
Abstract
Chloroplast photorelocation movement in green plants is generally mediated by blue light. However, in cryptogam plants, including ferns, mosses, and algae, both red light and blue light are effective. Although the photoreceptors required for this phenomenon have been identified, the mechanisms underlying this movement response are not yet known. In order to analyze this response in more detail, chloroplast movement was induced in dark-adapted Adiantum capillus-veneris gametophyte cells by partial cell irradiation with a microbeam of red and/or blue light. In each case, chloroplasts were found to move toward the microbeam-irradiated area. A second microbeam was also applied to the cell at a separate location before the chloroplasts had reached the destination of the first microbeam. Under these conditions, chloroplasts were found to change their direction of movement without turning and move toward the second microbeam-irradiated area after a lag time of a few minutes. These findings indicate that chloroplasts can move in any direction and do not exhibit a polarity for chloroplast accumulation movement. This phenomenon was analyzed in detail in Adiantum and subsequently confirmed in Arabidopsis thaliana palisade cells. Interestingly, the lag time for direction change toward the second microbeam in Adiantum was longer in the red light than in the blue light. However, the reason for this discrepancy is not yet understood.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Chloroplast photorelocation movement is a well-known phenomenon that has been studied since the 19th century (reviewed by Haupt 1999). Under weak light, chloroplasts move toward a light source in order to maximize their light absorption for efficient photosynthesis (called the “accumulation response”), but they move away from strong light in order to avoid photodamage (called the “avoidance response”) (Kasahara et al. 2002). These phenomena have been analyzed extensively from a physiological point of view (Haupt 1999; Wada et al. 2003; Suetsugu and Wada 2007), but the molecular mechanisms underlying chloroplast movement have not been studied until recently.
The photoreceptors responsible were identified in Arabidopsis thaliana (L.) Heynh. through the isolation of mutants altered in chloroplast movement. Phototropin 2 (phot2) mediates the avoidance response (Jarillo et al. 2001; Kagawa et al. 2001), whereas both phototropin 1 (phot1) and phot2 mediate the accumulation response (Sakai et al. 2001). Photoreceptors for chloroplast movement were also identified in the moss Physcomitrella patens (Hedw.) Brush and Schimp. subsp. patens (Kasahara et al. 2004) and in the fern Adiantum capillus-veneris L. (Kagawa et al. 2004; Kawai et al. 2003; Tsuboi et al. 2007). A chimeric photoreceptor neochrome 1 (neo1, formerly phytochrome3; Nozue et al. 1998), comprising a phototropin fused to a phytochrome chromophore-binding domain at the N-terminus, was identified as a red/far-red and blue light receptor that mediates red as well as blue light-induced chloroplast movement in A. capillus-veneris (Kawai et al. 2003). In P. patens, besides phototropins, phytochromes also participate in red light-induced chloroplast movement (Kadota et al. 2000; Kasahara et al. 2004; Mittmann et al. 2004; Uenaka and Kadota 2007).
Although the photoreceptors for chloroplast movement have been identified in most plant groups (Suetsugu and Wada 2007), the signal transduction pathways are not yet known. The mechanisms of movement have been especially obscure even though experimental results using cytoskeletal inhibitors have suggested the involvement of an actomyosin-dependent system (Suetsugu and Wada 2007). Interestingly, a microtubule system is used in the chloroplast movement of Physcomitrella patens under both red and blue light, but an actomyosin system is used only under blue light (Sato et al. 2001). In this study, we performed a detailed analysis of chloroplast behavior during movement mainly in the two-dimensional gametophyte cells of Adiantum by using a partial cell irradiation technique (Wada and Furuya 1978; Kagawa and Wada 1994, 1999). Our analysis was also extended to A. thaliana palisade cells. In each case, we found that chloroplasts were capable of moving in any direction without turning, suggesting that chloroplasts exhibit no polarity for movement.
Materials and methods
Plant material
Sterilized spores of Adiantum capillus-veneris were sown on the surface of White’s medium (Kagawa and Wada 1996), solidified with 0.5% agar in a Petri dish (6 cm in diameter), and cultured under continuous white light (20–30 μmol m−2 s−1) for 2–3 weeks until two-dimensional prothallia were obtained. The heart-shaped gametophytes were transferred into darkness and incubated for 24–26 h to allow chloroplasts along the periclinal walls to move toward the anticlinal walls (a phenomenon called “dark positioning”) (Tsuboi et al. 2007). Gametophytes that contained some chloroplasts still attached to the periclinal walls were used for the following experiments (Fig. 2).
For Arabidopsis thaliana, seeds were sown on a culture medium with 1/3 Murashige and Skoog inorganic salt and 1% sucrose, solidified with 0.8% agar. After treatment at 4°C for 2 days, plants were grown under a cycle of 16 h white light at approximately 100 μmol m−2 s−1 and 8 h dark at 23°C in an incubator (Biotron LH300-RPSMP; Nippon Medical and Chemical) for 2 weeks and then transferred onto soil and cultured under about 60 μmol m−2 s−1 white light (16 h)/dark (8 h) cycle at 23°C for at least 1 week. Chloroplast movement was observed as described elsewhere (Kagawa and Wada 2002).
Microbeam irradiation
Dark-adapted gametophytes with a few chloroplasts still remaining along the periclinal wall of each cell were transferred into a custom-made cuvette (25 mm in diameter, 5 mm in height) under dim green safe-light conditions (Kadota et al. 1985). The cuvette was made from two steel rings and two round glasses with a silicon rubber-ring spacer in between (Wada et al. 1983). The gametophytes were placed on a microbeam irradiator under dark conditions (see Tsuboi et al. 2006). Chloroplast positioning was observed using infrared light obtained by infrared filter (IR85; Hoya, Tokyo) on a monitor screen attached to a camera that is sensitive to infrared light (C2400-07ER; Hamamatsu Photonics, Hamamatsu, Japan).
A small round area (10 μm in diameter) located near a chloroplast and under the periclinal wall of the gametophyte was irradiated with either a microbeam of red light (166 μmol m−2 s−1) with a transmission peak at 660 nm obtained through an interference filter (half band-width, 34 nm; IF-BPF-660, Vacuum Optics Co. of Japan, Tokyo) or with a microbeam of blue light (113 μmol m−2 s−1) with a transmission peak at 453 nm obtained through an interference filter (half band-width, 31 nm; IF-BPF-453, Vacuum Optics Co. of Japan, Tokyo) and a blue plastic film (#63; Ryuden-sha, Tokyo). Far-red light (8.9 μmol m−2 s−1 and 305 μmol m−2 s−1) was obtained through an FR filter (IF-BPF-740, Vacuum Optics Co. of Japan, Tokyo) with a transmission peak at 740 nm (half band-width, 36 nm). The light sources of blue and red microbeams and far-red and infrared light were obtained from the same halogen lamp (Focusline 12 V-100 W HAL, Philips Lighting, Netherland).
The microbeam and its position could be observed under infrared light at a higher fluence rate than the beam used to observe the gametophytes. Therefore, placement of the red or blue microbeam in relation to a target chloroplast could be analyzed without any interference from visible light. After establishing the irradiation site with the infrared microbeam, the microbeam light was changed from infrared to either red or blue light when needed. For sequential irradiation with two microbeams, the second microbeam of blue or red light with the identical size and fluence rate was presented for 1 min on another part of the cell before the chloroplast reached the first microbeam-irradiated area.
Analysis of chloroplast movement
The path of chloroplast movement during the accumulation response induced by a 1-min exposure to the blue or red microbeam was traced every 15 s using the photographs taken at 15-s intervals under infrared light conditions. The direction of the long axis of the chloroplast at the beginning (just after the start of movement, see the inset of Fig. 2b) and at the end of movement (just before the cessation of movement) was determined every minute from the photographs taken at 15-s intervals. The images obtained were processed and analyzed with “ImageJ” software version 1.33 (shared with http://rsbweb.nih.gov/ij/).
The photographs of chloroplast autofluorescence within Adiantum gametophyte cells were taken every 5 min using fluorescence microscopy (Axioskop; Carl Zeiss, Germany) fitted with a digital camera (DFC340FX; Leica Microsystems, Germany) in order to detect the intrachloroplast positional change of grana stacks during chloroplast movement.
Results
We first examined whether chloroplasts moved along the plasma membrane by either rolling or by sliding (and thereby always attaching a fixed side of chloroplast onto the plasma membrane). Photographs of a pattern of grana stacks of a moving chloroplast were taken from the plasma membrane side every 5 min for 20 min. A track of the moving chloroplast was traced (Fig. 1a, inset) and the distribution patterns of grana stacks in each photograph were compared (Fig. 1b–f). During the 20-min period of observation, the chloroplast moved more than 10 μm. However, the distribution and pattern of grana stacks did not change as was shown by the constant distances measured among some of the grana stacks (Fig. 1g). These findings indicate that chloroplasts do not roll during movement, but slide along the plasma membrane keeping one side in contact with the plasma membrane. Based on these findings, we subsequently analyzed chloroplast movement two-dimensionally rather than monitoring three-dimensional changes, including rolling.

Fluorescence images show the distribution pattern of granas in a chloroplast during its accumulation response induced by whole-cell irradiation with red light (4.5 μmol m−2 s−1) of a dark-adapted gametophyte. Note that the distribution pattern of many granas seen in the chloroplast does not change during the movement. a Outlines of a chloroplast and the path of the center of the chloroplast (inset) during the movement. Intrachloroplast spots distinguishable during the movement are indicated with arrows (1, 2, and 3). b–f Fluorescence photographs of the chloroplast (arrow head) taken every 5 min for 20 min. g Distances among the distinct spots 1–2, 1–3, and 2–3 obtained in each photograph from b–f are plotted. Scale bar = 5 μm
When a blue microbeam was irradiated as a small, round spot (10 μm in diameter) for 1 min near a chloroplast located along the periclinal wall (Fig. 2a, a2), the chloroplast moved toward the irradiated area soon after, or even before, the microbeam was switched off and eventually reached the previously irradiated area (Fig. 2a, a3; see the supplemental movie M1 and Fig. 2b). In this case, we show one representative example, but all results were reproducible in other samples. Figure 2b shows the path of the center of a chloroplast moving toward the area irradiated with the blue microbeam and the directional change of the long axis of the chloroplast (inset) during the 32-min period of movement. The chloroplast began to move toward the microbeam-irradiated area within 1 min after microbeam irradiation, but no apparent directional change of the long axis of the chloroplast occurred during this period of movement. The angle made at the intersection of an imaginary line running through the long axis of the chloroplast with an imaginary line that connected the center of the microbeam to the center of the chloroplast was measured every minute, and the data are shown in Fig. 2c. The angle was maintained at about 80 ± 20° during chloroplast movement. These results indicate that chloroplasts initiate movement to any direction without turning, suggesting that chloroplasts have no polarity for directional movement.

a Photographs showing the chloroplast movement induced by a blue microbeam. a1 A dark-adapted fern gametophyte cell (before microbeam irradiation) with a few chloroplasts still attached to the periclinal wall. A chloroplast at the center of the cell (arrow) was continually observed under infrared light. a2 Chloroplast position (arrow) at the start of a 1-min irradiation with the blue microbeam (large arrowhead). a3 The chloroplast (arrow) reached the beam-irradiated area (arrowhead), as viewed with a beam of infrared light, 29 min after microbeam irradiation. For more details, see the supplemental movie, M1. Photographs were taken every 15 s under infrared light. All data for Fig. 2b–d were obtained from these photographs. Scale bar = 10 μm. b The path of the center of the chloroplast and the directional change of the long axis of the chloroplast (inset) during the movement induced by a 1-min exposure to the blue microbeam. The chloroplast stayed in the original position (arrows) for a few minutes after the onset of microbeam irradiation and then moved toward the beam-irradiated area. The movement path data were obtained every 15 s, and the directional change data (inset) were obtained every minute. Scale bar = 5 μm. c The direction change of the long axis of a chloroplast during the movement induced by 1 min of irradiation by the blue microbeam indicated by the gray bar. Chloroplast direction is shown as the angle formed by an imaginary line of the long axis of the chloroplast and an imaginary line connecting the center of the chloroplast at the original position to the center of the microbeam-irradiated area (see inset). The chloroplast direction was maintained around 80°. d The speed of chloroplast movement toward the microbeam of blue light is shown. Chloroplast movement was induced by 1 min of irradiation by the blue microbeam shown as a gray bar. After a few minutes of lag time, the chloroplasts started to move toward the microbeam-irradiated area at about 0.29 μm min−1. Data were obtained every 15 s
The speed of movement induced by the blue microbeam was measured (Fig. 2d). The representative chloroplast shown in Fig. 2d started to move toward the microbeam-irradiated area after a lag time of about 4 min. Once the chloroplast started to move, the speed of movement was about 0.29 μm min−1 until the chloroplast reached the irradiated area. The mean speed was about 0.28 ± 0.02 μm min−1 under blue microbeam (n = 45) and about 0.32 ± 0.02 μm min−1 under red microbeam (n = 27) (Table 1) with no significant difference between the two speeds (p > 0.05).
Since the results shown in Fig. 2b and c were obtained by one representative example, the relationship between the direction of the chloroplast before microbeam irradiation and the direction of the chloroplast after the onset of movement was studied using 32 different samples under the blue microbeam and 27 different samples under the red microbeam (Fig. 3). If chloroplasts have a polarity (i.e., a head and a tail) for moving along their long or short axes and have to change direction before movement, the data should theoretically fall along the dotted or dashed lines shown in Fig. 3, respectively. However, this was found not to be the case. Most data are located around 0° on the ordinate axis (Fig. 3a). Even when chloroplasts had almost reached the microbeam-irradiated area, their directions were not substantially different from those measured before movement (Fig. 3b). Moreover, even if they turned, the angles were less than 90° towards either plus or minus, again indicating that chloroplasts do not have polarity for movement.

Shown is the relationship between the original direction of the chloroplast and chloroplast rotation just after the onset of movement (a) or just before the cessation of movement near the microbeam (b). The abscissa shows the angle between the long axis of the chloroplast and an imaginary line connecting the center of the chloroplast to the center of the microbeam (shown as θ in the inset) before microbeam irradiation. The ordinate shows the rotated angle of the chloroplasts calculated as the difference in the angle of the chloroplast before and after the onset of movement. The circle drawn with the white dotted line (inset) shows the outline of a chloroplast. Solid circles represent data obtained from chloroplasts irradiated for 1 min by the blue light (n = 32), and open circles represent chloroplasts irradiated for 1 min by the red light (n = 27). The dotted lines indicate an ideal line along which data points should fall if chloroplasts have a polarity of movement parallel to the long axis of the chloroplast and have to rotate before directional movement occurs. The dashed lines indicate the case when polarity is parallel to the short axis of the chloroplast
Next, we analyzed chloroplast movement under conditions in which two microbeams were presented sequentially and at different locations. When a blue microbeam irradiated a small, round spot (10 μm in diameter) for 1 min near a chloroplast located along the periclinal wall (Fig. 4a, a2), the chloroplast moved towards the microbeam-irradiated area. Before the chloroplast reached the irradiated area, a second blue microbeam of identical size and fluence rate was presented for 1 min at another location near the chloroplast (Fig. 4a, a3). After a lag time of a few minutes, the chloroplast moved toward the newly irradiated area without turning (Fig. 4a, a4; see supplemental movie, M2). The movement path and speed of the chloroplasts during this directional change were analyzed in detail (Fig. 4b–d). Figure 4b shows the path of the center of a chloroplast that is moving toward the microbeams of blue light and the direction of the long axis of the chloroplast (Fig. 4b, inset). The directional change in angle of the long axis of the chloroplast during the movement is shown in Fig. 4c. The direction of the long axis of the chloroplast was maintained at about 160 ± 20° during the movement toward the first and second microbeams. The speed of chloroplast movement toward the first blue microbeam-irradiated area was about 0.20 mm min−1, and the speed toward the second blue microbeam-irradiated area was about 0.18 mm min−1 (Fig. 4d).

a Photographs showing the chloroplast movement induced by two sequential blue microbeam irradiations. a1 A dark-adapted fern gametophyte cell (before microbeam irradiation) with a few chloroplasts still attached along the periclinal wall. The chloroplast at the center of the cell (arrow) was continually observed under infrared light. a2 Onset of the first 1-min irradiation with the blue microbeam (arrowhead). a3 The second 1-min irradiation with a blue microbeam with the same fluence rate and diameter as the first beam. Before the chloroplast reached the first irradiated area, the second microbeam was presented at a different location in the same cell (arrowhead). This photograph was taken 18 min after the first beam irradiation. a4 The chloroplast moved toward the second microbeam-irradiated area (arrowhead). This photograph was taken 20 min after the second microbeam irradiation. The white squiggling lines in panels a3 and a4 show the movement paths of the chloroplast. For more details, see a movie of this figure submitted as supplemental movie M2. Photographs were taken every 15 s under infrared light. All data for Fig. 4b–d were obtained from these photographs. Scale bar = 10 μm. b Shown is the path of the center of a chloroplast and the directional change (inset) of the long axis of the chloroplast during the movement induced by 1-min presentations of the first (solid line) and the second (dotted line) blue microbeams. Data were recorded every 15 s or every minute (inset). Scale bar = 5 μm. c The direction change of the long axis of a chloroplast during the movement induced by 1-min presentations each of the first and the second blue microbeams (gray bars). Chloroplast direction is shown as the angle of the long axis of the chloroplast formed by imaginary lines connecting the center of the chloroplast at the original position and the first microbeam-irradiated area. The chloroplast direction was maintained at about 160° during both phases of microbeam irradiation. These data were obtained at 1-min intervals. d The speed of chloroplast movement toward the first and second blue microbeam after each 1-min irradiation (gray bars) is shown. Note that no differences were found between the speeds of movement induced by the first and the second microbeams. Data were recorded every 15 s
To determine the overall trend of the results obtained using one representative chloroplast in Fig. 4b, c, it was tested whether or not chloroplasts turn when they switch direction and move toward the second blue (n = 32) or red microbeam (n = 14) (Fig. 5). The abscissa in Fig. 5 shows the angle between two imaginary lines connecting the center of the chloroplast to the centers of the first and the second microbeams (θ: see inset) at the time of irradiation by the second microbeam. The ordinate shows the angle that the chloroplast turned, i.e., the difference between the direction of the long axis of the chloroplast just before irradiation by the second microbeam and the direction immediately after the chloroplasts started to move toward the second microbeam. Irrespective of the wavelength of the microbeam used, most data points were located around 0° on the ordinate, indicating that chloroplasts changed their moving direction without turning or with only a small turn towards a random direction, if any. Again, these results indicate that chloroplasts can move to any direction without having to turn, suggesting that they have no polarity for directional movement.

The relationship between the degree of direction change of chloroplast movement and the degree of chloroplast rotation before and after the second microbeam irradiation. Namely, the relation between the angle made by two imaginary lines connecting the chloroplast with the first and the second microbeams, indicated by θ in the inset (corresponding to changes in the direction of chloroplast movement induced by the second microbeam) and the direction change of a long axis of chloroplast just before irradiation by the second microbeam and immediately after the chloroplast started to move towards the second microbeam is shown. After the first microbeam irradiation, before chloroplasts reached the first microbeam-irradiated area, the second microbeam was given at various parts of the cell to change the direction of chloroplast movement with various angles shown by θ. Closed diamonds represent the data obtained from chloroplasts irradiated for 1 min by the first and the second blue light (n = 32), and open diamonds represent chloroplasts irradiated for 1 min by the first and the second red light (n = 14). Other details are the same as Fig. 3
Regarding the speed of chloroplast movement, although a lag time was needed for chloroplasts to change their direction and move toward the second microbeam, once the chloroplasts started moving toward the second beam, the speed of movement was not so different for the different light conditions, i.e., about 0.25 ± 0.01 μm min−1 towards the second blue microbeam (n = 26) and about 0.33 ± 0.03 μm min−1 towards the second red microbeam (n = 25) (Table 1). The apparent difference in speeds initiated by the two beams was not statistically significant (p > 0.05).
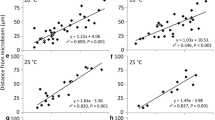
The lag time needed for the chloroplasts moving towards the first blue microbeam to change direction toward the second blue microbeam was 2–4 min (Fig. 6b). Yet, the lag time for chloroplasts moving towards the first red microbeam to change direction toward the second red microbeam was 6–12 min (Fig. 6b). By contrast, the lag time needed for stationary chloroplasts to start moving towards the first red or blue microbeam was 2–4 min (Fig. 6a). The difference between the short and long lag times was statistically significant (p < 0.05). When the first microbeam was red light, the chloroplast took a longer lag time to change direction towards the second microbeam compared to when the first microbeam was blue light, irrespective of the color of the second microbeam (Fig. 6b).

The relationship between the distances from each microbeam to the chloroplasts and lag times before stationary chloroplasts start to move towards the first microbeam (a) and the lag times before the chloroplasts moving towards the first microbeam change direction to move toward the second microbeam (b and c). a The lag times for the first microbeam on average were about 3.1 min under red (n = 44) (open circles: 1R) and about 2.7 min under blue microbeam (n = 48) (closed circles: 1B). b The lag times for the second microbeams. When the first microbeam was red (open symbols), the lag time was long, about 6.2 min (n = 21). However, when the first one was blue (n = 48) (closed symbols), the lag time was short, about 2.8 min, irrespective of the color of the second beam [triangles: second blue microbeam (2B), or squares: second red microbeam (2R)]. c Far-red light effects on the long lag times. After red light irradiation for the first microbeam, far-red light of 8.9 μmol m−2 s−1 (open symbols; wFR, n = 12) or 305 μmol m−2 s−1 (closed symbols; sFR, n = 11) was given just before the second blue microbeam irradiation (circles; 2B) or second red microbeam (diamonds; 2R)
The response to the red light involves neo1, a chimeric photoreceptor between phytochrome and phototropin (Kawai et al. 2003). Therefore, prior to presentation of the second microbeam, cells were irradiated with far-red light (8.9 or 305 μmol m−2 s−1; n = 12 or 11, respectively) for 1 min in order to photoconvert Pfr back to Pr and negate neo1 photoactivation. However, the lag time to respond to the second red or the second blue microbeam was identical in the presence or absence of a far-red light with a fluence rate strong enough to convert Pfr to Pr (Fig. 6b, c).
Finally, we performed similar experiments using Arabidopsis thaliana palisade cells to determine whether the mode of chloroplast movement in Adiantum was conserved in Arabidopsis. Indeed, we found that Arabidopsis chloroplasts showed very similar behavior to those of Adiantum for moving and changing their direction (Fig. 7 and supplemental movie M3).

a Photographs showing chloroplast movement in an Arabidopsis palisade cell (shown with a dotted line) induced by sequential irradiation of two blue microbeams. A cell shown with small arrowheads is an epidermal cell over the palisade cell. a1 A dark-adapted palisade cell with a few chloroplasts still remaining along the periclinal wall is shown. The chloroplast at the center of the cell (arrow) was continually observed and photographed under red light. a2 Onset of continuous irradiation with a blue microbeam (large arrowhead) (113 μmol m−2 s−1). a3 The positions of the chloroplast and the second blue microbeam (large arrowhead) with the same fluence rate and diameter as the first beam. Before the chloroplast reached the first microbeam, the second microbeam was presented at a different location in the same cell. This photograph was taken 8 min after the first beam irradiation. a4 The chloroplast (arrow) moved toward the second microbeam-irradiated area. This photograph was taken 8 min after the second beam irradiation. For more details, see a movie of this figure submitted as supplemental data M3. White squiggling lines in panels a3 and a4 show the movement path of the chloroplast. All data for Figs. 7b–d were obtained from these photographs. Scale bar = 10 μm. b Path of the center of the chloroplast and the directional change (inset) of the long axis of the chloroplast during the movement induced by the first (solid line) and the second (dotted line) blue microbeams. Data were obtained every 15 s or every minute (inset). Scale bar = 5 μm. c The direction change of the long axis of the chloroplast during the movement induced by continuous irradiation of the first and the second blue microbeams started at the timings shown with arrows. Chloroplast direction is shown as the angle of the long axis of the chloroplast formed by imaginary lines connecting the center of the chloroplast at the original position and the first microbeam-irradiated area. The chloroplast direction was maintained around 180° before and after the moving direction change induced by the second microbeam irradiation. These data were obtained at 1-min intervals. d The speed of chloroplast movement toward the first and the second blue microbeam. The timings of microbeam irradiation are shown with arrows. Data were obtained every 15 s
Discussion
This work was performed using two-dimensional gametophyte cells of the fern Adiantum in which chloroplast movement is not only clearly observed but also easily manipulated by using microbeam irradiation. Arabidopsis palisade cells were also used, but presented more difficulties, not only because of their small size, but primarily because the palisade cells are located under epidermal cells, which prevents the microbeam from being sharply focused and impedes the observation of chloroplast movement. In addition, fern gametophyte cells are very sensitive to light. Thus, ferns represent the best plant material for partial cell irradiation experiments with a microbeam (Wada 2007, 2008).
In this study, chloroplast movement in fern gametophytes was induced by microbeam irradiation and chloroplasts were found to move in any direction without turning irrespective of the wavelength of light used, either under red or blue light, i.e., they can move back and forth, even to the side, without turning (Figs. 2b, c; 3; 4b, c; 5). These findings suggest that chloroplasts have no polarity, at least for directional movement. Previous research suggested that chloroplasts may use preexisting actin filaments for their movement (Wada et al. 2003), but it is difficult to understand how chloroplasts would use preexisting cytoskeletons to move in any direction with the same speed (Figs. 2d, 4d, and Table 1) or to change the moving direction within a few minutes of lag time (Fig. 6).
It is thought that plant organelle movement is dependent on the actomyosin system (Wada and Suetsugu 2004). In the Adiantum gametophyte, myosin inhibitors, N-ethylmaleimide (NEM) and 2,3-butanedione monoxime (BDM), inhibited light-induced and mechanical stress-induced chloroplast movement (Kadota and Wada 1992; Sato et al. 1999), as is the case for other plant species (Tlalka and Gabryś 1993; Malec et al. 1996; Paves and Truve 2007), suggesting that chloroplast movement in Adiantum may be also dependent on the actomyosin system. However, comprehensive studies of myosin function in tobacco and in Arabidopsis strongly argue that myosins mediate chloroplast photorelocation movement, because T-DNA knockout, RNAi knockdown, and dominant negative suppression of certain myosin genes clearly affect the movement of mitochondria, peroxisomes, and Golgi bodies but cause no effect on chloroplast photorelocation movement (Avisar et al. 2008; Peremyslov et al. 2008). Therefore, given that both Arabidopsis and Adiantum show very similar behavior of chloroplast movement (Fig. 7), chloroplasts may generally move in any direction by utilizing actin filaments independent of myosins.
The lag time needed to change the direction of movement toward the second microbeam differed for red and blue light treatments (Fig. 6) and may depend on the photoreceptors mediating the response, although far-red light-dependency was not found in our study. The photoreceptors involved in chloroplast photorelocation movement in Adiantum were found to be neo1 for red light-induced movement (Kawai et al. 2003) and phot2 for the avoidance response under strong blue light (Kagawa et al. 2004). The photoreceptors required for the blue light-induced accumulation response may be phot1, phot2, and neo1, but this possibility has not been clarified because a phot1 mutant is not available in Adiantum. The lag time between microbeam irradiation and the start of chloroplast movement toward the microbeam was studied in relation to the distance between the chloroplasts and the microbeam using dark-adapted Adiantum gametophytes (Kagawa and Wada 1996). These authors found that there was almost no lag time under blue microbeam irradiation, but the lag time under red microbeam irradiation was dependent on the distance between the chloroplast and the microbeam-irradiated area. This finding suggests that the speed of signal transfer under blue light was faster than that under red light. The experimental results by Kagawa and Wada (1996) are in accordance with those obtained in this study. One possible explanation for this discrepancy is that neo1 alone is activated during red microbeam irradiation, but all of phot1, phot2, and neo1 are activated during blue microbeam irradiation. Since both the phototropins and neo1 may share the same signal and signal transduction pathways for chloroplast movement, the intensity of the signal released from these three different kinds of photoreceptors at the blue microbeam-irradiated area would be several times greater than the intensity at the red microbeam-irradiated area. However, the signal may persist for longer after the red light is turned off compared to that occurring after the blue light is turned, since the first red light but not blue light delayed moving to the second microbeam (either red or blue light). The Pfr form of phytochrome (i.e., the phytochrome part of neo1) reportedly has a long life time (Kagawa et al. 1994; Yatsuhashi and Kobayashi 1993). Thus, the involvement of neo1 Pfr may explain the long lag time needed to respond to the second microbeam. Red light generates a higher Pfr-to-Pr ratio than blue light and hence more Pfr would remain after the red light is turned off compared to the blue light.
Together, the findings of the present study show that chloroplasts can move in any direction without rotation, which means that they do not have any polarity for movement. However, the mechanisms for this movement are not known. Precise observation of actin filaments located around moving chloroplasts using transgenic plants that express GFP-talin or GFP-fimbrin for visualization of actin filament structures will be necessary to gain further insights into the movement mechanics of chloroplasts.
References
Avisar D, Prokhnevsky AI, Makarova KS, Koonin EV, Dolja VV (2008) Myosin XI-K is required for rapid trafficking of Golgi stacks, peroxisomes, and mitochondria in leaf cells of Nicotiana benthamiana. Plant Physiol 146:1098–1108
Haupt W (1999) Chloroplast movement: from phenomenology to molecular biology. Progress Bot 60:3–36
Jarillo JA, Gabryś H, Capel J, Alonso JM, Ecker JR, Cashmore AR (2001) Phototropin-related NPL1 controls chloroplast relocation induced by blue light. Nature 410:952–954
Kadota A, Wada M (1992) Photoorientation of chloroplasts in protonemal cells of the fern Adiantum as analyzed by use of a video-tracking system. Bot Mag Tokyo 105:265–279
Kadota A, Wada M, Furuya M (1985) Phytochrome-mediated polarotropism of Adiantum capillus-veneris L. protonemata as analyzed by microbeam irradiation with polarized light. Planta 165:30–36
Kadota A, Sato Y, Wada M (2000) Intracellular chloroplast photorelocation in the moss Physcomitrella patens is mediated by phytochrome as well as by a blue-light receptor. Planta 210:932–937
Kagawa T, Wada M (1994) Brief irradiation with red or blue light induces orientational movement of chloroplasts in dark-adapted prothallial cells of the fern Adiantum. J Plant Res 107:389–398
Kagawa T, Wada M (1996) Phytochrome and blue light-absorbing pigment-mediated directional movement of chloroplasts in dark-adapted prothallial cells of fern Adiantum as analyzed by microbeam irradiation. Planta 198:488–493
Kagawa T, Wada M (1999) Chloroplast avoidance response induced by high-fluence blue light in prothallial cells of the fern Adiantum capillus-veneris as analyzed by microbeam irradiation. Plant Physiol 119:917–923
Kagawa T, Wada M (2002) Blue light-induced chloroplast relocation. Plant Cell Physiol 43:367–371
Kagawa T, Kadota A, Wada M (1994) Phytochrome-mediated photoorientation of chloroplasts in protonemal cells of the fern Adiantum can be induced by a brief irradiation with red light. Plant Cell Physiol 35:371–377
Kagawa T, Sakai T, Suetsugu N, Oikawa K, Ishiguro S, Kato T, Tabata S, Okada K, Wada M (2001) Arabidopsis NPL1: a phototropin homologue controlling the chloroplast high-light avoidance response. Science 291:2138–2141
Kagawa T, Kasahara M, Abe T, Yoshida S, Wada M (2004) Function analysis of Acphot2 using mutants deficient in blue light-induced chloroplast avoidance movement of the fern Adiantum capillus-veneris L. Plant Cell Physiol 45:416–426
Kasahara M, Kagawa T, Oikawa K, Suetsugu N, Miyao M, Wada M (2002) Chloroplast avoidance movement reduces photodamage in plants. Nature 420:829–832
Kasahara M, Kagawa T, Sato Y, Kiyosue T, Wada M (2004) Phototropins mediate blue and red light-induced chloroplast movements in Physcomitrella patens. Plant Physiol 135:1388–1397
Kawai H, Kanegae T, Christensen S, Kiyosue T, Sato Y, Imaizumi T, Kadota A, Wada M (2003) Responses of ferns to red light are mediated by an unconventional photoreceptor. Nature 421:287–290
Malec P, Rinaldi RA, Gabryś H (1996) Light-induced chloroplast movements in Lemna trisulca: identification of the motile system. Plant Sci 120:127–137
Mittmann F, Brückner G, Zeidler M, Repp A, Abts T, Hartmann E, Hughes J (2004) Targeted knockout in Physcomitrella reveals direct actions of phytochrome in the cytoplasm. Proc Natl Acad Sci USA 101:13939–13944
Nozue K, Kanegae T, Imaizumi T, Fukuda S, Okamoto H, Yeh KC, Lagarias JC, Wada M (1998) A phytochrome from the fern Adiantum with features of the putative photoreceptor NPH1. Proc Natl Acad Sci USA 95:15826–15830
Paves H, Truve E (2007) Myosin inhibitors block accumulation movement of chloroplasts in Arabidopsis thaliana. Protoplasma 230:165–169
Peremyslov VV, Prokhnevsky AI, Aviser D, Dolja VV (2008) Two class XI myosins function in organelle trafficking and root hair development in Arabidopsis. Plant Physiol 146:1109–1116
Sakai T, Kagawa T, Kasahara M, Swartz TE, Christie JM, Briggs WR, Wada M, Okada K (2001) Arabidopsis nph1 and npl1: blue-light receptors that mediate both phototropism and chloroplast relocation. Proc Natl Acad Sci USA 98:6969–6974
Sato Y, Kadota A, Wada M (1999) Mechanically induced avoidance response of chloroplasts in fern protonemal cells. Plant Physiol 121:37–44
Sato Y, Wada M, Kadota A (2001) Choice of tracks, microtubules and/or actin filaments for chloroplast photo-movement is differentially controlled by phytochrome and a blue light receptor. J Cell Sci 114:269–279
Suetsugu N, Wada M (2007) Chloroplast photorelocation movement mediated by phototropin family proteins in green plants. Biol Chem 388:927–935
Tlalka M, Gabryś H (1993) Influence of calcium on blue-light-induced chloroplast movement in Lemna trisulca L. Planta 189:491–498
Tsuboi H, Suetsugu N, Wada M (2006) Negative phototropic response of rhizoid cells in the fern Adiantum capillus-veneris. J Plant Res 19:505–512
Tsuboi H, Suetsugu N, Kawai-Toyooka H, Wada M (2007) Phototropins and neochrome1 mediate nuclear movement in the fern Adiantum capillus-veneris. Plant Cell Physiol 48:892–896
Uenaka H, Kadota A (2007) Functional analyses of the Physcomitrella patens phytochromes in regulating chloroplast avoidance movement. Plant J 151:1050–1061
Wada M (2007) The fern as a model system to study photomorphogenesis. J Plant Res 120:3–16
Wada M (2008) Photoresponses in fern gametophytes. In: Ranker TA, Haufler CH (eds) Biology and evolution of ferns and lycophytes. Cambridge University Press, Cambridge, pp 3–48
Wada M, Furuya M (1978) Effects of narrow-beam irradiations with blue and far-red light on the timing of cell division in Adiantum gametophytes. Planta 138:85–90
Wada M, Kadota A, Furuya M (1983) Intracellular localization and dichroic orientation of phytochrome in plasma membrane and/or ectoplasm of a centrifuged protonema of fern Adiantum capillus-veneris. Plant Cell Physiol 24:1441–1447
Wada M, Kagawa T, Sato Y (2003) Chloroplast movement. Annu Rev Plant Biol 54:455–468
Wada M, Suetsugu N (2004) Plant organelle positioning. Curr Opin Plant Biol 7:626–631
Yatsuhashi H, Kobayashi H (1993) Dual involvement of phytochrome in light-oriented chloroplast movement in Dryopteris sparsa protonemata. J Photochem Photobiol B Biol 19:25–31
Acknowledgements
We thank Dr. John M. Christie for critical reading of the manuscript. We thank Dr. Noriyuki Suetsugu for useful discussion, critical reading, and revision of the manuscript. We thank Dr. Yutaka Kodama for writing macros for the Image J software and Ms. Ayumi Iwata for technical assistance. This work was partly supported by the Japanese Ministry of Education, Sports, Science, and Technology (MEXT 13139203, 17084006 to M.W.), the Japan Society of Promotion of Science (JSPS 13304061, 16107002 to M.W.), and a Research Fellowship for Young Scientists (to H.T).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Movie M1. This is a time-lapse video of the chloroplast movement shown in Fig. 2. Note the linear movement of the chloroplast toward the beam of blue light. Images were acquired in 15-s intervals (AVI 3651 kb)
Movie M2. This is a time-lapse video of the chloroplast movement shown in Fig. 4. Images were acquired in 15-s intervals (AVI 7112 kb)
10265_2008_199_MOESM3_ESM.avi
Movie M3. This is a time-lapse movie of the chloroplast movement in Arabidopsis thaliana used for the analysis of Fig. 7. Note the linear movement of the chloroplast toward the beams of blue light. Images were acquired in 15-s intervals (AVI 5149 kb)
Rights and permissions
About this article
Cite this article
Tsuboi, H., Yamashita, H. & Wada, M. Chloroplasts do not have a polarity for light-induced accumulation movement. J Plant Res 122, 131–140 (2009). https://doi.org/10.1007/s10265-008-0199-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10265-008-0199-z