
Abstract
Nitrogen (N) input by atmospheric deposition and human activity enhances the availability of N in various ecosystems, which may further affect N and phosphorus (P) cycling and use by plants. However, the internal use of N, P, and N:P stoichiometry by plants in response to N supply, particularly for grass species in a desert steppe ecosystem, remains unclear. In this work, a field experiment was conducted at an infertile area in a desert steppe to investigate the effects of N fertilizer addition rates on the stoichiometry of N and P in a dominant grass species, Seriphidium korovinii. Results showed that for both aboveground and fine roots of S. korovinii, N inputs exponentially increased the N concentration and N:P ratios while P concentrations decreased. Meanwhile, the relationships between N and P concentrations for both aboveground and fine roots were significantly negative. Furthermore, while the N concentrations in the plants were relatively low, P concentrations were higher than the global means, resulting in a relatively low N:P ratio. These results suggest that the stoichiometric characteristics of N were different from that of P for this desert plant species. Results also show that the intraspecific variations in the main element traits (N, P, and N:P ratios) were consistent at the whole-plant level. Our results also suggest that N should be part of any short-term fertilization plan that is part of a management strategy designed to restore degraded desert grassland. These findings highlight that nutrient addition by atmospheric N deposition and human activity can have significant effects on the internal use of N and P by plants. Therefore, establishing a nutrient-conservation strategy for desert grasslands is important.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Nitrogen (N) and phosphorus (P) are the main nutrient elements in terrestrial ecosystems, and these elements control many biogeochemical processes (Lü et al. 2013; Sterner and Elser 2002). Global climate changes and human activities have influenced the dynamics of nutrient cycling in most ecosystems (Aerts et al. 2007; Kozovits et al. 2007). The deposition of atmospheric N has rapidly increased as a result of increased human activities and global climate change, and this deposition has led to an increased availability of N in various ecosystems (Galloway et al. 2004).Moreover, N fertilization is widely used as a primary means of improving grassland productivity and restoring degraded grasslands (Li et al. 2015). In previous years, numerous studies have drawn their attention toward the stoichiometry of plants, which could reflect the nutrient cycling dynamics and function of ecosystems under N fertilized conditions (Lü et al. 2013; Van Heerwaarden et al. 2003).
Increased N availability in soils can alter plant community composition through its effects on nutrient cycling and storage in plants (Li et al. 2016). In general, the addition of N fertilizer enhances the availability of inorganic soil N while increasing the N concentration and N:P ratio in plant green leaves (Lü et al. 2012a; Van Heerwaarden et al. 2003; Xia and Wan 2008). However, Kozovits et al. (2007) reported that four years of N addition did not significantly affect N concentrations in four of five plant species grown on a dystrophic soil in central Brazil. Some field experiments were conducted to investigate the effects of N addition on green leaf P concentrations. However, results are conflicting with either positive (Lü et al. 2013), negative or no significant effects (Van Heerwaarden et al. 2003). These inconsistent results indicated that plant nutrient concentrations are influenced by site- and species-specific nutrient availability under various environmental conditions. Considering these diverse results, the majority of the existing studies have not demonstrated a clear relationship between tissue chemistry with both stoichiometric ratios and N supply among plants. Hence, additional evidence from various types of ecosystems is needed to identify the general patterns in plant tissue stoichiometry in response to N addition.
Nutrient availability is an important factor in controlling plant productivity. Fine roots, usually defined as roots that are <2 mm in diameter, are important for the uptake and internal use of nutrients in plants of terrestrial ecosystems (Gordon and Jackson 2000; Ostertag 2001; Yuan et al. 2011). The nutrient dynamics and status of aboveground biomass, which includes leaves, stems, and reproductive organs, are important in plant nutrient economy and ecosystem nutrient cycling (Lü et al. 2012b). Hence, the dynamic changes of nutrient concentrations in the aboveground and fine roots of plants in response to N fertilization should be investigated. In previous years, researchers have recognized that a strong link exists between the above- and below-ground components of plants (Geng et al. 2014). This observation signifies that N and P concentrations as well as the N:P ratios of leaves and fine roots are positively correlated (Geng et al. 2014; Holdaway et al. 2011; Liu et al. 2010). However, the relationship between aboveground biomass and the fine root stoichiometry of plants has not been extensively analyzed.
The desert grassland on the northern slope of Kunlun Mountain in northwest China contains infertile sites with low vegetation coverage dominated by Seriphidium korovinii (Poljak.) Poliak. While data related to the deposition of atmospheric N are unavailable in this area, studies from other grasslands in China suggest atmospheric N deposition and N input caused by human activity can have a significant effect on nutrient and water use in the ecosystem (Lü et al. 2012b, 2013). A field experiment was conducted at the grasslands of Kunlun Mountain to test the following hypotheses: (1) artificial N addition would increase N concentrations while decreasing P concentrations of aboveground and fine roots of S. korovinii; (2) for both N and P, the stoichiometric relationship between aboveground and fine roots are positively correlated.
Materials and methods
Study site and experimental design
The investigation was performed in the desert grassland of Kunlun Mountain in south central Xinjiang Autonomous Region in northwest China at an elevation of 2400 m (36°22′54"N, 80°43′38"E). The meteorological data were collected from 2011 to 2013 by weather station. The mean annual temperature was 3 °C, and the mean annual precipitation varied from 80 to 100 mm. More than 85% of the total precipitation occurred during the growing season from May to October. The brown desert soil supported a vegetation type of desert grasses. The study area had been fenced since 2009 to prevent the grazing of large animals. The perennial grass S. korovinii dominated the plant community at this site, and represented almost 80% of the total vegetation. S. korovinii is the main source of forage for local livestock, and its growing period is mainly between April and September. Other companion species also occur here including Corispermum mongolicum Iljin, Bassia dasyphylla (Fisch. & C. A. Mey.) Kuntze, and Ceratoides lateens (J.F.Gmel.) Revealet Holmgren.
This study was conducted from May to September 2013. Five levels of N addition were used in the experiment (i.e., 0, 0.4, 0.8, 1.6, and 3.2 mol N m−2 year−1of urea), and 0.05 mol P m−2 of KH2PO4 was added to each plot (Lü et al. 2013; Yu et al. 2010). Treatments were laid out in a randomized block design with four replicates of 4 × 4 m plots. The four replicates of five nitrogen addition treatments created a total of these 20 plots. All plots were separated from each other by a 1 m buffer area. On May 20, all fertilizers were applied to the plot surfaces and were thoroughly mixed with soil.
Plant and soil sampling and measurements
On September 2, the aboveground biomass of S. koroviniiin one 1 m × 1 m subplots within each plot was collected, and any impurities, dry tissue, and litter were removed. The root material was collected in each subplot within a depth of 0–20 cm. Subsequently, living roots (fine roots with a diameter of <2 mm) were separated from dead roots based on the water content of roots. All root samples were washed with running water. The aboveground biomass and fine roots were oven-dried at 75 °C for 48 h. Next, root samples were ground and evenly mixed for elemental analysis. The total N concentrations (% of dry mass) of the aboveground and fine roots were determined colorimetrically using the Kjeldahl acid-digestion method with an Alpkem Auto Analyzer (Kjeldahl System 1026 distilling unit, Hillerød, Denmark). This method was performed after N in the tissue samples was extracted with sulfuric acid. The total P concentrations (% of dry mass) were determined colorimetrically after wet digestion with H2SO4 + HClO4(Parkinson and Allen 1975).
On September 2, four subsamples of soil (0–20 cm) were randomly collected from each plot using a 2 cm-diameter soil auger. These soil subsamples were thoroughly mixed and combined as a single composite sample. All soil samples were sieved through a 2-mm mesh to remove any roots and impurities. Inorganic N (e.g., ammonium and nitrate nitrogen) in the soil samples was measured with a flow injection autoanalyzer (FIAstar 5000, Foss Tecator, Hillerød, Denmark). The available P concentrations in the soil samples were determined by initially extracting P with 0.5 mol L−1 of NaHCO3 from air-dried soil samples. The extracted P was analyzed via the ammonium molybdate method (Sparks et al. 1996). The concentrations of inorganic N and available P in the soil were determined based on the soil dry weight; soil dry weight was determined after drying the soil at 105 °C for 48 h (Yu et al. 2010).
Statistical analysis
Statistical analyses were performed in SPSS version 16.0 (SPSS Inc., Chicago, IL, USA). Data were tested for normality using the Kolmogorov–Smirnov test and for equality of the error variance using Levene’s test. General linear regression models (y = ax + b) and nonlinear regression models (y = axb) were used to determine (1) the relationship between the N application rate and available soil nutrients, (2) the relationship between N application rates and N as well as P concentrations in both aboveground and fine root (and the relationship between N application rates and N:P ratios), (3) the relationship between available soil nutrients and N as well as P concentrations in both aboveground or fine root (and the relationship soil nutrients and N:P ratios), (4) the relationship between N as well as P concentrations in aboveground (as well as N:P ratios) and those of fine root.
Results
N addition significantly enhanced the soil N availability with available N increasing from 5.74 ± 0.23 to 29.66 ± 1.02 mg kg−1 with increased N input; soil N availability was positively correlated with N addition rates (P < 0.001, R 2 = 0.91). Meanwhile, the available soil P did not change significantly with increasing N input (P = 0.320). Available soil N:P ratios were positively correlated with N addition rates and increased from 1.35 ± 0.11 to 6.03 ± 0.28 with increased N input (P < 0.001, R 2 = 0.90; Fig. 1).

Relationships between N addition rates and soil available nutrient by general linear regression analysis
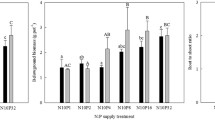
The N concentrations in aboveground were positively correlated with N addition rates (P < 0.001, R 2 = 0.80).In contrast, the P concentrations in aboveground were negatively correlated with N addition rates (P < 0.01, R 2 = 0.40). These results showed that the aboveground N:P concentration ratios were positively correlated with N addition rates (P < 0.001, R 2 = 0.83; Fig. 2). Similarly, the N concentrations and N:P ratios of fine roots were also positively correlated with N addition rates (P < 0.001, R 2 = 0.60; P < 0.001, R 2 = 0.67; respectively). Meanwhile, the P concentrations of fine roots were negatively correlated with N addition rates (P < 0.001, R 2 = 0.44; Fig. 3). Negative relationships were found between N and P concentrations in both the aboveground and fine roots (R 2 = 0.40 and 0.48, respectively; Fig. 4).

Relationships between N addition rates and aboveground N (a), P (b) concentrations and N:P ratios (c) in Seriphidium korovinii by nonlinear regression analysis

Relationships between N addition rates and fine roots N (a), P (b) concentrations and N:P ratios (c) in Seriphidium korovinii by nonlinear regression analysis

Relationships between aboveground (a), or fine roots (b) N and P concentrations in Seriphidium korovinii by general linear regression analysis
The relationships among the N and P concentrations as well as the N:P ratios of aboveground biomass and available soil nutrients were consistent, such that all of the former were positively correlated with the latter (available soil nutrients) (P < 0.001, R 2 = 0.64; P < 0.01, R 2 = 0.29; P < 0.001, R 2 = 0.64; respectively; Fig. 5). The N concentrations and N:P ratios of the fine roots were similar to those of the aboveground biomass (P < 0.001, R 2 = 0.48; P < 0.001, R 2 = 0.39; respectively), whereas the P concentrations of the fine roots did not exhibit significant changes (P = 0. 055; Fig. 6). The stoichiometry of the aboveground biomasswas positively correlated with fine root biomass in S. korovinii(P < 0.001, R 2 = 0.71; P < 0.01, R 2 = 0.85; P < 0.001, R 2 = 0.78; respectively; Fig. 7).

Relationships between soil available nutrient and aboveground N (a), P (b) concentrations and N:P ratios (c) in Seriphidium korovinii by general linear regression analysis

Relationships between soil available nutrient and fine roots N (a), P (b) concentrations and N:P ratios (c) in Seriphidium korovinii by general linear regression analysis

Relationships between aboveground and fine roots in N (a), P (b) concentrations and N:P ratios (c) of Seriphidium korovinii by general linear regression analysis
Discussion
Our results clearly show that short-term N addition affected the stoichiometry of N and P in the aboveground as well as the fine root and affected their relationship for this dominant desert grass species, S. korovinii. In the current study, the N:P ratio in the aboveground and fine root was lower than the critical value reported previously (Güsewell 2004), suggesting this grass species was N-limited. Therefore, short-term N fertilization should be used as a management strategy to restore the degraded desert grasslands of the Kunlun Mountain area. Our results clearly show that for both aboveground and fine roots in plants, N addition positively increased the N concentration while the P concentration decreased. These results suggest that desert plants differ in their stoichiometric characteristics in terms of N and P use. Significant positive relationships were observed between the aboveground and fine root stoichiometry, suggesting that nutrient concentrations and transport to aboveground and fine roots in individual plants are tightly connected. Such knowledge would increase our understanding of the internal N or P use of different tissues and help land managers develop management strategies for desert grasses in response to the nutrient additions caused by climate change or human activities.
The addition of N fertilizer can provide extra available N in the soil pool for plant uptake, as indicated by the increased N concentrations observed in plant tissues after N fertilization (Holub and Tůma 2010; Lü et al. 2013). Previous studies have reported that N concentrations in plant tissues are positively correlated with P concentrations (Geng et al. 2014; Kerkhoff et al. 2006; Liu et al. 2010). In our study, however, N concentrations of both aboveground and fine roots increased with fertilizer N addition, whereas P concentrations in plant decreased. Interestingly, the initial N addition from 0 to 11.2 g resulted in drastically decreased plant P concentration whereas increasing the N fertilization rate resulted in a much smaller decrease in plant P. The addition of N fertilizer thus resulted in a negative relationship between N and P concentrations in both the aboveground and fine roots of S. korovinii. Similar negative relationships between plant N and P concentrations were also reported previously by Menge and Field (2007) and Van Heerwaarden et al. (2003).
N and P are both critical constituents of many molecules in plant tissues, and these constituents are strongly linked to each other. A previous study reported that critical N:P values determine the cutoff for N or P limitation for terrestrial plants that range from 10 to 20 in terrestrial foliage in a study that ignored species-specific adaptations to their environment (Güsewell 2004). Our results indicated that S. korovinii was N-limited rather than P-limited, as indicated by the N:P ratios of 3.31 and 2.19, and these ratios increased after N fertilization for the aboveground and fine roots, respectively. However, Drenovsky and Richards (2004) reported that the critical N:P values obtained from their experiment were inaccurate in a desert ecosystem. Plant N:P ratios are primarily regulated through the adjustment of N and P uptake rates induced by whole-plant signaling mechanisms (Forde 2002; Imsande and Touraine 1994).These results suggested that plants regulated the N and P uptake rates by both positive and negative patterns. Furthermore, N-deficient plants of S. korovinii could enhance the rate of N uptake and reduce the rate of P uptake. Hence, the results of the present study specified that N fertilization caused a larger N pool and smaller P pool of both aboveground biomass and fine roots of S. koroviniiin degraded desert grassland. Therefore, N fertilization should be appropriate when short-term fertilization is used as a management strategy to restore degraded desert grassland in this region.
The nutrient concentrations in leaf tissue were consistently correlated with that in roots, implying that trade-offs exist in managing plant organ economics and stoichiometric regularities of plant organs (Geng et al. 2014; Kerkhoff et al. 2006). Hence, the main focus of research related to N use and limitation should be to understand the role of plant organs (Freschet et al. 2010). We recognized that the relationships between N, P, and N:P ratios with aboveground biomass and fine roots have similar patterns with those relationships in leaves and roots (Geng et al. 2014; Holdaway et al. 2011; Kerkhoff et al. 2006; Liu et al. 2010). Freschet et al. (2010) reported that the nutrient content of root tissues tended to be similar to that in leaves and stems. These researchers further suggested that the N and P concentrations as well as the N:P ratios of the aboveground and fine roots are consistently correlated. Our results provide strong evidence that intraspecific variations in the main element traits (N, P, and N:P ratios) are consistent not only at the level of individual plant organ, but are also consistent at the whole-plant level. The patterns of the relationships between aboveground and fine root maybe caused by the adaptive strategy used by S. korovinii to thrive in areas with relatively infertile soils.
In general, the nutrient concentrations of plant tissues are positively correlated with soil nutrients (Hendricks et al. 2000; Lü et al. 2013; Nadelhoffer 2000). Our results showed that fine root P concentrations were marginally significantly related to available soil nutrients (P = 0. 055; Fig. 6); however, those of aboveground were determined to be significantly related to available soil nutrients. We assumed that N fertilizer alters the original status of nutrient limitation in plants, resulting in conservative P use. Therefore, plants preferentially transported P from the fine roots to the aboveground because of the increased demand for P in plant photosynthetic tissues. In addition, N:P ratios were also regulated by the stoichiometry of plant tissues and directly affected by soil nutrient availability.
Previous studies reported that the geometric means of leaf N and P concentrations were 18.6 and 1.21 mg g−1, respectively, with an N:P ratio of 14.4 (Han et al. 2005). Geng et al. (2014) claimed that the average values of the fine root N and P concentrations of 139 species in northwest China were 11.4 and 0.83 mg g−1, respectively, with an average N:P ratio of 13.73. Meanwhile, the global means of the fine root concentrations of N and P were 11.7 and 1.1 mg g−1, respectively, with an average N:P ratio of 11(Jackson et al. 1997). Our results showed that the N and P concentrations of the aboveground of S. korovinii without N fertilizer input were 9.87 and 2.98 mg g−1, respectively, with an N:P ratio of 3.31. Furthermore, the N and P concentrations of the fine roots of S. korovinii were 4.95 and 2.28 mg g−1, respectively, with an N:P ratio of 2.19. This finding suggested that the N concentrations of the aboveground and fine roots of S. korovinii were much lower than the global average, whereas those of P was much higher than the global or China’s geometric means, resulting in a lower N:P ratio. These results may account for the species-specific adaptive strategy of S. korovinii, which adapts to low N conditions by have lower N requirements or tighter N recycling.
In conclusion, this study demonstrated the conservative nature of the response of stoichiometry in the aboveground and fine roots of N-deficient plants to N addition in an infertile area. N input resulted in a higher N pool. However, the P pool did not change significantly in the soil after 1 year of N addition. Short-term N addition resulted in conservative P use. These results suggest that N fertilizer should be employed when short-term fertilization is used as a management strategy to restore degraded desert grasslands in this region. The N:P ratios were regulated by both positive and negative patterns. Moreover, we determined that plants tend to transport P from the fine root to aboveground for photosynthesis, and the intraspecific variations in the main element traits (N, P, and N:P ratios) were consistent at the whole-plant level.
References
Aerts R, Cornelissen J, Van Logtestijn R, Callaghan T (2007) Climate change has only a minor impact on nutrient resorption parameters in a high-latitude peatland. Oecologia 151:132–139
Drenovsky RE, Richards JH (2004) Critical N:P values: predicting nutrient deficiencies in desert shrublands. Plant Soil 259:59–69
FordeBG 2002 The role of long-distance signalling in plant responses to nitrate and other nutrients. J Exp Bot 53:39–43
Freschet GT, Cornelissen JHC, Van Logtestijn RSP, Aerts R (2010) Evidence of the ‘plant economics spectrum’ in a subarctic flora. J Ecol 98:362–373
Galloway JN, Dentener FJ, Capone DG, Boyer EW, Howarth RW, Seitzinger SP, Asner GP, Cleveland C, Green P, Holland E (2004) Nitrogen cycles: past, present, and future. Biogeochemistry 70:153–226
Geng Y, Wang L, Jin D, Liu H, He J-S (2014) Alpine climate alters the relationships between leaf and root morphological traits but not chemical traits. Oecologia 175:445–455
Gordon WS, Jackson RB (2000) Nutrient concentrations in fine roots. Ecology 81:275–280
Güsewell S (2004) N: P ratios in terrestrial plants: variation and functional significance. New phytol 164:243–266
Han W, Fang J, Guo D, Zhang Y (2005) Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol 168:377–385
Hendricks JJ, Aber JD, Nadelhoffer KJ, Hallett RD (2000) Nitrogen controls on fine root substrate quality in temperate forest ecosystems. Ecosystems 3:57–69
Holdaway RJ, Richardson SJ, Dickie IA, Peltzer DA, Coomes DA (2011) Species-and community-level patterns in fine root traits along a 120 000-year soil chronosequence in temperate rain forest. J Ecol 99:954–963
Holub P, Tůma I (2010) The effect of enhanced nitrogen on aboveground biomass allocation and nutrient resorption in the fern Athyrium distentifolium. Plant Ecol 207:373–380
Imsande J and Touraine B (1994) N demand and the regulation of nitrate uptake. Plant Physiol 105:3
Jackson RB, Mooney H, Schulze E-D (1997) A global budget for fine root biomass, surface area, and nutrient contents. P Natl Acad Sci USA 94:7362–7366
Kerkhoff AJ, Fagan WF, Elser JJ, Enquist BJ (2006) Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants. Am Nat 168:E103–E122
Kozovits A, Bustamante M, Garofalo C, Bucci S, Franco A, Goldstein G, Meinzer F (2007) Nutrient resorption and patterns of litter production and decomposition in a Neotropical Savanna. Funct Ecol 21:1034–1043
Li Y, Song XL, Zhao JN, WangH, BaiL, Yang DL (2015) Responses of plant diversity and primary productivity to nutrient addition in a Stipa baicalensis grassland, China. J Integr Agr 14:2099–2108
Li L, Gao X, Li X, Lin L, Zeng F, Gui D, Lu Y (2016) Nitrogen (N) and phosphorus (P) resorption of two dominant alpine perennial grass species in response to contrasting N and P availability. Environ Exp Bot 127:37–44
Liu G, Freschet GT, Pan X, Cornelissen JH, Li Y, Dong M (2010) Coordinated variation in leaf and root traits across multiple spatial scales in Chinese semi-arid and arid ecosystems. New Phytol 188:543–553
Lü X-T, Kong D-L, Pan Q-M, Simmons ME, Han X-G (2012a) Nitrogen and water availability interact to affect leaf stoichiometry in a semi-arid grassland. Oecologia 168:301–310
Lü XT, Freschet GT, Flynn DF, Han XG (2012b) Plasticity in leaf and stem nutrient resorption proficiency potentially reinforces plant–soil feedbacks and microscale heterogeneity in a semi-arid grassland. J Ecol 100:144–150
Lü XT, Reed S, Yu Q, He NP, Wang ZW, Han XG (2013) Convergent responses of nitrogen and phosphorus resorption to nitrogen inputs in a semiarid grassland. Global Change Biol 19:2775–2784
Menge DNL, Field CB (2007) Simulated global changes alter phosphorus demand in annual grassland. Global Change Biol 13:2582–2591
Nadelhoffer KJ (2000) The potential effects of nitrogen deposition on fine-root production in forest ecosystems. New Phytol 147:131–139
Ostertag R (2001) Effects of nitrogen and phosphorus availability on fine-root dynamics in Hawaiian montane forests. Ecology 82:485–499
Parkinson J, Allen S (1975) A wet oxidation procedure suitable for the determination of nitrogen and mineral nutrients in biological material. Commun Soil Sci Plant 6:1–11
Sparks DL, Page A, Helmke P, Loeppert R, Soltanpour P, Tabatabai M, Johnston C, Sumner M (1996) Methods of soil analysis. Part 3-chemical methods. Soil Science Society of America Inc, Madison
Sterner RW, Elser JJ (2002) Ecological stoichiometry: the biology of elements from molecules to the biosphere. Princeton University, Princeton
Van Heerwaarden L, Toet S, Aerts R (2003) Nitrogen and phosphorus resorption efficiency and proficiency in six sub-arctic bog species after 4 years of nitrogen fertilization. J Ecol 91:1060–1070
Xia J, Wan S (2008) Global response patterns of terrestrial plant species to nitrogen addition. New Phytol 179:428–439
Yu Q, Chen Q, Elser JJ, He N, Wu H, Zhang G, Wu J, Bai Y, Han X (2010) Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability. Ecol Lett 13:1390–1399
Yuan Z, Chen HY, Reich PB (2011) Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus. Natcommun 2:344
Acknowledgements
This study was supported jointly by the West Light Project of the Chinese Academy of Sciences (2015-XBQN-B-21), the China 1000 Talent Program (Y472171), and National Natural Science Foundation of China (No. 31570002, U1603343). We would like to thank Fengli Chen, Jianrong Lu, Gangliang Tang, Zichun Guo, Yang Ma for assistance with field and laboratory work.
Author contributions
LL is responsible for conception and design, analysis, data collection, manuscript write and critical revision of the article and overall responsibility. XG is responsible for critical revision, DG, BL and BZ are responsible for analysis and data collection, XL is responsible for obtained funding.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Li, L., Gao, X., Gui, D. et al. Stoichiometry in aboveground and fine roots of Seriphidium korovinii in desert grassland in response to artificial nitrogen addition. J Plant Res 130, 689–697 (2017). https://doi.org/10.1007/s10265-017-0930-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10265-017-0930-8