelly, M., Liu, D., McPherson, B. et al. Spatial pattern dynamics of oak mortality and associated disease symptoms in a California hardwood forest affected by sudden oak death. J For Res 13, 312–319 (2008). https://doi.org/10.1007/s10310-008-0083-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10310-008-0083-7