
Abstract
A new species of structurally preserved fern rhizome, Ashicaulis plumites (Osmundaceae, Filicales), is described from the Middle Jurassic Tiaojishan Formation in western Liaoning Province, NE China. The new species is characterized by a peculiar sclerenchyma mass in the petiolar vascular bundle concavity. This sclerenchyma mass varies from a linear-shape to a mushroom-like shape with a remarkable outward protuberance, which distinguishes the present new species from other Ashicaulis species. Such a protuberance is very rare among osmundaceous ferns, and should represent a unique type for sclerenchymatous tissue in the osmundaceous vascular bundle concavity. Recognition of the peculiar structure of this new fossil species enriches anatomical diversity of permineralized osmundaceous ferns, indicating that the family Osmundaceae might have experienced a remarkable diversification during the Middle Jurassic in NE China. The new species show anatomical similarities to Osmunda pluma Miller from the Palaeocene of North America. The occurrence of A. plumites in the Middle Jurassic of China provides a new clue for understanding the evolution of some members of the living subgenus Osmunda.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Osmundaceae, a basal family of living filicalean ferns, is regarded as a sister clade to all other leptosporangiate ferns based on molecular data (Pryer et al. 2004; Rai and Graham 2010; Smith et al. 2006). Living Osmundaceae comprises about 21 species of four genera, including Osmunda Linnaeus, Todea Willdenow, Leptopteris Presl as well as Osmundastrum Presl (McAvoy 2011; Metzgar et al. 2008). This family is significant among fern taxa due to its extensive fossil record with more than 200 species (Tidwell and Ash 1994; Tidwell and Clifford 1995). Among them, over 80 species are recognized based on permineralized rhizome specimens (Tian et al. 2008). Compared to fossil foliages, the permineralized materials are crucial for studying the classification, phylogeny and evolution of the Osmundaceae because they bear more anatomical and phylogenetic information (Tian et al. 2008). However, the anatomical diversity and phytogeographic history of the fossil Osmundaceae are not well understood.
The western Liaoning and northern Hebei Provinces of northeastern China hosts one of the most significant Jurassic fossil localities and centers of diversification for osmundaceous rhizomes in the Northern Hemisphere (Tian et al. 2013a). To date, six species of Ashicaulis and two species of Millerocaulis were documented in this region, including A. hebeiensis (Wang) Tidwell, A. liaoningensis (Zhang et Zheng) Tidwell, A. macromedullosus Matsumoto, Saiki, Zhang, Zheng et Wang, A. claytoniites Cheng, A. beipiaoensis Tian, Wang, Zhang, Jiang et Dilcher and A. wangii Tian et Wang as well as M. sinica Cheng et Li and M. preosmunda Cheng, Wang et Li (Cheng and Li 2007; Cheng et al. 2007; Cheng 2011; Matsumoto et al. 2006; Tian et al. 2013a, b; Wang 1983; Zhang and Zheng 1991). In addition, Yang et al. (2010) briefly described a well preserved osmundaceous rhizome with fronds and in situ spores from the Middle Jurassic of Chaoyang, western Liaoning. These records provide important evidences for understanding the palaeogeographical distribution of osmundaceous ferns. In this paper, we describe a new species, i.e., A. plumites sp. nov. from the Middle Jurassic Tiaojishan Formation in western Liaoning, NE China. Compared to other fossil and living osmundaceous ferns, this new species is characterized by a specialized petiolar structure yielding a sclerenchyma mass with a distinctive outward protuberance in the petiolar vascular bundle concavity, which distinguishes the present new species from other Ashicaulis species. Such a protuberance has been only observed in Osmunda pluma Miller from the Palaeocene of North America, and represents a distinct type for sclerenchymatous tissue in the osmundaceous vascular bundle concavity. The specialized structure of this new fossil species enriches anatomical diversity of permineralized osmundaceous ferns, indicating that the family Osmundaceae might have experienced a remarkable diversification during the Middle Jurassic in NE China.
Materials and methods
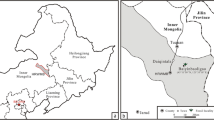
The fossil materials include two structurally preserved rhizome specimens. They were collected from the Middle Jurassic Tiaojishan (=former Lanqi) Formation in Taizishan and Laimaying Villages of Changgao Town, Beipiao City, Liaoning Province, NE China. The Tiaojishan Formation, with a thickness of about 2300 m, mainly consists of intermediate extrusive and pyroclastic rocks with intercalations of basic volcanic and sedimentary rocks (Wang et al. 1989). Over the past several years, a large number of plant fossils with exceptional preservations have been collected and investigated from the Middle Jurassic deposits in the western Liaoning region, NE China, including cycad stems and conifer wood (Jiang et al. 2008, 2012; Zhang et al. 2006). Many leaf fossils have also been reported from this formation, including ferns, cycads, ginkgophytes, conifers, sphenophytes and bryophytes as well (e.g. Wang et al. 2006; Zheng and Zhang 1982; Zhang and Zheng 1987). The Tiaojishan formation is generally considered to be the Middle Jurassic in age (e.g. Duan et al. 2009; Zhang and Zheng 1987; Zhang et al. 2008), though some recent high-precision 40Ar/39Ar data of volcanic rock dating revealed an age of 160.7 ± 0.4 or 158.7 ± 0.6 million years (Ma) (late Middle Jurassic to early Late Jurassic) for the lowest part of the Tiaojishan Formation (Chang et al. 2009).
The specimens described here are permineralized with silica. Thin sections were prepared by standard methods (Jones and Rowe 1999). Specimens described in this paper include two rhizomes with two microscopic slides made from each of the rhizome. All specimens and slides are housed in Paleontological Museum of Liaoning, Shenyang, China, with the registration numbers PMOL–B01252 and PMOL–B01253. Photographs were taken with ACT–1C for DXM1200C Software adapted to Nikon E600 ECLIPSE Microscope, and NLS–Element F 3.0 Software adapted to Nikon SM800 Stereomicroscope.
Systematics
Order––Filicales
Family––Osmundaceae Berchtold et Presl, 1820
Subfamily––Osmundoideae sensu Tidwell, 1994
Genus––Ashicaulis Tidwell 1994
Species––Ashicaulis plumites Tian et Wang sp. nov.

A. plumites sp. nov. (Holotype, PMOL–B01252) a Cross section of the rhizome with a single stem surrounded by petiole bases and roots, scale bar 1.0 cm. b Cross section of the stem, showing the xylem cylinder (XC), leaf trace (LT), inner cortex (IC) and outer cortex (OC), scale bar 1.0 mm. c Enlargement of xylem cylinder (XC), showing an ectophloic–dictyoxylic siphonostele, leaf traces (LT) and root trace (RT) in inner cortex (IC), scale bar 0.4 mm. d Cross section of pith (P), showing parenchymatous cells, scale bar 0.1 mm. e Cross section, showing a leaf trace with one protoxylem cluster (arrow) in inner cortex (IC), scale bar 0.25 mm. f Cross section, showing a leaf race with two protoxylem clusters (arrows) in outer cortex (OC), scale bar 0.25 mm. g Cross section of an adventitious root with diarch xylem, scale bar 0.1 mm

A. plumites sp. nov. (Holotype, PMOL–B01252) a Cross section of a petiole base near point of attachment to the stem, showing homogeneous sclerotic ring (SR), vascular bundle (VB) and small sclerenchymatous masses (arrows) in stipular wing (SW), scale bar 0.4 mm. b, c Petiole base at basal level, showing homogeneous sclerotic ring (SR), C-shaped vascular bundle (VB) and sclerenchyma masses (SM) in stipular wings (SW) and vascular bundle concavity (arrows), scale bar 1.0 mm (b), 0.5 mm (c). d, e Petiole bases at median level, showing heterogeneous sclerotic ring (SR) with a thin layer of thick-walled fibers on the abaxial side (black arrows), a curved clavate sclerenchyma mass (SM) in vascular bundle (VB) concavity and a sclerenchyma mass (SM) in stipular wing (SW), scale bar 0.5 mm. f Petiole base at more diatal level, showing the heterogeneous sclerotic ring (SR) with its abaxial side occupied by a thick-walled fiber arch, and a huge sclerenchyma mass (SM) with outward protuberance in vascular bundle (VB) concavity, scale bar 1.0 mm. g, h Petiole bases at distal end of stipular zone, showing thick-walled fiber arch (arrows) bifurcating into two lateral parts along abaxial side of sclerotic ring (SR), and sclerenchyma mass (SM) with outward protuberance in vascular bundle concavity, scale bar 1.0 mm

A. plumites sp. nov. Sketch drawings based on the paratype (PMOL–B01253), showing serial transverse sections of petiole bases (abaxial side upper). Scale bar 1.0 mm. Notes: Gray colour indicates thin-walled fibers; black colour indicates the thick-walled fibers; the letters indicate different stages of petiole bases

A. plumites sp. nov. (Paratype, PMOL–B01253) a External view of specimen, scale bar 1.0 cm. b Cross section of rhizome, showing pith (P), xylem cylinder (XC), inner cortex (IC) and outer cortex (OC), scale bar 2.0 mm. c Cross section of stem, showing the xylem cylinder (XC), an ectophloic dictyoxylic siphonostele with definite leaf gaps, inner cortex (IC), outer cortex (OC) and leaf traces (LT), scale bar 1.0 mm. d–h Petiole bases at different levels, showing vascular bundle (VB), heterogeneous sclerotic ring (SR) with abaxial sclerenchymatous fiber belt (SFB), stipular wing (SW) and the varieties of sclerenchyma arrangement (SM) in vascular bundle concavity and stipular wings; arrows indicate thick-walled fiber arch on abaxial side of sclerotic ring, scale bar 0.5 mm. i, j Petiole bases above stipular level with a huge mushroom-like sclerenchymatous mass in vascular bundle concavity, and thick-walled fiber arch bifurcated into two lateral parts (arrows), scale bar 1.0 mm (i), 2.0 mm (j)
Species diagnosis
Permineralized rhizome surrounded by petiole bases and adventitious roots. Ectophloic dictyoxylic siphonostele. Pith 1.0–2.2 mm in diameter. Xylem cylinder with about 20 xylem strands, 0.5–0.6 mm in thickness (9–10 tracheids). Xylem segments separated by complete leaf gaps. Cortex two-layered; inner cortex containing about 15 leaf traces; outer cortex with about 13 leaf traces. Leaf trace reniform to C-shaped, adaxially concave, protoxylem cluster single, endarch, first bifurcating in the outer cortex. Near point of attachment of the petiole base to the stem, a small sclerenchymatous mass occurring in the vascular bundle concavity and in each stipular wing. Distally, sclerenchyma tissue in the vascular bundle concavity undergoing a series of shape variation from ovate to crescent, curving clavate and mushroom-like shaped, and finally becoming mushroom-like shaped just above the stipular region. Sclerotic ring heterogeneous, near the basal level of the stipular region, a thin thick-walled fiber arch appearing on the abaxial side; then it completely occupying the whole abaxial side of the sclerotic ring, and bifurcating into two lateral masses near the high level of the stipular region. The sclerenchyma tissue in the stipular wing increasing in size distally, and varies to linear shape near the middle of the stipular region. Adventitious roots diarch.
Holotype: PMOL–B01252.
Paratype: PMOL–B01253.
Repository: All specimens are deposited in the Palaeontological Museum of Liaoning, Shenyang, China.
Type locality: Taizishan and Laimaying Villages of Changgao Town, Beipiao, Liaoning Province, NE China.
Horizon: The Tiaojishan Formation, Middle Jurassic.
Etymology: The specific epithet plumites is proposed to indicate the resemblance of the new species to the fossil taxon O. pluma in anatomical characters, which was reported from the Paleocene Fort Union Formation of North Dakota, USA (Miller 1967).
Description
Two permineralized rhizome specimens, numbered PMOL–B01252 and PMOL–B01253, are designated as the new species. The specimen PMOL–B01252 is selected as the holotype for bearing well-preserved anatomical structures. The paratype, PMOL–B01253 is not as well-preserved as the holotype; however, its petiolar structure is well-preserved which is critical for the classification of Osmundaceae.
The holotype is preserved as a conical to cylindrical stem with a maximum diameter of 6.0 cm (Fig. 1a). A transverse section of the holotype shows only a single stele surrounded by a sheath of petiole bases and adventitious roots (Fig. 1a).
The stem is about 5.5–8.0 mm in diameter, and comprised of a pith, a xylem cylinder and a two-layered cortex (Fig. 1b). The pith is 1.0–2.2 mm across, and composed of homogeneous isodiametric parenchymatous cells (Fig. 1d). The xylem cylinder is somewhat twisted in the holotype, and consists of about 20 xylem strands, each about 0.5–0.6 mm thick (9–10 tracheids) (Fig. 1c). The xylem strands with mesarch protoxylem elements are mainly composed of metaxylem tracheids surrounding the protoxylem points (Fig. 1c). The xylem cylinder is dissected mostly by definite leaf gaps which are debatably immediate type (Fig. 1c). The phloem is ectophloic, but not well preserved.
The cortex contains many helically arranged leaf traces. The whole cortex can be differentiated into two distinct layers, i.e. a 0.85 mm thick inner cortex, and a 1.5–2.5 mm thick outer cortex (Fig. 1b). The parenchymatous inner cortex is poorly preserved, and contains over 15 leaf traces; the outer cortex is sclerenchymatous with 13 leaf traces (Fig. 1b). The leaf trace is reniform, with a single endarch protoxylem cluster when departing from the stele (Fig. 1e), and becomes C-shaped with two protoxylem strands when entering into the outer cortex (Fig. 1f). Upon departing from the stele, the leaf trace enters the petiolar zone.
The petiole base bears two typical stipular wings (Figs. 2, 3, 4d–h). The petiolar size, shape and sclerenchyma arrangement varies at different levels. Immediately proximal to the junction of the stem, the petiole base is characterized by a homogeneous sclerotic ring composed of thin-walled fibers and a C-shaped vascular strand with more than two adaxial protoxylem clusters (Figs. 2a, b, 3); at this level, a very small sclerenchyma mass occurs in each petiolar stipular wing (Figs. 2a, b, 3). Distally, the sclerenchyma tissue in the petiolar vascular bundle concavity undergoes a series of shape variation subsequently from ovate, crescent, curved clavate to a mushroom or umbrella shape (Figs. 2c–h, 3, 4d–j). Just in the stipular region, the petiolar sclerotic ring varies distally from homogenous to heterogeneous; at the basal level of the stipular region, a thin thick-walled fiber arch appears on the abaxial side of the sclerotic ring (Figs. 2d, e, 3, 4d, e). Distally, the thick-walled fiber arch completely occupies the whole abaxial side of the sclerotic ring (Figs. 2f, 3, 4f). Near the high level of the stipular region, the thick-walled fiber arch bifurcates into two individual masses on the lateral part of the abaxial side of the sclerotic ring (Figs. 2g, h, 3, 4g, h). The sclerenchyma tissue in the stipular wing increases in size distally, and becomes linear shape near the middle of the stipular region. No sclerenchyma tissue occurs in other part of the petiolar cortical region. Adventitious roots bear typical diarch xylem (Fig. 1g), and always arise endogenously from the stem xylem cylinder (Fig. 1c).
Comparison
Osmundaceous taxa are characterized by the two-layered cortex, the C-shaped leaf traces, the stipular wings, and the sclerotic ring encircling the vascular strands of the stipular petiole base. (Bower 1926; Hewitson 1962; Miller 1971; Ogura 1972; Tidwell and Ash 1994). Among the two subfamilies of Osmundaceae, the Thamnopteroideae characterizes itself by a protostele, whereas Osmundoideae has a dictyoxylic siphonostele or a dictyostele (Tidwell and Ash 1994). Anatomical characters of the present fossil specimens are consistent with the anatomical features of the Osmundaceae, and particularly with that of the subfamily Osmundoideae. The Osmundoideae includes five fossil genera (Palaeosmunda Gould, Ashicaulis Tidwell, Millerocaulis Erasmus ex Tidwell emend. Tidwell, Osmundacaulis Miller emend. Tidwell and Aurealcaulis Tidwell et Parker) (Tian et al. 2008). The combination of anatomical feature (ectophloic dictyoxylic siphonostele, distinct leaf gaps and oval sclerotic ring) indicates the affinity of the new species to the genus Ashicaulis. Most of the anatomically preserved osmundaceous trunks are not attributable to the living species, and their fronds, sporangia, and spores are almost unknown. These fossils could represent either species of extinct genera or extinct species of genera with living representatives (Cheng 2011; Jud et al. 2008; Rothwell et al. 2002). The new species described here should be taxonomically assigned to Ashicaulis.
According to Tidwell (1994), the genus Ashicaulis was restricted to osmundaceous species that have xylem cylinders bearing numerous definite leaf gaps. Nearly 30 species have been referred to Ashicaulis so far worldwide (Cheng 2011; Tian et al. 2008, 2013a, b). The petiolar sclerenchyma arrangement is considered as one of the key features for classifying osmundaceous species (Hewitson 1962; Miller 1971). Most of the reported Ashicaulis species are characterized by a homogeneous petiolar sclerotic ring, while only five species are with a heterogeneous sclerotic ring as does our present material. These are A. johnstonii (Tidwell, Munzing et Banks) Tidwell, A. kidstonii (Stopes) Tidwell, A. liaoningensis, A. claytoniites and A. wangii (Cheng 2011; Stopes 1921; Tian et al. 2013b; Tidwell 1994; Tidwell et al. 1991). Among these five species, A. johnstonii and A. kidstonii are reported from the Upper Triassic and the Cretaceous of Australia, respectively (Stopes 1921; Tidwell et al. 1991), and other three species are all described from the Middle Jurassic of western Liaoning, NE China. Since the presence of heterogeneous petiolar sclerotic ring, A. plumites is compared to these five species (Table 1).
Compared to the new species, A. kidstonii has two sclerenchyma masses in the petiolar vascular bundle concavity (Miller 1971; Stopes 1921). In A. claytoniites, the whole periphery of the sclerotic ring is encircled by thick-walled fiber (Cheng 2011). In A. johnstonii and A. liaoningensis, the thick-walled fiber arch on the abaxial side of the sclerotic ring never bifurcates into two lateral parts (Tidwell et al. 1991; Zhang and Zheng 1991). The new species bears great similarities to A. wangii in the structure of the petiolar sclerotic ring as well as the sclerenchyma mass in the stipular wing; however, differs from the later by the absence of sclerenchymatous clusters in the petiolar inner cortex (Tian et al. 2013b).
A. plumites is characterized by a unique sclerenchyma mass in the petiolar vascular bundle concavity. This sclerenchyma mass is characterized by a remarkable outward protuberance which distinguishes itself from other five Ashicaulis species. The mushroom or umbrella form of the sclerenchyma is very rare among osmundaceous ferns, and is only documented in O. pluma Miller from the Palaeocene of North Dakota, America (Miller 1971). These two species are also very similar to each other in the type of petiolar sclerotic ring and sclerenchyma mass in the stipular wings. However, many scattered sclerenchymatous clusters occur in the inner and outer petiolar cortex of O. pluma (Miller 1971) which are absent in A. plumites. Based on the above comparisons, the present fossils are designated as a distinct new species of Ashicaulis.
It is noted that the genus Millerocaulis resembles Ashicaulis in the anatomical structures, geological range and palaeo-geographical distribution pattern (Tian et al. 2008). Vera (2008) proposed the recombination of Ashicaulis Tidwell and Millerocaulis Erasmus ex Tidwell, and pointed out that recent specimens referred to Ashicaulis should be compared to previously described species of both Ashicaulis and Millerocaulis. To date, about 11 species of Millerocaulis have been reported worldwide ranging from the Triassic to the Cretaceous (Tidwell 1986; Vera 2010). Among them, six species bear heterogeneous petiolar sclerotic ring, represented by M. preosmunda, M. sinica, M. embreei Stockey et Smith, M. juandahensis Tidwell et Clifford, M. indentata Hill, Forsyth et Green and M. tekelili Vera (Cheng and Li 2007; Cheng et al. 2007; Hill et al. 1989; Stockey and Smith 2000; Tidwell and Clifford 1995; Vera 2010). Though sharing a heterogeneous petiolar sclerotic ring, all these six species are distinct from A. plumites. In M. embreei, M. tekelili and M. indentata, numerous sclerenchyma clusters occur in the petiolar cortex (Stockey and Smith 2000; Vera 2010), while they are absent in A. plumites. In M. juandahensis, two individual sclerenchyma masses exist in the vascular bundle concavity (Tidwell and Clifford 1995). Compared to A. plumites, M. preosmunda and M. sinica bear one large mass and several small sclerenchyma clusters in the stipular wing (Cheng and Li 2007; Cheng et al. 2007). Furthermore, among these six species, there is no outward protuberance in the sclerenchyma mass of the vascular bundle concavity.
Discussion
For osmundaceous ferns, sclerenchymatous tissues are well-developed in several regions of the petiole base (i.e., the vascular bundle concavity, inner and outer petiolar cortex and the sclerotic ring) (Hewitson 1962; Miller 1971). The distribution pattern of the petiolar sclerenchyma is crucial for the taxonomy and phylogeny of osmundaceous ferns (Hewitson 1962; Miller 1967; Tidwell and Ash 1994). As far as the sclerenchymatous tissues in the vascular bundle concavity is concerned, three major types are generally recognized among the genus Ashicaulis, including (1) those lacking sclerenchyma tissues, represented by A. macromedullus, A. websteri (Tidwell, Munzing et Banks) Tidwell, A. Kolbei (Seward) Tidwell, A. beardmorensis (Schopf) Tidwell, A. amajolensis (Sharma) Tidwell, A. sahnii (Vishnu-Mittre) Tidwell (Matsumoto et al. 2006; Schopf 1978; Seward 1907; Sharma 1973; Tidwell 1994; Tidwell et al. 1991; Vishnu-Mittre 1955); (2) those with one crescent or linear shaped sclerenchyma mass, including A. beipiaoensis, A. wangii, A. johnstonii, A. liaoningensis, A. santaecruscis (Herbst) Tidwell, A. wadei (Tidwell et Rushforth) Tidwell. (Herbst 1977; Tian et al. 2013a, b; Tidwell and Rushforth 1970; Tidwell et al. 1991; Zhang and Zheng 1991); (3) those with two sclerenchyma masses, represented by A. gibbiana (Kidston et Gwynne-Vaughan) Tidwell, A. patagonica (Archangelsky et de la Sota) Tidwell, A. wrighti (Tidwell, Munzing et Banks) Tidwell, A. brogani (Tidwell, Munzing et Banks) Tidwell, A. sawanensis (Tidwell, Munzing et Banks) Tidwell, A. australis Vera. (Archangelsky and de la Sota 1962; Herbst 1977; Kidston and Gwynne–Vaughan 1907; Tidwell et al. 1991; Vera 2007).
The new species A. plumites is similar to the above mentioned type 2. However, the sclerenchyma mass of type 2 is always inwardly concave. In contrast, the sclerenchyma mass in A. plumites varies from a linear shape to a mushroom-like shape with remarkable outward protuberance. Therefore, A. plumites should represent a peculiar type of sclerenchymatous tissues in the osmundaceous vascular bundle concavity.
To explain the inducement mechanism of such a peculiar feature, the phyto-physiology of the petiolar sclerenchymatous tissues is noted. Combined with other parenchymatous tissues, these sclerenchymatous tissues play a role in providing a mechanical strength for the fronds. Undoubtedly, the more petiolar sclerenchymatous tissues the higher mechanical strength is provided. Different strategies are performed by osmundaceous plants to get greater mechanical strength. For instance, osmundaceous rhizomes reported from the Southern Hemisphere are mostly with a homogeneous sclerotic ring with thin-walled fiber and numerous well-developed small sclerenchymatous masses in the petiolar inner cortex and stipular wings (e.g. Ashicaulis australis, A. rajmabalensis, A. sahnii, Millerocaulis tekelili, Osmundacaulis janii) (Gupta 1968; Pigg 1993; Tidwell and Vera 2007, 2010; Vishnu-Mittre 1955). In contrast, most Chinese fossils bear a heterogeneous sclerotic ring with thick-walled fibers and a large sclerenchymatous mass in the vascular bundle concavity. As far as the new species described here is concerned, A. plumites develops a specialized sclerenchymatous mass with outward protuberance in the vascular bundle concavity. The distinct strategy to get mechanical strength between the Chinese and the Southern Hemisphere forms may indicate a divergence in character evolution due to geographical isolation. It is of interest that a large desert arid area was distributed in the low latitudes of both hemispheres during the Triassic to Jurassic periods (Boucot et al. 2009). These arid areas might have acted as a natural barrier and geographical isolation for the migration and gene exchange of Ashicaulis.
It is noted, Ashicaulis and Millerocaulis share close similarities in anatomical structures to the living Osmunda than other osmundaceous fossil rhizome taxa (Cheng 2011). They are considered to be the closest ancestors of the extant Osmundaceae (Rothwell et al. 2002; Stockey and Smith 2000; Tidwell and Ash 1994). Of interest, the most taxa of the living genus Osmunda are also characterized by a heterogeneous petiolar sclerotic ring. As mentioned above, A. plumites show similarity to the Palaeocene O. pluma from North America in the petiolar structures. Another species, A. wangii Tian et Wang, recently documented from the Jurassic of the western Liaoning (Tian et al. 2013b), is also structurally similar to O. pluma in the type of the thick-walled fiber arch on the abaxial sclerotic ring and the well-developed sclerenchymatous clusters in the inner petiolar cortex. Generally, O. pluma seems to be a blending outcome of A. plumites and A. wangii.
Miller (1971) gave a remarkable conclusion on the phylogeny of both living and fossil osmundaceous ferns based on anatomical features. In his cladistic analysis, O. claytoniana Linnaeus, O. nathorstii Miller, O. pluma, O. oregonensis (Arnold) Miller, O. regalis Linnaeus, O. japonica Thunberg and O. lancea Thunberg form one of the paraphyletic groups of subgenus Osmunda (Miller 1971). Among these species, O. claytoniana and O. nathorstii form a secondary branch, the rest forms another branch. Miller (1971) emphasized that in O. pluma and O. oregonensis, bifurcation of the petiolar abaxial thick-walled fiber arch occurs just above the stipular region, while in O. regalis, O. japonica, and O. lancea division of the arch occurs in the lower one-third of the stipular region with the two lateral clusters changing almost immediately to form a narrow, inconspicuous band in the abaxial semicircle of the ring which extends upward the remaining length of the stipular region. O. claytoniana is similar to O. regalis, O. japonica and O. lancea, but changes are arrested with the formation of two lateral masses of sclerenchyma which extend upward throughout the remainder of the stipular region (Miller 1967). Totally, in the present new species, bifurcation of the thick-walled fiber arch also occurs in the stipular region and extends upward just as in the O. claytoniana. Considering the remarkable anatomical similarity of A. plumites, O. pluma and O. claytoniana, the new species should also be a member of this paraphyly.
Yatabe et al. (1999) discussed the phylogeny of Osmundaceae based on rbcL nucleotide sequences. In their neighbor-joining tree, O. claytoniana, O. regalis, O. japonica and O. lancea also form a paraphyly (Yatabe et al. 1999). The divergence time between this paraphyly and other members of Osmundaceae was evaluated to be about 210 Ma (Yatabe et al. 1999). The occurrence of A. plumites in the Middle Jurassic of China provides a new clue for understanding the divergence time of this paraphyletic group.
In conclusion, the current report of the new fossil species A. plumites enriches the anatomical diversity of permineralized osmundaceous ferns, and provides further information for understanding the evolution of the osmundaceous ferns.
References
Archangelsky S, de la Sota ER (1962) Estudio anatomico de un estípite petrificado de “Osmundites” de edad Jurássica, procedente del Gran Bajo de San Julián, Provincia de Santa Cruz. Ameghiniana 2:153–164
Boucot AJ, Chen X, Scotese CR, Fan JX (2009) Phanerozoic global climatic reconstruction. Science Press, Beijing (in Chinese)
Bower F (1926) The ferns (Filicales). The Eusporangiatae and other relatively primitive ferns, vol 2. Cambridge University Press, Cambridge
Chang SC, Zhang HC, Renne PR, Fang Y (2009) High-precision 40Ar/39Ar age constraints on the basal Lanqi formation and its implications for the origin of angiosperm plants. Earth Planet Sci Lett 279:212–221
Cheng YM (2011) A new species of Ashicaulis (Osmundaceae) from the Mesozoic of China: a close relative of living Osmunda claytoniana L. Rev Palaeobot Palynol 165:96–102
Cheng YM, Li CS (2007) A new species of Millerocaulis (Osmundaceae, Filicales) from the Middle Jurassic of China. Rev Palaeobot Palynol 144:249–259
Cheng YM, Wang YF, Li CS (2007) A new species of Millerocaulis (Osmundaceae) from the Middle Jurassic of China and its implication for evolution of Osmunda. Int J Plant Sci 168:1351–1358
Duan Y, Zheng SL, Hu DY, Wang LJ, Wang LW (2009) Preliminary report on Middle Jurassic strata and fossils from Linglongta area of Jianchang, Liaoning. Glob Geol 28:143–147
Gupta KM (1968) On a new species of Osmundites, O. rajmahalense sp. nov. from the Rajmahal Hill, Bihar. Proc India Natl Sci Acad 55:428–429
Herbst R (1977) Dos nuevas especies de Osmundacaulis (Osmundaceae, Filices) y otros restos de Osmundales de Argentina. Facena 1:19–44
Hewitson W (1962) Comparative morphology of the Osmundaceae. Ann Missouri Bot Garden 49:57–93
Hill RS, Forsyth SM, Green F (1989) A new genus of osmundaceous stem from the Upper Triassic of Tasmania. Palaeontology 32:287–296
Jiang HE, Ferguson DK, Li CS, Cheng YM (2008) Fossil coniferous wood from the Middle Jurassic of Liaoning Province, China. Rev Palaeobot Palynol 150:37–47
Jiang ZK, Wang YD, Zheng SL, Zhang W, Tian N (2012) Occurrence of Sciadopitys-like fossil wood (Conifer) in the Jurassic of western Liaoning and its evolutionary implications. Chin Sci Bull 57:569–572
Jones TP, Rowe NP (1999) Fossil plants and spores: modern techniques. Geological Society, London
Jud N, Rothwell GW, Stockey RA (2008) Todea from the lower cretaceous of western North America: implications for the phylogeny, systematics, and evolution of modern Osmundaceae. Am J Bot 95:330–339
Kidston R, Gwynne–Vaughan DT (1907) On the fossil Osmundaceae. Pt I Trans R Soc Edinb 45:759–780
Matsumoto M, Saiki K, Zhang W, Zheng SL, Wang YD (2006) A new species of osmundaceous fern rhizome, Ashicaulis macromedullosus sp. nov. from the Middle Jurassic, northern China. Palaeontol Res 10:195–205
McAvoy WA (2011) A new combination in the fern genus Osmundastrum (Osmundaceae). Novon 21:354–356
Metzgar JS, Skog JE, Zimmer EA, Pryer KM (2008) The paraphyly of Osmunda is confirmed by phylogenetic analyses of seven plastid Loci. Syst Bot 33:31–36
Miller CN (1967) Evolution of the fern genus Osmunda. Contrib Mus Paleontol Univ Mich 21:139–203
Miller CN (1971) Evolution of the fern family Osmundaceae based on anatomical studies. Contrib Mus Paleontol Univ Mich 23:105–169
Ogura Y (1972) Comparative anatomy of the vegetative organs of the Pteridophytes. Handbuch der Pflanzenanatomie, Berlín, p 502
Pryer KM, Schuettpelz E, Wolf PG, Schneider H, Smith AR, Cranfill R (2004) Phylogeny and evolution of ferns (Monilophytes) with a focus on the early leptosporangiate divergences. Am J Bot 91:1582–1598
Rai HS, Graham SW (2010) Utility of a large, multigene plastid data set in inferring higher-order relationships in ferns and relatives (monilophytes). Am J Bot 97:1444–1456
Rothwell GW, Taylor EL, Taylor TN (2002) Ashicaulis woolfei nov. sp.: additional evidence for the antiquity of osmundaceous ferns from the Triassic of Antarctica. Am J Bot 89:352–361
Schopf JM (1978) An unusual Osmundaceous specimen from Antarctica. Can J Bot 56:3083–3095
Seward AC (1907) Notes on fossil plants from South Africa. Geol Mag 4:482–487
Sharma BD (1973) Anatomy of Osmundaceous rhizomes collected from the Middle Jurassic of Amarjola in the Rajmahal Hills, India. Palaeontographica B 140:151–160
Smith AR, Pryer KM, Schuettpelz E, Korall P, Schneider H, Wolf PG (2006) A classification for extant ferns. Taxon 55:705–731
Stockey RA, Smith SY (2000) A new species of Millerocaulis (Osmundaceae) from the lower cretaceous of California. Int J Plant Sci 161:159–166
Stopes MC (1921) The missing link in Osmundites. Ann Bot 35:55–61
Tian N, Wang YD, Jiang ZK (2008) Permineralized rhizomes of the Osmundaceae (Filicales): diversity and tempo–spatial distribution pattern. Palaeoworld 17:183–200
Tian N, Wang YD, Zhang W, Jiang ZK, David DL (2013a) Ashicaulis beipiaoensis, a new species of Osmundaceous fern from the Middle Jurassic of Liaoning Province, Northeastern China. Int J Plant Sci 174:328–339
Tian N, Wang YD, Zhang W, Jiang ZK (2013b) A new structurally preserved fern rhizome of Osmundaceae (Filicales) Ashicaulis wangii sp. nov. from the Jurassic of western Liaoning and its significances for paleogeography and evolution. Sci China-Earth Sci (in press)
Tidwell WD (1986) Millerocaulis, a new genus with species formerly in Osmundacaulis Miller (fossils: Osmundaceae). Sida Contrib Bot 11:401–405
Tidwell WD (1994) Ashicaulis, a new genus for some species of Millerocaulis (Osmundaceae). Sida Contrib Bot 16:253–261
Tidwell WD, Munzing GE, Banks MR (1991) Millerocaulis species (Osmundaceae) from Tasmania, Australia. Palaeontographica Abt B 223:91–105
Tidwell WD, Clifford HT (1995) Three new species of Millerocaulis (Osmundaceae) from Queensland, Australia. Austr Syst Bot 8:667–685
Tidwell WD, Ash SR (1994) A review of selected Triassic to early cretaceous ferns. J Plant Res 107:417–442
Tidwell WD, Rushforth SR (1970) Osmundacaulis wadei, a new osmundaceous species from the Morrison formation (Jurassic) of Utah. Bull Torrey Bot Club 97:137–144
Tidwell WD, Pigg KB (1993) New species of Osmundacaulis emend. from Tasmania, Australia. Palaeontographica Abt B 230:141–158
Vera EI (2007) A new species of Ashicaulis Tidwell (Osmundaceae) from Aptian strata of Livingston Island, Antarctica. Cretac Res 28:500–508
Vera EI (2008) Proposal to emend the genus Millerocaulis Erasmus ex Tidwell 1986 to recombine the genera Ashicaulis Tidwell 1994 and Millerocaulis Tidwell emend. Tidwell 1994. Ameghiniana 45:693–698
Vera EI (2010) A new specimen of Millerocaulis (Osmundales: Osmundaceae) from the Cerro Negro Formation (Lower Cretaceous), Antarctica. Rev Mus Argentino Cienc Nat 12:41–46
Vishnu-Mittre (1955) Osmundites sahnii sp. nov., a new species of permineralized osmundaceous rhizomes from India. Palaeobotanist 4:113–118
Wang WL, Zheng SL, Zhang LJ, Pu RG, Zhang WG, Wu HZ, Ju RH, Dong GY, Qi H (1989) Mesozoic stratigraphy and palaeontology of Western Liaoning, vol 1. Geological Publishing House, Beijing (in Chinese with English abstract)
Wang YD, Saiki K, Zhang W, Zheng SL (2006) Biodiversity and palaeo-climate of the Middle Jurassic floras from the Tiaojishan formation in western Liaoning, China. Prog Nat Sci 16:222–230
Wang ZQ (1983) Osmundacaulis hebeiensis, a new species of fossil rhizomes from the Middle Jurassic of China. Rev Palaeobot Palynol 39:87–107
Yatabe Y, Nishida H, Murakami N (1999) Phylogeny of Osmundaceae inferred from rbcL nucleotide sequences and comparison to the fossil evidences. J Plant Res 112:397–404
Yang XJ, Zhang W, Zheng SL (2010) An osmundaceous rhizome with sterile and fertile fronds and in situ spores from the Jurassic of western Liaoning. China Sci Bull 55:3864–3867
Zhang H, Wang MX, Liu XM (2008) Constraints on the upper boundary age of the Tiaojishan Formation volcanic rocks in West Liaoning-North Hebei by LA–ICP–MS dating. China Sci Bull 53:1815–1824 (in Chinese)
Zhang W, Zheng SL (1987) Early Mesozoic fossil plants in western Liaoning, northeast China. In: Yu XH, Wang WL, Liu XT, Zhang W, Zheng SL, Zhang ZC, Yu JS, Ma FZ, Dong GY, Yao PY (eds) Mesozoic stratigraphy and palaeontology of western Liaoning, vol 3. Geological Publishing House, Beijing (in Chinese with English abstract), pp 239–268
Zhang W, Zheng SL (1991) A new species of osmundaceous rhizome from Middle Jurassic of Liaoning, China. Acta Paleontol Sin 30:714–727 (in Chinese with English abstract)
Zhang W, Wang YD, Saiki K, Li N, Zheng SL (2006) A structurally preserved Cycad-like stem, Lioxylon gen. et sp. nov., from the Jurassic in western Liaoning, China. Prog Nat Sci 26:236–248
Zheng SL, Zhang W (1982) New materials of the Middle Jurassic fossil plants from western Liaoning and their stratigraphic significance. Bull Shenyang Inst Geol Min Res Chin Acad Geol Sci 4:160–168 (in Chinese with English abstract)
Acknowledgments
We thank Prof. Shaolin Zheng (Shenyang Institute of Geology and Mineral Resources, Ministry of Land and Resources, Shenyang, China) for his support in field collection and specimen analysis. Special appreciate is due to Prof. R. Herbst (Instituto Superior de Geología Consejo Nacional de Investigaciones Cientificasy Técnicas, Argentina) and Dr. E.I. Vera (División Paleobotánica, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Buenos Aires, Argentina) for providing references. We thank Prof. Harufumi Nishida (Chuo University, Japan) and other two reviewers for their helpful comments and suggestions of the manuscript. This study is jointly supported by the State Key Programme (973 project) of Basic Research of Ministry of Science and Technology, China (2012CB822003); the National Natural Sciences Foundation of China (NSFC 41272010, 41302004 and 40972008), the Team Program of Scientific Innovation and Interdisciplinary Cooperation of CAS; the State Key Laboratory of Palaeobiology and Stratigraphy (Nanjing Institute of Geology and Palaeontology, CAS) (No. 133113); the Science Research Project of Liaoning Provincial Education Department (L2012391) and the Doctoral Fund of Shenyang Normal University.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Tian, N., Wang, YD., Philippe, M. et al. A specialized new species of Ashicaulis (Osmundaceae, Filicales) from the Jurassic of Liaoning, NE China. J Plant Res 127, 209–219 (2014). https://doi.org/10.1007/s10265-013-0611-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10265-013-0611-1