
Abstract
Many legumes form nitrogen-fixing root nodules. An elevation of nitrogen fixation in such legumes would have significant implications for plant growth and biomass production in agriculture. To identify the genetic basis for the regulation of nitrogen fixation, quantitative trait locus (QTL) analysis was conducted with recombinant inbred lines derived from the cross Miyakojima MG-20 × Gifu B-129 in the model legume Lotus japonicus. This population was inoculated with Mesorhizobium loti MAFF303099 and grown for 14 days in pods containing vermiculite. Phenotypic data were collected for acetylene reduction activity (ARA) per plant (ARA/P), ARA per nodule weight (ARA/NW), ARA per nodule number (ARA/NN), NN per plant, NW per plant, stem length (SL), SL without inoculation (SLbac−), shoot dry weight without inoculation (SWbac−), root length without inoculation (RLbac−), and root dry weight (RWbac−), and finally 34 QTLs were identified. ARA/P, ARA/NN, NW, and SL showed strong correlations and QTL co-localization, suggesting that several plant characteristics important for symbiotic nitrogen fixation are controlled by the same locus. QTLs for ARA/P, ARA/NN, NW, and SL, co-localized around marker TM0832 on chromosome 4, were also co-localized with previously reported QTLs for seed mass. This is the first report of QTL analysis for symbiotic nitrogen fixation activity traits.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Many legumes, such as soybean (Glycine max), pea (Pisum sativum), chickpea (Cicer arietinum), and peanut (Arachis hypogaea), are used in the human diet as sources of protein. To maintain the yield of these legumes, enormous amounts of synthetic nitrogen fertilizer are applied to fields each year. In the case of soybean, worldwide production has been estimated to be 220 million tonnes per year, requiring 22 million tonnes of synthetic nitrogen fertilizer (FAO-STAT, http://faostat.fao.org/). This amount of fertilizer represents 27% of the ammonia nitrogen fertilizer produced per year and is equivalent to at least 44 billion m3 of natural gas used as a fuel. In addition, serious environmental problems, such as the loss of excess nitrates and nitrites to groundwater, are caused by the application of synthetic nitrogen fertilizers. To address these problems, there is a worldwide movement toward the enhancement of biological nitrogen fixation and away from dependence on industrial nitrogen fixation.
Leguminous plants form nitrogen-fixing root nodules following reciprocal signal exchange between the plant and rhizobium bacteria (Zuanazzi et al. 1998; Hayashi et al. 2000; Hirsch et al. 2003). Studies of the regulation of nodulation and nitrogen fixation activity have identified mutants involved in the symbiotic relationship between legumes and rhizobia. Although research on the early stages of nodulation has advanced rapidly (Penmetsa and Cook. 1997; Krusell et al. 2002; Nishimura et al. 2002a, b; Oka-Kira et al. 2005; Schnabel et al. 2005; Gonzalez-Rizzo et al. 2006; Murray et al. 2007; Tirichine et al. 2007; Magori et al. 2009), the regulation of nitrogen fixation in the host plant is less well understood. In the model legume plant Lotus japonicus, only four loci related to nitrogen fixation have been identified: ineffective greenish nodules1 (Ign1), stationary endosymbiont nodule1 (Sen1), symbiotic sulfate transporter1 (Sst1), and fail in enlargement of infected cells1 (Fen1) (Suganuma et al. 2003; Krusell et al. 2005; Kumagai et al. 2007; Hakoyama et al. 2009). Mutants at these four loci are all Fix– in phenotype, showing a loss of or defect in nitrogen fixation. The enf1 mutant, which we previously reported, showed elevation of nitrogen fixation activity and enhancement of biomass production without accompanying adverse growth effects (Tominaga et al. 2009), but the gene has not yet been mapped. So far, little of the knowledge of nitrogen fixation genes obtained from studies of model legumes has been applied to the improvement of leguminous crop plants.
Nitrogen fixation activity traits are controlled by multiple quantitative trait loci (QTLs) including 4 genes such as Ign1, Sen1, Sst1, and Fen1 (Suganuma et al. 2003; Krusell et al. 2005; Kumagai et al. 2007; Hakoyama et al. 2009). Although understanding of these QTLs is very important for application to agriculture, there have been no reports of QTL analysis for symbiotic nitrogen fixation activity between legumes and rhizobia. One of the reasons for this lack of information is that most legume crops have features that make genetic analysis difficult, such as large genome size or polyploidy and recalcitrance to transformation. Recently, genetic and genomic information on L. japonicus, including a large-scale genomic DNA sequence, DNA markers, and a high-density genetic linkage map, have accumulated rapidly (Kawaguchi et al. 2001; Gepts et al. 2005; Udvardi et al. 2005; Sato and Tabata 2006; Sato et al. 2001, 2007, 2008); these tools have enabled us to perform QTL analysis in this species. Moreover, considerable material resources such as seeds and DNA clones have been collected and are available through the National BioResource Project in Japan (http://www.legumebase.brc.miyazaki-u.ac.jp/index.jsp).
Here, we carried out a QTL analysis of L. japonicus that considered the following traits related to symbiosis: acetylene reduction activity per plant (ARA/P), ARA per nodule weight (ARA/NW), ARA per nodule number (ARA/NN), NN per plant, NW per plant, stem length (SL), SL without inoculation (SLbac−), shoot dry weight without inoculation (SWbac−), root length without inoculation (RLbac−), and root dry weight (RWbac−).
Materials and methods
Plant materials and growth conditions
A total of 142 and 117 L. japonicus recombinant inbred lines (RILs) were used for mapping under inoculated and non-inoculated growth conditions, respectively. The RILs were derived from a cross between the lines Gifu B-129 (B-129) and Miyakojima MG-20 (MG-20). These lines were self-pollinated to the F8 generation from F2 seeds developed by Dr. M. Kawaguchi (National Institute for Basic Biology) at the Kazusa DNA Research Institute. The seeds were obtained from the National BioResource Project at the Miyazaki University (http://www.legumebase.brc.miyazaki-u.ac.jp/index.jsp) (Gondo et al. 2007).
Seeds of each recombinant inbred or parental line were surface sterilized by immersion in sodium hypochlorite [2% (v/v) containing 0.1% (v/v) Tween 20] for 20 min and rinsed several times with sterile distilled water. After overnight imbibition, the swollen seeds were sown in vermiculite-filled pots that were watered with nitrogen-free B&D medium (Broughton and Dilworth 1971) with or without 1.0 × 107 cells/mL Mesorhizobium loti MAFF303099 (Keele et al. 1969; Kaneko et al. 2000; Saeki and Kouchi. 2000), which had been grown in yeast–mannitol liquid medium (Keele et al. 1969). The plants were grown at 24°C under 16-h-light/8-h-dark conditions at a light intensity of 150 μmol m−2 s−1. In a pilot study, seeds were also sown on 0.8% (w/v) agar medium, and the plates were incubated at 24°C in the dark. After 3 days, the germinated seedlings were transplanted onto B&D medium containing 1.5% (w/v) agar and concomitantly inoculated with 1.0 × 107 cells/mL M. loti per plant (Suzuki et al. 2011). The plants were grown at 24°C under 16-h-light/8-h-dark conditions at a light intensity of 150 μmol m−2 s−1. Six plants out of nine plants grown in vermiculite-filled pots or B&D medium containing 1.5% (w/v) agar were randomly selected, investigated, and experimental values per plant were calculated.
Correlation analysis
The data were used to calculate mean values for each line. Correlations between the ten traits were analyzed by using the Ekuseru-Toukei 2008 software package (Social Survey Research Information Co., Ltd., Tokyo, Japan).
QTL mapping
A total of 96 microsatellite markers chosen from a genetic linkage map of L. japonicus were used for QTL mapping within the LjMG RIL population (Hayashi et al. 2001; Sato et al. 2001). The current marker positions and genotype data for each RI line are available on the website; http://www.kazusa.or.jp/lotus/RIline/index.html.
Interval mapping of each symbiotic trait was carried out with the MapQTL 5.0 package (Van Ooijen 2004) using the multiple QTL method (MQM). Cofactors were identified with the automatic cofactor selection option, and MQM mapping was performed with a step size of 1.0 cM. The logarithmic odds (LOD) significance threshold for each trait was determined for a significance level of 5.0% via a permutation test (1,000 replications; Churchill and Doerge 1994). Moreover, considering the cumulative distribution function of the maximum LOD on a chromosome for QTL analysis based on two QTL genotypes in an RI family (Van Ooijen 1999), the LOD significance threshold was determined to be 2.0 for a 5.0% significance level. The QTL positions were assigned to the positions of maximum local LOD scores.
Acetylene-reduction assay for nitrogenase activity
Nine plants grown in a vermiculite-filled pot watered with B&D medium were inoculated with M. loti. Of these, six randomly selected plants were placed in a 34-mL test tube, which was then covered with a rubber serum cap and degassed for 30 s. Acetylene that was diluted five times was injected into the tube, which was then incubated in a growth chamber at 25°C. After 2 h of incubation, the amount of ethylene formed was determined by gas chromatography (Suzuki et al. 2008; Tominaga et al. 2009).
Results
Symbiotic phenotypes of parent lines
To obtain reliable results in our QTL analysis, we looked for conditions under which we could detect large, reproducible differences in experimental values between MG-20 and B-129. We investigated symbiosis phenotypes (i.e. ARA/P, NN, and NW) of both parental lines grown on B&D agar medium or in vermiculite-filled pots watered with B&D medium for 14, 21, and 28 days after inoculation (DAI).
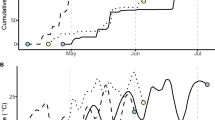
Most experimental values for symbiosis-related phenotypes of plants grown in vermiculite were higher than those of plants grown on agar medium at 14, 21, and 28 DAI (Fig. 1). In the case of plants grown in vermiculite, ARA/P and NW of MG-20 were significantly higher than those of B-129 at all time points (Fig. 1a, b), and NN of MG-20 was significantly higher than that of B-129 at 14 DAI (Fig. 1c). Moreover, ARA/P and NW of B-129 grown in vermiculite were very low at 14 DAI (Fig. 1a, b). The difference in ARA/P between MG-20 and B-129 was the greatest in plants grown in vermiculite and measured at 14 DAI (MG20, 57.1; B-129, 7.5), and the standard deviations for this treatment were small (MG-20, 1.9; B-129, 3.3; P = 1.3 × 10−4; Fig. 1a). Therefore, we determined that 14 DAI was the best time for the analysis of symbiosis-related traits in the RI lines.

Symbiosis-related phenotypes of Lotus japonicus Miyakojima MG-20 (MG-20) and Gifu B-129 (B-129) parent lines. Mesorhizobium loti-inoculated plants were grown for 14, 21, or 28 days in pots containing vermiculite watered with B&D medium (a–c) or grown on B&D agar-solidified (1.5%, w/v) medium (d–f). a, d Acetylene reduction activity per plant. b, e Nodule weight per plant. c, f Nodule number per plant. A total of 18–24 plants were examined in each experiment. Bars represent SD. Statistical significance is indicated by asterisks (*P < 0.05, **P < 0.01, by Student’s t test)
Analysis of phenotypic variation in six symbiosis-related traits and four non-symbiotic traits
Six symbiosis-related traits (ARA/P, ARA/NW, ARA/NN, NN, NW, and SL) were measured in the LjMG RILs and the parental lines at 14 DAI (Table 1). For each trait, the most commonly obtained value among the RILs (indicated by the peak in the histogram) was located between the parental values (Fig. 2). In general, MG-20 grows faster than B-129. To eliminate influence of the differences in growth rate between MG-20 and B-129, we investigated the LjMG RILs and the parental lines at 14 days after sowing (DAS) without inoculation for four non-symbiotic traits (SLbac−, RLbac−, SWbac−, and RWbac−). For each trait except RWbac−, the most commonly obtained value among the RILs (indicated by the peak in the histogram) was located between the parental values; for RWbac−, the B-129 parental value was over the peak of the histogram (Fig. 2). In symbiotic traits, each pair of traits except for NN and ARA/NN showed a significant correlation (Fig. 3). The greatest correlations were those between ARA/P and ARA/NN (r 2 = 0.88) and between ARA/P and NW (r 2 = 0.84). ARA/NW showed negative correlations with the other five traits; all of the other significant correlations were positive. In non-symbiotic traits, each pair of traits except for SLbac− and RLbac−, SLbac− and RWbac− showed a significant positive correlation (Fig. 3). Moreover, SLbac− trait had strong correlation with SL traits (r = 0.73), and also showed significant positive correlation to symbiotic traits except for ARA/NW.

Phenotypic values of Lotus japonicus Miyakojima MG-20 × Gifu B-129 recombinant inbred lines. The mean parental values are indicated by arrows. M Miyakojima MG-20, B Gifu B-129; n = 6 for SLbac−, SWbac−, RLbac−, and RWbac−, n = 30 for SL, n = 48 for ARA/P, ARA/NN, ARA/NW, NW, and NN

Coefficients of correlation between six traits in Lotus japonicus Miyakojima MG-20 × Gifu B-129 recombinant inbred lines. Values shown are significant at P < 0.05. ns, not significant; ARA/P acetylene reduction activity per plant; ARA/NW acetylene reduction activity per nodule weight; ARA/NN acetylene reduction activity per nodule number; NN nodule number per plant; NW nodule weight per plant; SL stem length; SLbac−, stem length without inoculation; RLbac−, root length without inoculation; SWbac−, shoot dry weight without inoculation; RWbac−, root dry weight without inoculation
QTL analysis by MQM
The LOD threshold at the 5% significance level was different for each trait: 3.5 for ARA/P, 3.4 for ARA/NN, 16.7 for ARA/NW, 3.3 for NN, 3.2 for NW, 3.1 for SL, 2.7 for SLbac−, 2.7 for RLbac−, 2.8 for SWbac−, and 2.7 for RWbac−. In addition, considering the cumulative distribution function of the maximum LOD on a chromosome for QTL analysis based on two QTL genotypes in an RI family (Van Ooijen 1999), we determined the LOD significance threshold at 5.0% to be 2.0, so we used 2.0 as the minimum LOD score for QTL detection in this study. QTL positions were assigned at the positions of maximum local LOD scores. A total of 34 significant QTLs were identified for 9 traits, comprising 9 QTLs for nitrogen fixation traits (ARA/P, ARA/NN, and ARA/NW), 6 QTLs for nodulation traits (NN and NW), 10 QTLs for SL, and 9 QTLs for non-symbiotic traits (SLbac−, RLbac−, and SWbac−) (Table 2; Fig. 4).

Molecular linkage map of L. japonicus and the locations for symbiosis-related QTLs. Circles containing arrowheads indicate the positions of QTLs with significant LOD scores: the size of each circle indicates the LOD score, as described in the figure key, and the direction of each arrow indicates the direction of effect of the B-129 allele. Orange or blue circles indicate QTLs determined under growth condition with or without inoculation, respectively
Traits related to nitrogen fixation activity (ARA/P, ARA/NN, ARA/NW)
Four ARA/P QTLs were detected on chromosomes 2, 4 and 5; at each of the four loci, the B-129 allele was associated with lower ARA/P. The QTL on chromosome 2 showed the largest effect, explaining 15.1% of the total variation. Two ARA/NN QTLs were detected on chromosomes 2 and 4; again, the B-129 allele was associated with a lower value for the trait. These QTLs explained 11.1 and 8.5% of the total variation, respectively. Three ARA/NW QTLs were detected on chromosomes 3, 4, and 5. The B-129 QTL alleles on chromosome 4 and 5 were associated with higher ARA/NW and the allele on chromosome 3 was associated with lower ARA/NW. The QTL on chromosome 3 explained 6.9% of the total variation, whereas those on chromosome 4 and 5 explained 10.8 and 13.0% of total variation, respectively. QLTs for ARA/P and ARA/NN were co-located on chromosomes 2 (marker TM0002) and 4 (TM0832), and the additive effects of the B-129 allele were negative for all four QTLs. QTLs for ARA/P and ARA/NW were co-located on chromosomes 4 (TM0030–TM0664) and 5 (TM0095), with additive effects in opposite directions for the two traits (e.g., B-129 alleles were associated with lower ARA/P but higher ARA/NW).
Traits related to nodulation morphology (NN, NW)
A single NN QTL was detected, on chromosome 3. This QTL explained 21.6% of the total variation for this trait, and the B-129 allele led to an increase in NN. Five NW QTLs were detected on chromosomes 2, 3, 4, and 5. The B-129 allele for the QTL on chromosome 3 was associated with higher NW, and the B-129 allele for the QTLs on chromosomes 2, 4, and 5 was associated with lower NW. The QTL on chromosome 4 near TM0832 showed the largest effect among the NW QTLs, explaining 8.9% of the total variation for this trait.
Several of the NW QTLs were co-located with QTLs for nitrogen fixation traits. The four NW QTLs on chromosomes 2, 4, and 5 were each co-located with a QTL for ARA/P and sometimes with a QTL for ARA/NN. Among the QTLs on chromosome 2 (TM0002) and one group on chromosome 4 (TM0832), the B-129 alleles were associated with a reduction in all of the co-located traits; among the other two groups (chromosome 4, TM0030–TM0664, and chromosome 5, TM0095), the B-129 alleles at the co-located QTLs had additive effects in different directions.
Traits related to plant growth (SL)
Because stem length is different between MG-20 and B-129 (Table 1), we investigated it in the RILs at 14 DAI. Ten SL QTLs were detected, with each chromosome having at least one. The B-129 alleles on chromosome 3 and 6 were associated with increased SL; those on chromosomes 1, 2, 4, and 5 were associated with decreased SL. The QTL at TM0113–TM0805 on chromosome 1 showed the largest effect, explaining 13.3% of the total variation in SL. Two of the SL QTLs were co-located with ARA/P and NW QTLs on chromosomes 4 (TM0832) and 5 (TM0095); in each set, the B-129 alleles were associated with lower values for each trait.
Traits related to non-symbiotic plant growth (SLbac−, RLbac−, SWbac−, RWbac−)
In general, MG-20 grows faster than B-129. Therefore, there is a possibility that the differences in symbiotic traits (i.e. ARA/P, ARA/NN, ARA/NW, NW, NN, and SL) observed between MG-20 and B-129 are simply due to faster growth rate in MG-20. To confirm this, we investigated the LjMG RILs and the parental lines at 14 DAS without inoculation; non-symbiotic condition. Six SLbac− QTLs were detected on chromosomes 1, 2, 3, 5, and 6. The B-129 alleles on chromosome 3 and 6 were associated with increased SLbac−; those on chromosomes 1, 2, and 5 were associated with decreased SLbac−. The QTL at TM0027–TM0063 on chromosome 1 showed the largest effect, explaining 16.7% of the total variation in SLbac−. A single RLbac− QTL was detected, on chromosome 5. This QTL explained 8.3% of the total variation for this trait, and the B-129 allele led to a decrease in RLbac−. Two SWbac− QTL were detected on chromosomes 1 and 5; the B-129 allele was associated with a lower value for the trait. These QTLs explained 8.9 and 10.7% of the total variation, respectively. There was no detection of the QTL for RWbac− trait. Four of the SLbac− QTLs were co-located with SL QTLs on chromosomes 1, 2, and 3; in each set, the B-129 alleles at the co-located QTLs had additive effects in the same directions. Two of the SWbac− QTLs were co-located with SL QTLs on chromosomes 1 and 5 with additive effects of the same directions.
Comparison of RILs classified by genotype
For each marker locus, we classified each of the 142 RILs as either “MG-20 type” or “B-129 type” and then calculated the mean value for each trait within each of the two genotype categories. We used Student’s t test to identify significant differences in trait values between the two genotypes at each locus.
Figure 5 shows the results of comparisons for QTLs on chromosomes 2, 4, and 5, which include co-localized QTLs. For DNA markers linked to QTLs identified in this study, the differences in mean experimental values between the MG-20-type and B-129-type RILs were larger than for other DNA markers on the same chromosome (Fig. 5). Furthermore, the level of significance was higher for differences at QTL-linked markers than for other markers. In addition, even though no SL QTL had been detected near TM0002 on chromosome 2, the difference in values between MG-20-type and B-129-type RILs for this marker was large and statistically significant (Fig. 5).

Comparison of L. japonicus RILs classified by genotype. The 142 RILs were divided into “MG20-type” and “B-129-type” on the basis of genotype data at each DNA marker on a chromosome 2, b chromosome 4, and c chromosome 5. The line graphs show the average of experimental values for the RILs of each genotype at each locus. Bars indicate significant differences calculated by student t test. The relevant co-located QTL markers are enclosed with the square
Discussion
We sought to identify QTLs for symbiotic nitrogen fixation activity traits in the leguminous model plant L. japonicus. A total of 34 QTLs were identified for ten traits examined, and many of these QTLs were co-located.
QTL studies of agronomic traits in legumes under greenhouse conditions show that environmental factors strongly influence QTL expression (Julier et al. 2007; Gondo et al. 2007). Because nodule development and nitrogen fixation activity are greatly influenced by the soil condition and other environmental conditions, we conducted a pilot study to find growth conditions under which we could detect large, stable differences in experimental values between parents MG-20 and B-129. Since the differences in experimental values of ARA/P, NW, and NN between MG-20 and B-129 were the largest and the standard deviation was the smallest when plants were grown in vermiculite and measured at 14 DAI (Fig. 1), we judged that these conditions were the most suitable for this QTL analysis.
Gondo et al. (2007) investigated PH (plant height) QTLs in the same L. japonicus RIL population as used here and identified QTLs at TM0996–TM0190 on chromosome 3 and at TM0756–TM0336 on chromosome 6. These QTLs were co-located with SL QTLs (Fig. 4; Table 2), but the additive effects of the B-129 alleles were in opposite directions. These differences in the direction of additive effects might have been caused by different growth conditions and measurement definitions in the two studies: PH was defined by Gondo et al. (2007) as plant height from the ground to the top of the branching plant at flowering, whereas SL was defined here as the stem length from the ground to the top of the non-branching plant during the vegetative stage. In any case, the DNA region around markers TM0996 and TM0756 appears to regulate plant height at both the vegetative and flowering stages.
There have been reports of QTL co-localization in crops such as rice (Xiong et al. 1999), maize (Khavkin and Coe. 1997), and soybean (Zhang et al. 2004). Here, we found groups of three or more QTLs co-located at TM0002 on chromosome 2 (ARA/P, ARA/NN, and NW), at TM0832 on chromosome 4 (ARA/P, ARA/NN, NW, and SL), at TM0030 on chromosome 4 (ARA/P, ARA/NW, and NW), and at TM0095 on chromosome 5 (ARA/P, ARA/NW, NW, and SL), with additive effects all in the same direction except for ARA/NW (Fig. 4; Table 2). The correlations between the six traits were consistent with these results: there were significant positive correlations between ARA/P, ARA/NN, NW, NN, and SL (except for NN with ARA/NN), and significant negative correlations between ARA/NW and the five other traits (Fig. 3). Furthermore, the differences in phenotypic values between the MG-20-type and B-129-type RILs were the largest for the DNA markers linked to sets of co-located QTLs (TM0002, TM0832, TM0030, and TM0095), and these differences were highly significant (Fig. 5). These results suggest that plant height is up-regulated in part by the same alleles due to the increased nitrogen fixation activity caused by the gain of nodule weight.
Four of the SLbac− QTLs were co-located with SL QTLs on chromosomes 1, 2, and 3 with additive effects in same directions. This result indicates that these QTLs were caused by the differences of growth rate between MG-20 and B-129. On the other hand, symbiotic QTLs (i.e. ARA/P, ARA/NN, ARA/NW, NW and NN) were not co-located with SLbac− QTLs. These results strongly suggested that symbiotic QTLs were distinguishable from non-symbiotic QTLs. Moreover, confidence interval (CI) of the ARA/P, ARA/NW, NW, and SL QTLs near TM0095 on chromosome 5 overlapped with that of SWbac−, but not with that of SLbac− and RLbac−. Therefore, QTLs on chromosome 5 are caused by symbiosis, but may be affected by the differences of growth rate between MG-20 and B-129 weakly. Furthermore, SL QTLs that were not co-located with SLbac− or symbiotic QTLs on chromosome 4 (TM0266–TM0097), on chromosome 5 (TM0186–TM0341), and on chromosome 6 (TM0014, TM0139–TM0756) may be not caused by the symbiosis and the difference of growth rate between parents. In conclusion, symbiotic QTLs have detected on chromosome 2 (TM0550), on chromosome 3 (TM0083, TM0203), on chromosome 4 (TM0832, TM0030–TM0664), and chromosome 5 (TM0095). These results strongly suggest that the QTL analysis at early stage of symbiosis (14 DAI) is effective to detect significant symbiotic QTLs.
There are only four reports of symbiotic nitrogen fixation genes in L. japonicus: ineffective greenish nodules1 (Ign1), stationary endosymbiont nodule1 (Sen1), symbiotic sulfate transporter1 (Sst1), and fail in enlargement of infected cells1 (Fen1) (Suganuma et al. 2003; Krusell et al. 2005; Kumagai et al. 2007; Hakoyama et al. 2009). Interestingly, the confidence interval (CI) of ARA/P, ARA/NN, NW, and SL QTLs at TM0832 on chromosome 4 overlapped with that for the Sen1 locus (8.4–8.8 cM) (Suganuma et al. 2003; Sandal et al. 2006). Furthermore, the QTLs of ARA/P, ARA/NN, NW, and SL that are co-located near TM0832 on chromosome 4 were found in the same location as the QTLs Seed Mass-1 (linked to TM0832; CI 8.6–12.6 cM) and Seed Mass-2 (linked to TM0832; CI 6.6–15.6 cM) investigated by Klein and Grusak (2009). (Seed mass was defined by Klein and Grusak (2009) as the total seed mass per pot divided by the number of seeds per pot). The seed mass QTLs were mapped in the same L. japonicus RIL population as used here, and for all of these co-located QTLs, the B-129 alleles led to reduced values for the trait (Fig. 4). It is usually said that 70–80% of the nitrogen in soybean accumulates during the seed filling period (Hanway and Weber. 1971), so the nitrogen fixation activity during this period greatly influences yield. Since the ARA/P, ARA/NN, NW, and SL QTLs identified at 14 DAI were co-localized with seed mass QTLs, we speculate that an allele near TM0832 on chromosome 4 influences plant growth and yield derived from regulation of nitrogen fixation activity at all growth stages from the vegetative growth period to the reproductive growth period. Hence, the difference in DNA sequence around the TM0832 marker between MG-20 and B-129 appears to be an important target for molecular breeding.
Moreover, the CI of the ARA/P, ARA/NW, NW, and SL QTLs near TM0095 on chromosome 5 overlapped with that of the Sym7 locus (16.9–22.3 cM). The sym7 mutant is Hist− (mutants have partially developed nodules) (Schauser et al. 1998; Sandal et al. 2006). Therefore, it appears that a locus near TM0095 on chromosome 5 regulates nitrogen fixation activity and plant growth derived from regulation of nodule development, so this locus may be an additional target for the improvement of legume crops.
Genetic studies in agronomically important legume crops have been limited by large genome size, polyploidy, and difficulty of transformation. To solve this problem, model plants have often been used to identify agronomic traits of interest, to create comparative maps (Choi et al. 2004; Cannon et al. 2006), and to perform synteny analysis (Choi et al. 2004; Zhu et al. 2005). Here, a QTL for NN was identified at TM0083 on chromosome 3, with an increase in NN associated with the B-129 allele. This is the first report of a QTL affecting nodule number in the model legume L. japonicus; until now, the only report of an NN QTL had been in pea (Pisum sativum) (Bourion et al. 2010). Although comparative analysis has revealed conservation between a region of chromosome 3 of L. japonicus and chromosomes 3 and 5 of pea (Zhu et al. 2005), the details of the corresponding genomic regions in each plant have not been reported. In the future, advances in pea genome research will allow us to conduct synteny analysis of the NN QTLs between L. japonicus and pea.
Because the full genome sequence of soybean is now available (Schmutz et al. 2010), the application of L. japonicus QTLs to soybean is expected to advance rapidly. In rice, improvement in lodging resistance and productivity has been achieved by QTL analysis combined with positional cloning (Ookawa et al. 2010). The results of our experiment identified important symbiosis-related genomic regions, including alleles around TM0832 and TM0095, and clearly showed that QTL analysis of symbiotic traits during vegetative growth periods can provide valuable information for molecular breeding. Our future efforts will be focused on the application of symbiotic QTLs discovered in L. japonicus to the study and improvement of legume crops.
References
Bourion V, Rizvi SM, Fournier S, de Larambergue H, Galmiche F, Marget P, Duc G, Burstin J (2010) Genetic dissection of nitrogen nutrition in pea through a QTL approach of root, nodule, and shoot variability. Theor Appl Genet 121:71–86
Broughton WJ, Dilworth MJ (1971) Control of leghaemoglobin synthesis in snake beans. Biochem J 125:1075–1080
Cannon SB, Sterck L, Rombauts S, Sato S, Cheung F, Gouzy J et al (2006) Legume genome evolution viewed through the Medicago truncatula and Lotus japonicus genomes. Proc Natl Acad Sci USA 103:14959–14964
Choi HK, Mun JH, Kim DJ, Zhu H, Baek JM, Mudge J, Roe B, Ellis THN, Doyle J, Kiss GB et al (2004) Estimating genome conservation between crop and model legume species. Proc Natl Acad Sci USA 101:15289–15294
Churchill GA, Doerge RW (1994) Empirical threshold values for quantitative trait mapping. Genetics 138:963–971
Gepts P, Beavis WD, Brummer EC, Shoemaker RC, Stalker HT, Weeden NF, Young ND (2005) Legumes as a model plant family. Genomics for food and feed report of the cross-legume advances through genomics conference. Plant Physiol 137:1228–1235
Gondo T, Sato S, Okumura K, Tabata S, Akashi R, Isobe S (2007) Quantitative trait locus analysis of multiple agronomic traits in the model legume Lotus japonicus. Genome 50:627–637
Gonzalez-Rizzo S, Crespi M, Frugier F (2006) The Medicago truncatula CRE1 cytokinin receptor regulates lateral root development and early symbiotic interaction with Sinorhizobium meliloti. Plant Cell 18:2680–2693
Hakoyama T, Niimi K, Watanabe H, Tabata R, Matsubara J, Sato S et al (2009) Host plant genome overcomes the lack of a bacterial gene for symbiotic nitrogen fixation. Nature 462:514–517
Hanway JJ, Weber CR (1971) Accumulation of N, P, and K by soybean (Glycine max (L.) Merrill) plants. Agron J 63:406–408
Hayashi M, Imaizumi-Anraku H, Akao S, Kawaguchi M (2000) Nodule organogenesis in Lotus japonicus. J Plant Res 112:489–495
Hayashi M, Miyahara A, Sato S, Kato T, Yoshikawa M, Taketa M et al (2001) Construction of a genetic linkage map of the model legume Lotus japonicus using an intraspecific F2 population. DNA Res 8:301–310
Hirsch AM, Bauer WD, Bird DM, Cullimore J, Tyler B, Yoder JI (2003) Molecular signals and receptors: controlling rhizosphere interactions between plants and other organisms. Ecology 84:858–868
Julier B, Huguet T, Chardon F, Ayadi R, Pierre JB, Prosperi JM, Barre P, Huyghe C (2007) Identification of quantitative trait loci influencing aerial morphogenesis in the model legume Medicago truncatula. Theor Appl Genet 114:1391–1406
Kaneko T, Nakamura Y, Sato S, Asamizu E, Kato T, Sasamoto S, Watanabe A, Idesawa K, Ishikawa A, Kawashima K et al (2000) Complete genome structure of the nitrogen-fixing symbiotic bacterium Mesorhizobium loti. DNA Res 7:331–338
Kawaguchi M, Motomura T, Imaizumi-Anraku H, Akao S, Kawasaki S (2001) Providing the basis for genomics in Lotus japonicus: the accessions Miyakojima and Gifu are appropriate crossing partners for genetic analyses. Mol Genet Genomics 266:157–166
Keele BB, Hamilton PB, Elkan GH (1969) Glucose catabolism in Rhizobium japonicum. J Bacteriol 97:1184–1191
Khavkin E, Coe E (1997) Mapped genetic locations for developmental functions and QTLs reflect concerted groups in maize (Zea mays L.). Theor Appl Genet 95:343–352
Klein MA, Grusak MA (2009) Identification of nutrient, physical seed trait QTL in the model legume Lotus japonicus. Genome 52:677–691
Krusell L, Madsen LH, Sato S, Aubert G, Genua A, Szczyglowski K, Duc G, Kaneko T, Tabata S, de Bruijn F et al (2002) Shoot control of root development and nodulation is mediated by a receptor-like kinase. Nature 420:422–426
Krusell L, Krause K, Ott T, Desbrosses G, Kramer U, Sato S, Nakamura Y, Tabata S, James EK, Sandal N et al (2005) The sulfate transporter SST1 is crucial for symbiotic nitrogen fixation in Lotus japonicus root nodules. Plant Cell 17:1625–1636
Kumagai H, Hakoyama T, Umehara Y, Sato S, Kaneko T, Tabata S, Kouchi H (2007) A novel ankyrin-repeat membrane protein, IGN1, is required for persistence of nitrogen-fixing symbiosis in root nodules of Lotus japonicus. Plant Physiol 143:1293–1305
Magori S, Oka-Kira E, Shibata S, Umehara Y, Kouchi H, Hase Y, Tanaka A, Sato S, Tabata S, Kawaguchi M (2009) TOO MUCH LOVE, a root regulator associated with the long-distance control of nodulation in Lotus japonicus. Mol Plant Microbe Interact 22:259–268
Murray JD, Karas BJ, Sato S, Tabata S, Amyot L, Szczyglowski K (2007) A cytokinin perception mutant colonized by Rhizobium in the absence of nodule organogenesis. Science 315:101–104
Nishimura R, Hayashi M, Wu GJ, Kouchi H, Imaizumi-Anraku H, Murakami Y, Kawasaki S, Akao S, Ohmori M, Nagasawa M et al (2002a) HAR1 mediates systemic regulation of symbiotic organ development. Nature 420:426–429
Nishimura R, Ohmori M, Kawaguchi M (2002b) The novel symbiotic phenotype of enhanced-nodulating mutant of Lotus japonicus: astray mutant is an early nodulating mutant with wider nodulation zone. Plant Cell Physiol 43:853–859
Oka-Kira E, Tateno K, Miura K, Haga T, Hayashi M, Harada K, Sato S, Tabata S, Shikazono N, Tanaka A et al (2005) klavier (klv), a novel hypernodulation mutant of Lotus japonicus affected in vascular tissue organization and floral induction. Plant J 44:505–515
Ookawa T, Hobo T, Yano M, Murata K, Ando T, Miura H, Ochiai Y et al (2010) New approach for rice improvement using a pleiotropic QTL gene for lodging resistance and yield. Nat Commun 1:132. doi:10.1038/ncomms1132
Penmetsa RV, Cook DR (1997) A legume ethylene-insensitive mutant hyperinfected by its rhizobial symbiont. Science 275:527–530
Saeki K, Kouchi H (2000) The lotus symbiont, Mesorhizobium loti: molecular genetic techniques and application. J Plant Res 113:457–465
Sandal N, Petersen TR, Murray J et al (2006) Genetics of symbiosis in Lotus japonicus: recombinant inbred lines, comparative genetic maps, and map position of 35 symbiotic loci. Mol Plant Microbe Interact 19:80–91
Sato S, Tabata S (2006) Lotus japonicus as a platform for legume research. Curr Opin Plant Biol 9:128–132
Sato S, Kaneko T, Nakamura Y, Asamizu E, Kato T, Tabata S (2001) Structural analysis of a Lotus japonicus genome. I. Sequence features and mapping of fifty-six TAC clones which cover the 5.4 Mb regions of the genome. DNA Res 8:311–318
Sato S, Nakamura Y, Asamizu E, Isobe S, Tabata S (2007) Genome sequencing and genome resources in model legumes. Plant Physiol 144:588–593
Sato S, Nakamura Y, Kaneko T, Asamizu E, Kato T, Nakao M et al (2008) Genome structure of the legume, Lotus japonicus. DNA Res 15:227–239
Schauser L, Handberg K, Sandal N, Stiller J, Thykjær T, Pajuelo E, Nielsen A, Stougaard J (1998) Symbiotic mutants deficient in nodule establishment identified after T-DNA transformation of Lotus japonicus. Mol Gen Genet 259:414–423
Schmutz J, Cannon SB, Schlueter J, Ma J, Mitros T, Nelson W et al (2010) Genome sequence of the palaeopolyploid soybean. Nature 463:178–183
Schnabel E, Journet EP, de Carvalho-Niebel F, Duc G, Frugoli J (2005) The Medicago truncatula SUNN gene encodes a CLV1-like leucine-rich repeat receptor kinase that regulates nodule number and root length. Plant Mol Biol 58:809–822
Suganuma N, Nakamura Y, Yamamoto M, Ohta T, Koiwa H, Akao S, Kawaguchi M (2003) The Lotus japonicus Sen1 gene controls rhizobial differentiation into nitrogen-fixing bacteroids in nodules. Mol Genet Genomics 269:312–320
Suzuki A, Yamashita K, Ishihara M, Nakahara K, Abe M, Kucho K, Uchiumi T, Higashi S, Arima S (2008) Enhanced symbiotic nitrogen fixation by Lotus japonicus containing an antisense β-1,3-glucanase gene. Plant Biotechnol 25:357–360
Suzuki A, Suriyagoda L, Shigeyama T, Tominaga A, Sasaki M et al (2011) Lotus japonicus nodulation is photomorphogenetically controlled by sensing the R/FR ratio through JA signaling. Proc Natl Acad Sci USA 108:16837–16842
Tirichine L, Sandal N, Madsen LH, Radutoiu S, Albrektsen AS, Sato S, Asamizu E, Tabata S, Stougaard J (2007) A gain-of-function mutation in a cytokinin receptor triggers spontaneous root nodule organogenesis. Science 315:104–107
Tominaga A, Nagata M, Futsuki K, Abe H, Uchiumi T, Abe M et al (2009) Enhanced nodulation and nitrogen fixation in the abscisic acid low-sensitive mutant enhanced nitrogen fixation1 of Lotus japonicus. Plant Physiol 151:1965–1976
Udvardi MK, Tabata S, Parniske M, Stougaard J (2005) Lotus japonicus: legume research in the fast lane. Trends Plant Sci 10:222–228
Van Ooijen JW (1999) LOD significance thresholds for QTL analysis in experimental populations of diploid species. Heredity 83:613–624
Van Ooijen BV (2004) MapQTL1 5, software for the mapping of quantitative trait loci in experimental populations. Kyazma BV, Wageningen
Xiong LZ, Liu KD, Dai XK, Xu CG, Zhang Q (1999) Identification of genetic factors controlling domestication-related traits of rice using an F2 population of a cross between Oryza sativa and O. rufipogon. Theor Appl Genet 98:243–251
Zhang WK, Wang YJ, Luo GZ, Zhang JS, He CY, Wu XL, Gai JY, Chen SY (2004) QTL mapping often agronomic traits on the soybean (Glycine max L. Merr.) genetic map and their association with EST markers. Theor Appl Genet 108:1131–1139
Zhu H, Choi H-K, Cook DR, Shoemaker RC (2005) Bridging model and crop legumes through comparative genomics. Plant Physiol 137:1189–1196
Zuanazzi JA, Clergeot PH, Quirion JC, Husson HP, Kondorosi A, Ratet P (1998) Production of Sinorhizobium meliloti nod gene activator and repressor flavonoids from Medicago sativa roots. Mol Plant Microbe Interact 11:784–794
Acknowledgments
F8 seeds of L. japonicus recombinant inbred lines (RILs) provided by the National Bio-resource Project of the Ministry of Education, Culture, Sports, Science and Technology, Japan. We are grateful to L. Suriyagoda for critical reading of the manuscript. This work was supported by a Grant-in-Aid for Scientific Research (B) from the Japan Society for the Promotion of Science (Grant no. 21380016 to A.S.), the Takano Life Science Research Foundation (Grant to A.S.), and the Sumitomo Foundation (Grant to A.S.), and the Foundation for Research Fellowships of Japan Society for the Promotion of Science for Young Scientists (DC2) (Grant no. 23-3498 to A.T.).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Tominaga, A., Gondo, T., Akashi, R. et al. Quantitative trait locus analysis of symbiotic nitrogen fixation activity in the model legume Lotus japonicus . J Plant Res 125, 395–406 (2012). https://doi.org/10.1007/s10265-011-0459-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10265-011-0459-1