
Abstract
Salt stress is one of the most serious factors limiting the productivity of agricultural crops. Increasing evidence has demonstrated that vacuolar Na+/H+ antiporters play a crucial role in plant salt tolerance. In the present study, we expressed the Suaeda salsa vacuolar Na+/H+ antiporter SsNHX1 in transgenic rice to investigate whether this can increase the salt tolerance of rice, and to study how overexpression of this gene affected other salt-tolerant mechanisms. It was found that transgenic rice plants showed markedly enhanced tolerance to salt stress and to water deprivation compared with non-transgenic controls upon salt stress imposition under outdoor conditions. Measurements of ion levels indicated that K+, Ca2+ and Mg2+ contents were all higher in transgenic plants than in non-transformed controls. Furthermore, shoot V-ATPase hydrolytic activity was dramatically increased in transgenics compared to that of non-transformed controls under salt stress conditions. Physiological analysis also showed that the photosynthetic activity of the transformed plants was higher whereas the same plants had reduced reactive oxygen species generation. In addition, the soluble sugar content increased in the transgenics compared with that in non-transgenics. These results imply that up-regulation of a vacuolar Na+/H+ antiporter gene in transgenic rice might cause pleiotropic up-regulation of other salt-resistance-related mechanisms to improve salt tolerance.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
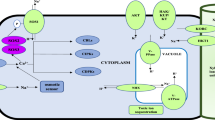
Salt stress affects plant growth and development in many different ways. To maintain growth and productivity, plants must adapt to stress conditions and exercise specific tolerance mechanisms. One mechanism involves removal of Na+ from the cytoplasm by transporting it into the vacuole via Na+/H+ exchangers driven by the electrochemical gradient of protons (H+) generated by the tonoplast H+-ATPase (V-ATPase) and H+-pyrophosphatase (V-PPase) (Niu et al. 1995; Qiu et al. 2004; Sze et al. 1999). In plants, Na+/H+ antiporters catalyze the exchange of Na+ for H+ across membranes and have a variety of functions, including maintenance of cellular ion homeostasis, and regulation of cytoplasmic pH and cell turgor (Horie and Schroede 2004). Increasing evidence has demonstrated that vacuolar Na+/H+ antiporters play a crucial role in plant salt tolerance. Improvement in salt tolerance evoked by overexpression of AtNHX1 was observed in Arabidopsis, tomato, Brassica and wheat (Apse et al. 1999; Zhang and Blumwald 2001; Zhang et al. 2001; Xue et al. 2004). Increased salt tolerance was also observed in transgenic rice carrying the OsNHX1 (NHX1 from Oryza sativa) and AgNHX1 (NHX1 from Atriplex gmelini) genes (Fukuda et al. 2004; Ohta et al. 2002). These results indicate that expression of a single Na+/H+ antiporter gene in plants can be effective in reducing Na+ toxicity. However, thus far the mechanisms underlying the enhancement of salt tolerance in Na+/H+ antiporter transformed plants are not yet clear (Tester and Davenpor 2003).
As mentioned above, several crop species have shown increased salt tolerance following transformation with the NHX1 gene originating from different plants; however, the NHX1 from the halophyte Suaeda salsa has not previously been overexpressed in rice. S. salsa is one of the most important leaf succulent euhalophytes in northern China, and can tolerate coastal seawater salinity and salinity fluctuations resulting from water evaporation and tidal inundation. In contrast to some other halophytic plants, S. salsa does not have salt glands or salt bladders on its leaves. Thus, this plant must compartmentalize the toxic Na+ in the vacuoles. Previous work in our laboratory has shown that expression of the S. salsa SsNHX1 is increased by salt stress. This result suggested that SsNHX1 might play an important role in the salt tolerance of S. salsa (Ma et al. 2004). In this work, we introduced the S. salsa vacuolar Na+/H+ antiporter SsNHX1 into rice to investigate whether the salt tolerance of the transgenic rice was increased. Furthermore, we attempted an in-depth study of how overexpression of this gene affects other salt-tolerant mechanisms. Possible mechanisms employed by SsNHX1-transgenic rice plants to improve salt tolerance are discussed.
Materials and methods
Vector construction and rice transformation
The Na+/H+ antiporter gene from S. salsa (SsNHX1) (Ma et al. 2004) was excised from the vector pMD18::SsNHX1 by digestion with KpnI and BamHI restriction enzymes, and the resulting fragment was inserted into the plant binary expression vector pROKII, between the cauliflower mosaic virus (CaMV) 35S promoter and octopine synthase terminator. The resulting plasmid, named pROKII/SsNHX1 (contained a selectable marker gene NPTII), was mobilized to Agrobacterium tumefaciens strain EHA105 and used for plant transformation.
For A. tumefaciens-mediated transformation, mature seeds of rice (Oryza sativa L. cv. Zhonghua No. 11) were used. Seeds were sterilized, and callus induction, co-cultivation with A. tumefaciens and plantlet regeneration were carried out by the method of Jang et al. (1999) and Liu et al. (1998) with some modifications. The transformed calli and plantlets were screened on Murashige and Skoog (1962) medium supplemented with 50 μg ml−1 kanamycin and 300 μg ml−1 cefotaxime. Putative primary transformants (T0 generation) were acclimatized in a greenhouse for 7 days, and then transplanted into big plastic pots (34 cm in diameter and 31 cm in height) filled with field soil. The transgenic plants were watered daily, and grew to maturity under outdoor conditions. Their offspring—T3 generation (each generation seeds were all screened on the selection medium)—were used for further experimental analysis.
Salt stress treatment of rice
Seeds of transgenic (T3 generation) and non-transgenic rice (cv. Zhonghua No. 11) were grown on selection and non-selection medium, respectively, for 2 weeks and then the T4 seedlings were transplanted into plastic pots (14 cm in diameter and 13 cm in height with small bottom outlet, each containing five seedlings) filled with field soil. These pots were immersed in big rectangle basins containing half-strength Hoagland nutrient solution, which was changed every 3 days under outdoor conditions. The average day/night temperature was 35°C/24°C, the relative humidity was 60%/80% and the maximum photosynthetically active radiation on a clear day was about 1,500 μmol m−2s−1.
After 3 weeks, the 5-week-old T4 seedlings were subjected to salt treatments under the same conditions. NaCl was dissolved in half-strength Hoagland nutrient solution and loaded in the big rectangle basins (the solution was changed every day). NaCl concentrations were stepped up in 50 mM/3 days increments until final concentrations (0, 150, 300 mM) were achieved. Three days after NaCl treatment, the seedlings were used for further physiological analysis.
RNA gel blot analysis
RNA gel blot analysis experiments were performed as described previously (Gao et al. 2003). Each lane contained 30 μg total RNA and a 600-bp fragment of the SsNHX1 5′-terminal region was used as the probe.
Measurement of relative water content
After salt treatment as described above, the relative water content (RWC) of the stressed shoots was measured. RWC was calculated as: RWC (%)=(FW−DW)/(TW−DW)×100, where FW is fresh weight, TW is turgid weight after soaking samples in deionized water for 24 h, and DW is dry weight after oven-drying samples at 85°C for 24 h. For each transgenic line, six measurements were carried out, each repeat containing ten shoots.
Analysis of root proton export capacity
To analyze root proton export capacity, transgenic and non-transgenic rice seeds were germinated and grown on Murashige and Skoog medium containing 150 mM NaCl for 15 days in a greenhouse with a photoperiod of 16 h light and 8 h darkness, a temperature of 25°C/20°C and a relative humidity of 60%/80%. Roots from each line (21 replicates) were used for the experiments according to the method of Yan et al. (2002) with some modifications. After washing with deionized water, roots were carefully spread on the surface of an agar sheet [0.75% (w/v) agar, 0.006% (w/v) bromocresol purple, 1 mM CaSO4 and 2.5 mM K2SO4, pH 6.5], gently pressed into the agar sheet and incubated for 7 h in darkness in a growth chamber. For study of the inhibitory effect of vanadate, 1 mM vanadate was included in the agar sheet.
Determination of the total K+, Na+, Ca2+ and Mg2+ concentration
Total K+, Na+, Ca2+ and Mg2+ contents of T4 transgenic rice plants were determined using an atomic absorption spectrophotometer (Z-8000, Hitachi, Tokyo) as described by Gao et al. (2003).
Isolation of membrane vesicles
For P-ATPase, V-ATPase and V-PPase hydrolytic activity analysis, tonoplast and plasma membrane vesicles were isolated from salt-stressed rice leaves and roots using a sucrose density gradient centrifugation technique as modified by Wang et al. (2001). The protein content was determined by the method of Bradford (1976).
Analysis of P-ATPase, V-ATPase and V-PPase hydrolytic activity
Activities of substrate hydrolysis of P-ATPase, V-ATPase and V-PPase were measured as described by Wang et al. (2001). P-ATPase activity was analyzed in the presence of 1 mM molybdate, 1 mM azide and 50 mM nitrate at 37°C. V-ATPase activity was analyzed in the presence of 1 mM molybdate, 1 mM azide and 1 mM vanadate at 37°C. V-PPase activity was analyzed in the presence of 1 mM molybdate, 1 mM azide, 50 mM nitrate and 1 mM vanadate at 37°C.
Measurement of the net photosynthetic rate
For net photosynthetic rate (P n) determination, rice growth and NaCl treatment were as above. P n was measured using an automatic photosynthetic measuring apparatus (Ciras-2, PPSystems, Hitchin, Hertfordshire, UK) as described by Qiu et al. (2003).
Direct localization of H2O2 in rice tissue
For detection of H2O2 generation, transgenic (T3 generation) and non-transgenic rice seeds were germinated and grown on Murashige and Skoog medium with or without 150 mM NaCl for 15 days in a greenhouse with a photoperiod of 16 h light and 8 h darkness, a temperature of 25°C/20°C and a relative humidity of 60%/80%. The middle part of the second leaf blade for each line (20 replicates) was excised for DAB (3,3-diaminobenzidine) staining and H2O2 localization detection as described by Talarczyk et al. (2002).
Malon dialdehyde content determination
Malon dialdehyde (MDA) content was tested by the method of Heath and Packer (1968).
Measurement of soluble sugar content
The soluble sugar content was measured by the modified method of Fairbairn (1953).
Results
SsNHX1 gene expression in transgenic rice
The Na+/H+ antiporter gene SsNHX1 (Ma et al. 2004; accession number AF370358) was introduced into a salt-sensitive rice cultivar (O. sativa cv. Zhonghua No. 11) by A. tumefaciens-mediated transformation under the control of the CaMV 35S promoter. A total of 26 transgenic lines carrying the SsNHX1 were identified by polymerase chain reaction (PCR) and reverse transcription (RT)-PCR analysis (data not shown), and two of these lines (Ss-21 and Ss-47) were selected for SsNHX1 expression analysis because their T3 offspring were homozygous. Transgene expression was examined by mRNA gel blot hybridization using total RNA from leaf tissues. As shown in Fig. 1, both transgenic lines accumulated the SsNHX1 transcript, but the mRNA levels were different in different lines (this result was similar to that of RT-PCR).

RNA gel blot analysis of SsNHX1 expression in transgenic rice plants. Each lane contained 30 μg total RNA. A 600-bp region of SsNHX1 5′ was used as the probe. Nt Non-transformed rice line; Ss-21, Ss-47 SsNHX1-transformed rice lines
Enhancing salt tolerance of transgenic rice
To test whether expression of the SsNHX1 in rice increased salt tolerance, 5-week-old T4 plants of the 26 transgenic lines and non-transgenic rice controls (cv. Zhonghua No. 11) were treated with 0, 150 and 300 mM NaCl for 3 days followed by non-stress treatment under outdoor conditions (see Materials and methods). Statistic analysis indicated that these transgenic lines showed markedly enhanced salt tolerance, and they behaved in similar manner upon salt stress imposition. Two of these transgenic lines (Ss-21 and Ss-47) were used for detailed experiments. No visual differences in growth and development between transgenic and non-transgenic rice lines were observed under non-salt stress conditions (Fig. 2a). However, when treated with 300 mM NaCl for 3 days (NaCl concentrations were stepped up in 50 mM/3 day increments until 300 mM was achieved for 3 days) under outdoor conditions, all non-transgenic rice plants wilted and showed salt-induced leaf rolling. In contrast, such symptoms were slight in SsNHX1 transgenic plants (Fig. 2b). To further examine the salt tolerance capacity of transgenic rice, changes in RWC in the same plant shoots caused by salt stress were measured. As shown in Fig. 2c, although RWC reduced in all experimental lines in response to the increase in NaCl concentration, the reduction was far greater in non-transformed lines compared to transgenic lines. Specifically, when plants were exposed to 300 mM NaCl for 3 days under outdoor conditions, the RWC decreased 49% in non-transgenics, but only 25% and 28% in transgenic plant lines Ss-21 and Ss-47, respectively. This result indicated that transgenics had a higher water retention capacity than non-transgenics. Similarly, both fresh weight and dry weight were also higher in transgenic lines than in non-transgenic controls (data not shown). Moreover, after removal of salt stress the transgenic plants recovered, whereas none of the control rice plants survived under the same conditions. A further remarkable characteristic of the transgenic plants was that their roots grew well and were able to export more protons out of root cells compared to the non-transgenic control plants (Fig. 3) upon salt treatment (see Materials and methods). Obviously, these results indicate increased resistance to salt stress in SsNHX1-transformed rice plants.

Growth (a, b) and relative water content (RWC) (c) of 5-week-old T4 transgenic rice plants upon NaCl treatment. a Before salt stress treatment. b Plants exposed to 300 mM NaCl for 3 days under outdoor conditions, NaCl concentrations were stepped up in 50 mM/3 day increments until a final concentration of 300 mM was achieved. c Plants treated with different concentrations of NaCl for 3 days under outdoor conditions. RWC (%)=(FW−DW)/(TW−DW)×100 (FW fresh weight, TW turgid weight after soaking samples in deionized water for 24 h, DW dry weight after oven-drying samples at 85°C for 24 h). For each line, six measurements were carried out, each repeat contain ten shoots. Nt, Non-transformed rice line; Ss-21, Ss-47, SsNHX1-transformed rice lines. Values represent means ±SD of six independent experiments (see Materials and methods)

Root proton export capacity in 15-day-old T4 SsNHX1-transgenic rice plants. Non-transgenic and T3 transgenic rice seeds were germinated and grown on MS+150 mM NaCl for 15 days in a growth chamber. After being washed with deionized water, roots were carefully spread on the surface of an agar sheet [0.75% (w/v) agar, 0.006% (w/v) bromocresol purple, 1 mM CaSO4 and 2.5 mM K2SO4, pH 6.5], gently pressed into the agar sheet and incubated for 7 h in darkness in a growth chamber. To assess the inhibitory effect of vanadate, 1 mM vanadate was included in the agar sheet. Nt Non-transformed rice line, Ss-21 SsNHX1-transformed rice line, V plus 1 mM vanadate, NV no vanadate; each line contained 21 replicates (see Materials and methods)
Reestablishing ion homeostasis in transgenic rice
Maintenance and reestablishment of cellular ion homeostasis during salt stress is crucial for plant survival and growth (Niu et al. 1995). To further investigate whether the increased salt tolerance of transgenic rice plants was related to ion levels, 5-week-old T4 plants of transgenic lines Ss-21 and Ss-47 and non-transgenic controls were exposed to 0, 150, 300 mM NaCl for 3 days under outdoor conditions, and total Na+, K+, Ca2+ and Mg2+ concentrations in the shoots were measured. As shown in Fig. 4, although Na+ increased (Fig. 4a) and both K+ (Fig. 4b) and the K+/Na+ ratio (Fig. 4c) decreased in both transgenic and non-transgenic plants following treatment with increased concentrations of NaCl, the transgenic rice plants contained more K+ and had a higher K+/Na+ ratio compared to non-transformed lines (Fig. 4b,c). We also observed 20–30% more Ca2+ in SsNHX1-transgenic plants than in non-transgenic plants (Fig. 4d) regardless of NaCl level, despite the fact that an increase in Ca2+ occurred in all experiment lines upon salt stress. Whereas Mg2+ increased at 150 mM NaCl and decreased at 300 mM NaCl, transgenic lines had a 30–45% higher Mg2+ concentration than non-transgenics (Fig. 4e). The ion concentrations in the transgenic plants were closely related to their root proton export capacity and salt tolerance.

Total Na+ (a), K+ (b), Ca2+ (d), Mg2+ (e) content, and K+/Na+ ratio (c) in 5-week-old T4 transgenic rice shoots treated with different concentrations of NaCl for 3 days under outdoor conditions. Nt Non-transformed rice line; Ss-21, Ss-47, SsNHX1-transformed rice lines. Values represent means ±SD of six independent experiments (see Materials and methods)
Increasing V-ATPase activity in transgenic rice plants
The data presented in Fig. 2 indicate that expression of SsNHX1 in transgenic rice can increase salt stress resistance, which may result from Na+ sequestration. Theoretically, the ability to compartmentalize sodium may result from a stimulation of the proton pumps that provide the driving force for increased sodium transport into the vacuole via Na+/H+ exchangers (Parks et al. 2002). Previously, no comparative analysis on V-ATPase and V-PPase activity has been performed in NHX1-transgenic plants. In the current study, to examine whether expressing SsNHX1 in transgenic rice induced an increase in H+-ATPase and H+-PPase activity, vesicles of plasma membrane and tonoplast were isolated by sucrose density gradient centrifugation from 5-week-old T4 transformed plants of Ss-21 and Ss-47 and non-transformed controls grown in the presence or absence of 150 mM NaCl for 3 days under outdoor conditions. Measurement of the hydrolytic activities of V-ATPase, V-PPase and P-ATPase revealed no obvious differences in these enzyme activities, either in shoots or in roots, between transgenics and non-transgenics under non-salt stress conditions (data not shown). However, shoot V-ATPase hydrolytic activity increased dramatically in transgenics when subjected to salt stress treatment, and was nearly twice that of non-transformed controls (Fig. 5a). In contrast, the activity of V-PPase and P-ATPase in the shoots was much lower than that of V-ATPase, and no marked differences in these enzyme activities could be seen between transgenic and non-transgenic rice. It was also observed that root V-ATPase activity was higher in transgenics than in non-transgenics, but lower compared to that of the shoot. Moreover, root P-ATPase activity in transgenics was also higher compared to non-transformed lines (Fig. 5b), in agreement with root proton export levels. These results support the idea that Na+ accumulation in SsNHX1-transgenic rice is energized by the H+ electrochemical gradient, particularly by V-ATPase.

Hydrolytic activity of V-ATPase, V-PPase and P-ATPase in 5-week-old T4 transgenic rice plant shoots (a) and roots (b) grown in the presence of 150 mM NaCl for 3 days under outdoor conditions. Nt Non-transformed rice line; Ss-21, Ss-47 SsNHX1-transformed rice lines. Values represent means ±SD of six independent experiments (see Materials and methods)
SsNHX1-transgenics maintain a higher level of photosynthesis
It is known that salt stress decreases plant photosynthetic capacity and causes inhibition of plant growth (Greenway and Munns 1980). To contribute to a better understanding of the role of SsNHX1 in transgenic rice response to salt stress, we analysed photosynthesis capacity by measuring the net photosynthetic rate (P n) and F v/F m ratio [chlorophyll fluorescence (F v/F m) ratio represents the activity of PSII] in transformants and non-transformants upon salt treatment. When 5-week-old T4 transgenic plants of Ss-21 and Ss-47 and non-transformed controls were treated with 0, 150, 300 mM NaCl for 3 days under outdoor conditions, P n declined in all experimental lines with increased NaCl concentrations in the watering solution (Fig. 6); however, a significantly greater decrease occurred in non-transformed lines compared to that in transgenic lines. For instance, P n decreased 79% in non-transgenic rice, but only 49% and 56% in transgenics Ss-21 and Ss-47, respectively, upon exposure to 300 mM NaCl for 3 days. Similarly, a decrease in the F v/F m ratio in the same plants was also observed. However, the F v/F m ratio of SsNHX1-transgenic plants was 15–20% higher than that of non-transformed plants upon 150 mM NaCl treatment for 5 days in a greenhouse (data not shown). These results implied that expression of SsNHX1 in transgenic rice might effectively reduce Na+ in the cytoplasm by sequestrating Na+ into vacuoles, thus protecting the photosynthetic machinery from Na+ toxicity and allowing a higher level of photosynthesis to be maintained.

Net photosynthetic rate of 5-week-old T4 SsNHX1-transgenic rice plants exposed to different concentrations of NaCl for 3 days under outdoor conditions. Nt Non-transformed rice line; Ss-21, Ss-47 SsNHX1-transformed rice lines. Values represent means ± SD of six independent experiments (see Materials and methods)
Reducing generation of reactive oxygen species in transformed rice
Salt stress has been reported to enhance production of reactive oxygen species (ROS; Mittova et al. 2004). To test whether the increased salt tolerance of SsNHX1 transgenic rice plants reduced production of ROS such as H2O2, the DAB (3,3-diaminobenzidine) staining method was applied to detect putative changes in H2O2 distribution in SsNHX1 transgenics Ss-21 and Ss-47 and non-transgenics upon salt stress imposition (Talarczyk et al. 2002). As shown in Fig. 7a, H2O2 caused by salt stress could be detected around the vascular bundle cells and mesophyll cells in all experimental plant leaf blades compared to non-stressed controls, but the generation of H2O2 was much less in transgenic lines Ss-21 and Ss-47 than in non-transgenic lines. The results coincided with the H2O2 (data not shown) and MDA (Fig. 7b) concentrations in the same plants. These results suggested that SsNHX1-transgenic rice might reduce H2O2 generation by increasing vacuolar Na+ compartmentation.

H2O2 generation (a) and malon dialdehyde (MDA) concentration (b) in SsNHX1-transgenic rice plants exposed to salt stress. a Visualization of H2O2 by DAB (3,3-diaminobenzidine) staining in 15-day-old T4 SsNHX1-transgenic rice leaf blades. The middle part of the second leaf blade for each line was excised for DAB staining and H2O2 localization. CK No salt stress; Nt non-transformed rice exposed to 150 mM NaCl for 15 days; Ss-21, Ss-47 transgenic rice exposed to 150 mM NaCl for 15 days; white arrows vascular bundle. Each line was represented by 20 plants (see Materials and methods). b Changes in MDA content caused by different concentrations of NaCl treatment for 3 days under outdoor conditions in 5-week-old T4 transgenic rice plant shoots. Values represent means ±SD of six independent experiments
Increased soluble sugars in transgenic rice plants
As presented above, transgenic rice carrying the SsNHX1 gene had enhanced salt tolerance, which may result from Na+ accumulation in the vacuole. As Na+ accumulates in the vacuole, osmotic potential in the cytoplasm must be balanced with that in the vacuole (Tester and Davenpor 2003). To detect any change in osmoprotectants in the transgenics, 5-week-old T4 transgenic plants of Ss-21 and Ss-47 and non-transformed controls were treated with 0, 150, 300 mM NaCl for 3 days under outdoor conditions, and both soluble sugar and proline contents in the shoots were measured. As shown in Fig. 8a, the soluble sugar content increased at 150 mM NaCl and decreased at 300 mM NaCl in all experimental plants, but in transgenic rice lines it was 1.83- to 2.32-fold higher compared to non-transgenic rice lines regardless of NaCl level. The proline content increased with increased NaCl concentration, and the non-transformed controls contained markedly more proline than the transgenics (Fig. 8b). Moreover, the proline content changed in a manner similar to that of MDA (Fig. 7b) and H2O2 (Fig. 7a) upon salt treatment. These results suggest that soluble sugars, but not proline, might play a role in osmotic adjustment under salt stress conditions in these transgenic rice plants.

Changes in soluble sugar (a) and proline (b) content in 5-week-old T4 transgenic rice plant shoots caused by different concentrations of NaCl treatment for 3 days under outdoor conditions. Nt Non-transformed rice line; Ss-21, Ss-47 SsNHX1-transformed rice lines. Values represent means ±SD of six independent experiments
Discussion
Higher K+, Ca2+ and Mg2+ ion levels might contribute to increased resistant to salt stress
In this work, we showed that although Na+ increased in both transgenic and non-transgenic rice plants in response to treatment with NaCl (Fig. 4a), the transgenics exhibited markedly enhanced salt tolerance (Fig. 2b, c) and water retention. These results are similar to those of Zhang et al. (2001) and Ohta et al. (2002). We ascribed these properties to the increased accumulation of Na+ in the vacuole due to expression of SsNHX1. Potassium is required for many functions in plants (Mäser et al. 2001). The ability of plants to maintain a high cytosolic K+/Na+ ratio is likely to be one of the key determinants in plant salt tolerance (Maathuis and Amtmann 1999). Xue et al. (2004) reported that AtNHX1-transgenic wheat had a higher K+ content and K+/Na+ ratio compared to non-transgenic plants; similar results were also observed in SsNHX1-transgenic rice in this study (Fig. 4b, c). Possibly, the relatively higher K+ level was related to the elevated Ca2+ levels in transgenics (Fig. 4d), because increased Ca2+ could stimulate net uptake of K+ by enhancing membrane integrity (Maathuis et al. 2003). Moreover, the relatively higher Ca2+ concentration in SsNHX1-transgenic rice might result from the higher capacity for root proton export, which in turn could enhance salt tolerance due to the fact that Ca2+ plays a number of roles in stabilizing cell walls and membranes and as a second messenger under salt stress conditions (Maathuis et al. 2003). Higher level of shoot Mg2+ content in the transgenic lines compared with non-transgenics was also observed (Fig. 4e). The significance of Mg2+ homeostasis has been particularly established with regard to its role in photosynthesis. Mg2+ is the central atom of the chlorophyll molecule, and fluctuations in Mg2+ levels in the chloroplast regulate the activity of key photosynthetic enzymes (Shaul 2002). The higher level of photosynthesis in transgenics may also be related to the increased Mg2+ content, in addition to the sequestering of Na+ in the vacuole and the protection of the photosynthetic machinery from Na+ toxicity due to the expression of SsNHX1. These results show that Na+, K+, Ca2+ and Mg2+ homeostasis is correlated with the level of salt-tolerance in these transgenic rice plants.
V-ATPase and P-ATPase may coordinate regulation of salt tolerance in transgenic rice
It has been suggested that most cations are transported against their electrochemical gradient by proton-coupled transporters rather than by primary ion pumps (Sze et al. 1999). Gaxiola et al. (2002) predicted that overexpression of AtNHX1 would increase vacuolar Na+ sequestration, implying that there is enough proton electrochemical gradient (PEG) to support the extra activity. Alternatively, AtNHX1 overexpression could trigger the activation of any of the vacuolar H+-pumps to provide the extra PEG required. However, to date there was no experimental evidence to link vacuolar Na+ accumulation with V-ATPase and/or V-PPase activity in vacuolar Na+/H+ antiporter-transgenic plants. In the present study, we demonstrated that, similar to the case of salt-tolerant plants (Barklaz et al. 1995; Wang et al. 2001; Parks et al. 2002), Na+ accumulated in SsNHX1-transgenic rice concurrently with increased V-ATPase activity induced by salt treatment (Fig. 5a, b). This result supported the idea that plants adapt to high Na+ levels in part by increasing the activity of proton pumps (Sze et al. 1999). One possible mechanism regulating the link between the activities of vacuolar Na+/H+ antiport and H+-transporting ATPase may be that ectopic expression of SsNHX1 in transgenic rice could trigger the activation of V-ATPase activity to provide the driving force for Na+ compartmentation. Another putative regulatory mechanism that could link the activities of vacuolar Na+/H+ antiport and V-ATPase may be inferred from the result that Ca2+ enhanced V-ATPase activity relatively more in transgenics (Fig. 4d), thereby increasing Na+ accumulation in the vacuole because the activity of V-ATPase was Ca2+-dependent upon salt stress (Han et al. 2005). Qiu et al. (2004) suggested that there might be coordinate regulation of the exchangers in the tonoplast and plasma membranes. We noticed that the activity of root P-ATPase was also higher in transgenics than in non-transgenic controls (Fig. 5b), which might imply that P-ATPase and V-ATPase are coordinately regulated in transgenic plant salt tolerance. Our results indicate that a similar mechanism possibly exists in transgenic rice. To our knowledge, this is the first experimental evidence linking V-ATPase activity with Na+ compartmentation in NHX1-transgenic plants.
Soluble sugars may be involved in osmotic adjustment in transgenic rice plants
As Na+ accumulated in the vacuole, osmotic balance between vacuolar and cytosolic compartments must be required. In the present study, we found that SsNHX1-transgenics contained more soluble sugars compared with non-transgenics upon salt stress imposition (Fig. 8a). Presumably, these soluble sugars resulted from the higher level of photosynthesis or from the metabolism of carbohydrate compounds in SsNHX1-transgenic rice upon exposure to salt stress. In contrast to other plants (Hong et al. 2000), the proline level was much lower in transformants than in non-transgenic controls (Fig. 8b), coinciding with levels of MDA and H2O2 in plants subjected to salt stress treatment. These results imply that soluble sugars, rather than proline, might contribute to osmotic adjustment in transgenic rice plants under salt stress conditions (Hu et al. 2000).
In addition, the lower generation of H2O2 and MDA in transgenic rice plants upon salt stress treatment supports the idea that expression of SsNHX1 could reduce Na+ toxicity to the cytoplasm and the photosynthetic machinery by increased sequestration into the vacuole.
Transgenic rice plants overexpressing AgNHX1 showed a strong tolerance to salt stress under the conditions of a 3-day exposure to 300 mM NaCl followed by non-stress treatments (Ohta et al. 2002); however, these transgenic studies were based on laboratory tests. It should be noticed that our results were obtained under outdoor conditions, approximating those of the field except for the controlled NaCl concentrations. These results may be more reliable than greenhouse data due to the fact that plants rarely experience stress from a single environmental source, and multi-stress interactions often occur under field conditions. Furthermore, in this study, we provided more detailed physiological evidence in SsNHX1-transgenic rice than that available in AgNHX1-transgenic rice plants.
In conclusion, the increase in Na+ accumulation and V-ATPase activity, together with other physiological characteristics of SsNHX1-transgenic rice suggest that up-regulation of SsNHX1 might cause pleiotropic up-regulation of a set of salt-tolerant-related mechanisms to confer higher levels of salt tolerance to the transgenic rice.
References
Apse MP, Aharon GS, Snedden WA, Blumwald E (1999) Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science 285:1256–1258
Barklaz BJ, Zingarelli L, Blumwald E, Smith JAC (1995) Tonoplast Na+/H+ antiport activity and its energization by the vacuolar H+-ATPase in the halophytic plant Mesembryan themum crystallinum L. Plant Physiol 109:549–556
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Fairbairn NJ (1953) A modified anthrone reagent. Chem Ind 4:86
Fukuda A, Chiba K, Maeda M, Nakamura A, Maeshima M, Tanaka Y (2004) Effect of salt and osmotic stresses on the expression of genes for the vacuolar H+-pyrophosphatase, H+-ATPase subunit A, and Na+/H+ antiporter from barley. Plant Cell Physiol 45:146–159
Gao XH, Ren ZH, Zhao YX, Zhang H (2003) Overexpression of SOD2 increases salt tolerance of Arabidopsis. Plant Physiol 133:1873–1881
Gaxiola RA, Fink GR, Hirschi KD (2002) Genetic manipulation of vacuolar proton pumps and transporters. Plant Physiol 129:967–973
Greenway H, Munns R (1980) Mechanisms of salt tolerance in non-halophytes. Annu Rev Plant Physiol 31:149–190
Han N, Shao Q, Lu CM, Wang BS (2005) The leaf tonoplast V-H+-ATPase activity of a C3 halophyte Suaeda salsa is enhanced by salt stress in a Ca-dependent mode. J Plant Physiol 162:267–274
Heath RL, Packer L (1968) Photoperoxidation in isolated chloroplasts I. Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophys 125:189–198
Hong ZL, Lakkineni K, Zhang ZM, Verma DPS (2000) Removal of feedback inhibition of Δ1-pyrroline-5-carboxylate synthetase results in increased proline accumulation and protection of plants from osmotic stress. Plant Physiol 122:1129–1136
Horie T, Schroede JI (2004) Sodium transporters in plants. Diverse genes and physiological functions. Plant Physiol 136:2457–2462
Hu YC, Schnyder H, Schmidhalter U (2000) Carbohydrate deposition and partitioning in elongating leaves of wheat under saline soil conditions. Aust J Plant Physiol 27:363–370
Jang IC, Nahm BH, Kim JK (1999) Subcellular targeting of green fluorescent protein to plastids in transgenic rice plants provides a high-level expression system. Mol Breed 5:453–461
Liu QQ, Zang JL, Wang ZY, Hong MM, Gu MH (1998) A highly efficient transformation system mediated by Agrobacterium tumefaciens in rice (Oryza sativa L.). Acta Phytophysiol Sin 24:259–271
Ma XL, Zhang Q, Shi HZ, Zhu JK, Zhao YX, Ma CL, Zhang H (2004) Molecular cloning and different expression of a vacuolar Na+/H+ antiporter gene in Suaeda salsa under salt stress. Biol Planta 48:219–225
Maathuis FJM, Amtmann A (1999) K+ nutrition and Na+ toxicity: the basis of cellular K+/Na+ ratios. Ann Bot 84:123–133
Maathuis FJ, Filatov V, Herzyk P, Krijger GC, Axelsen KB, Chen S, Green BJ, Li Y, Madagan KL, Sanchez-Fernandez R, Forde BG, Palmgren MG, Rea PA, Williams LE, Sanders D, Amtmann A (2003) Transcriptome analysis of root transporters reveals participation of multiple gene families in the response to cation stress. Plant J 35:675–692
Mäser P, Thomine S, Schroeder JI, Ward JM, Hirschi K, Sze H, Talke IN, Amtmann A, Maathuis FJM, Sanders D, Harper JF, Tchieu J, Gribskov M, Persans MW, Salt DE, Kim SA, Guerinot ML (2001) Phylogenetic relationships within cation transporter families of Arabidopsis. Plant Physiol 126:1646–1667
Mittova V, Guy M, Tal M, Volokita M (2004) Salinity up-regulates the antioxidative system in root mitochondria and peroxisomes of the wild salt-tolerant tomato species Lycopersicon pennellii. J Exp Bot 55:1105–1113
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassay with tobacco tissue cultures. Physiol Plant 15:473–497
Niu X, Bressan RA, Hasegawa PM, Pardo JM (1995) Ion homeostasis in NaCl stress environments. Plant Physiol 109:735–742
Ohta M, Hayashi Y, Nakashima A, Hamada A, Tanaka A, Nakamura T, Hayakawa T (2002) Introduction of a Na+/H+ antiporter gene from Atriplex gmelini confers salt tolerance to rice. FEBS Lett 532:279–282
Parks GE, Dietrich MA, Schumaker KS (2002) Increased vacuolar Na(+)/H(+) exchange activity in Salicornia bigelovii Torr. in response to NaCl. J Exp Bot 53:1055–1065
Qiu NW, Lu QT, Lu CM (2003) Photosynthesis, photosystem II efficiency and the xanthophyll cycle in the salt-adapted halophyte Atriplex centralasiatica. New Phytol 159:479–486
Qiu QS, Guo Y, Quintero FJ, Pardo JM, Schumaker KS, Zhu JK (2004) Regulation of vacuolar Na+/H+ exchange in Arabidopsis thaliana by the salt-overly-sensitive (SOS) pathway. J Biol Chem 279:207–215
Shaul O (2002) Magnesium transport and function in plants: the tip of the iceberg. Biometals 15:309–323
Sze H, Li X, Palmgren MG (1999) Energization of plant cell membranes by H+-pumping ATPases: regulation and biosynthesis. Plant Cell 11:677–689
Talarczyk A, Krzymowska M, Borucki W, Hennig J (2002) Effect of yeast CAT1 gene expression on response of tobacco plants to tobacco mosaic virus infection. Plant Physiol 129:1032–1044
Tester M, Davenpor R (2003) Na+ tolerance and Na+ transport in higher plants. Ann Bot 91:503–527
Wang BS, Lüttge U, Rataj R (2001) Effects of salt treatment and osmotic stress on V-ATPase and V-PPase in leaves of the halophyte Suaeda salsa. J Exp Bot 52:2355–2365
Xue ZY, Zhi D, Xue GP, Zhang H, Zhao YX, Xia GM (2004) Enhanced salt tolerance of transgenic wheat (Tritivum aestivum L.) expressing a vacuolar Na+/H+ antiporter gene with improved grain yields in saline soils in the field and a reduced level of leaf Na+. Plant Sci 167:849–859
Yan F, Zhu Y, Müller C, Zörb C, Schubert S (2002) Adaptation of H+-pumping and plasma membrane H+ ATPase activity in proteoid roots of white lupin under phosphate deficiency. Plant Physiol 129:50–63
Zhang HX, Blumwald E (2001) Transgenic salt tolerant tomato plants accumulate salt in foliage but not in fruit. Nat Biotechnol 19:765–768
Zhang HX, Hodson J, Williams JP, Blumwald E (2001) Engineering salt tolerant Brassica plants: characterization of yield and seed oil quality in transgenic plants with increased vacuolar sodium accumulation. Proc Natl Acad Sci USA 98:12832–12836
Acknowledgements
This work is supported financially by the National High Technology Research and Development Program of China (863 Program No. 2002AA629080) and the National Key Fundamental Research Program of China (973 Program No. 2006CB100104).
Author information
Authors and Affiliations
Corresponding author
Additional information
Fengyun Zhao and Zenglan Wang contributed equally to this work.
Note that this article has been retracted by the publisher.
An erratum to this article is available at http://dx.doi.org/10.1007/s10265-008-0214-4.
About this article
Cite this article
Zhao, F., Wang, Z., Zhang, Q. et al. RETRACTED ARTICLE: Analysis of the physiological mechanism of salt-tolerant transgenic rice carrying a vacuolar Na+/H+ antiporter gene from Suaeda salsa . J Plant Res 119, 95–104 (2006). https://doi.org/10.1007/s10265-005-0250-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10265-005-0250-2