
Abstract
Fish introduction, eutrophication and disappearance of aquatic vegetation are important disturbances of aquatic ecosystems, especially in plateau lakes, which are generally considered to be very vulnerable. Fish were introduced to Lake Dianchi, a eutrophic plateau lake in southwest China, in the late 1950s and 1970s. After the introduction, invasive fish became the dominant species, and the total fish yield increased. Meanwhile, the trophic level of Lake Dianchi had a tendency to increase in the past decades because of the increases in human activities in the watershed area. In addition, the area of aquatic vegetation decreased from more than 90 to 1.8% of the lake area from the 1950s to 2000. This study investigated the effects of fish introduction, eutrophication and aquatic vegetation on the diatom community of Lake Dianchi by examining the changes of microfossil diatom assemblage and abundance. Results showed that the absolute abundance and diatom assemblages changed after fish were introduced. The endemic species, Cyclotella rohomboideo-elliptica, disappeared with the introduction of fish and increasing trophic levels after 1958. Fragilaria crotonensis entered into the lake with the introduction of fish and gradually thrived in the lake after 1958. Diatom species numbers also decreased gradually from 21 to 9 from the past to present. Epiphytic diatoms disappeared with the decrease of aquatic vegetation after 1985. Our study indicated that eutrophication was the most important process determining diatom abundance, and fish introduction was a secondary process determining diatom abundance, while aquatic vegetation had a more important role in structuring the diatom community in this eutrophic plateau lake.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The Yunnan Plateau, located in southwestern China, ranges from the highest elevation of 6,740 m in the northwest to the lowest of 76 m in the southeast, and its average elevation is about 2,000 m above sea level. It has a distinctive monsoon climate with an annual mean temperature of about 15°C and an annual mean precipitation of about 1,000 mm (Nanjing Institute of Geography and Limnology 1989). This unique combination of topographic complexity and favorable moisture conditions in the region supports an enormous richness of biological diversity and high degrees of endemism (Zhang 1999). The lakes in Yunnan Plateau are a Global 200 Priority Ecoregion of the Palearctic Lake Ecosystems (Olsen and Dinerstein 1998). Thus, lakes in Yunnan Plateau have attracted many zoologists, botanists, naturalists and collectors since the end of the nineteenth century (Ley et al. 1963; Yang et al. 2005a, b; Li et al. 2007; Cui et al. 2008). Nevertheless, most of them have received little rigorous scientific examination, particularly of the relationship between diatom communities and the environment.
Lake Dianchi (24°51′N, 102°42′E) is a subtropical lake with a volume of 11.69 × 108 m3 and 297.9 km2 surface area located in central Yunnan Province; it is a eutrophic freshwater lake. The maximum and average depths are 6.5 and 2.93 m, respectively (Nanjing Institute of Geography and Limnology 1989; Wang and Dou 1998). The average annual rainfall is about 1,000 mm. In general, the rainy season occurs from May to October and the dry season from November to April (Wang and Dou 1998). With the development of agriculture, industry and urbanization in the region, increasing sewage discharge has had an effect on the trophic status and aquatic macrophytes (Luo et al. 2006). Aquatic macrophytes decreased from 90 to 1.8% of the lake area from the 1950s to 1990s. The community structure has been simplex and has deteriorated drastically (Yu et al. 2000). Numbers of aquatic macrophytes decreased from more than 100 species to 22 species from the 1950s to 1997 (Ley et al. 1963; Yu et al. 2000). In Lake Dianchi, TN and TP increased from 1988 to 2000, ranging from 1.02 to 11.89 mg/l and 0.109 to 1.06 mg/l, respectively (Tuo 2002; Meng 1999). Increasing TN and TP concentrations have caused a rapid outburst of cyanobacteria bloom in Lake Dianchi (Nanjing Institute of Geography and Limnology 1989; Wei et al. 1994).
The introduction of exotic fish from the Yangtze River to the lakes of Yunnan-Guizhou Plateau has led to significant ecological consequences in the lake ecosystem. Invasion by fish in Lake Dianchi has been divided into two periods. First, Silver and bighead carp and other fish were introduced into Lake Dianchi in 1958. Second, Neosalanx taihuensis was introduced into Lake Dianchi in 1979 (Wang and Dou 1998).
Diatoms are an abundant, diverse and important component of algal assemblages in freshwater lakes. They comprise a large portion of total algal biomass over a broad spectrum of lake trophic status (Hall and Smol 1999). Because diatoms are an abundant, high-quality food source for herbivores, they may play an important role in the aquatic food-web structure and functioning (Round et al. 1990). Because of the large number of ecologically sensitive species, which conveniently leave a fossil record, diatoms are widely used in paleoecological studies (Stoermer and Smol 1999).
Long-term data about the components (i.e., diatom assemblages) of the ecosystem are not available for Lake Dianchi concerning the full time scale of exotic fish, trophic status and aquatic vegetation impacts. Fortunately, lakes accumulate vast amounts of ecological and chemical information in their deep-water sediments, which can be used to reconstruct past changes in lake ecosystems (Hall and Smol 1999). Diatoms are well preserved in most lake sediments and are useful bioindicators in applied lake ecosystems studies. The purposes of this research were to describe diatom communities through sediments in Lake Dianchi and to discuss changes in diatom communities with the introduction of exotic fish, decreasing of aquatic macrophytes and increasing trophic status.
Materials and methods
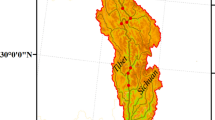
A box core from a 6.0-m-deep site in the north basin was taken by a Kajak gravity corer in June 2007 (Fig. 1). The subsampled core was sectioned by extrusion in 1-cm intervals from 0 to 10 cm and 2-cm intervals from 10 to 44 cm. Core sections were refrigerated for further processing. Diatom samples (about 0.2 g) were treated using HCl and H2O2 (Battarbee 1986). Four replicate sub-samples were potentially available from each sample. Diatom valves were enumerated on each of two prepared slides from each sample, resulting in two replicate abundance estimates. Diatoms were identified using oil immersion at 1,000× magnification under the Olympus microscope (BX51). Identification was performed using manuals such as Krammer and Lange-Bertalot (1988, 1991), Rumrich et al. (2000), Zhu and Chen (2000) and Lange-Bertalot (2001). 210Pb and 137Cs dating was measured in the Nanjing Institute of Geography and Limnology, the Chinese Academy of Sciences. Dried sediments were analyzed for 210Pb, 226Ra and 137Cs by direct gamma spectrometry using the Ortec HPGe GWL series of well-type, coaxial, low background, intrinsic germanium detectors (Appleby et al. 1986). 210Pb activity was determined via its direct gamma emissions at 46.5 keV and 226Ra via the 295.2 and 351.9 keV γ-rays emitted by its daughter isotope 214Pb following 3-week storage in sealed containers to allow secular equilibration to be established. 137Cs was measured by its emissions at 662.7 keV. Supported 210Pb in each sample was assumed to be in equilibrium with in situ 226Ra. Unsupported 210Pb activity at each depth was calculated by subtracting 226Ra activity from the total 210Pb activity (Wu et al. 2004).

Map of Lake Dianchi and the sample sites
Results
Sediment chronology was established by the stratigraphic decay profile of unsupported 210Pb activity and the constant rate of supply (CRS) model (Appleby and Oldfield 1978). The appearance and the peak of 137Cs activity at 31- and 25-cm depths represent the year 1952, the date when 137Cs fallout was first widely distributed, and 1963, the year of peak deposition in the northern hemisphere, respectively (Wu et al. 2007). The marker date for this peak agrees with the 210Pb date for these samples. The 210Pb dating data showed that the average deposition rate was 0.53 cm/year. There were 30 species from the core of Lake Dianchi. The diatom profile was divided into three zones by the absolute and relative abundance of the dominant species, Aulacoseira granulata, Cyclostephanos dubius, Stephanodiscus minutulus, Cyclotella rohomboideo-elliptica, Thalassiosira bramaputrae, Fragilaria crotonensis, Fragilaria aff. venter, Fragilaria parasitica and Gyrosigma acuminatum, and the absolute abundance of total diatoms, and the 210Pb dating data (Fig. 2).

Changes of absolute abundance (×108 valves g−1) and relative abundance (%) of different dominant taxa at core and the 210Pb dating data in Lake Dianchi
Zone 1: 12–0 cm, 1985–2007 a.d.
Zone 1 was characterized by predominant C. dubius whose relative abundance was more than 34%. The second dominant species was A. granulata, whose relative abundance ranged from 11.05 to 56.23%. The third species was S. minutulus, whose relative abundance ranged from 1.15 to 5.41%. Absolute abundance of total diatoms increased in this zone and ranged from 6.76 × 108 to 87.3 × 108 valves g−1. C. rohomboideo-elliptica and G. acuminatum did not appear in the zone. Relative abundance of F. aff. venter ranged from 0 to 6.27%. T. bramaputrae disappeared gradually. F. crotonensis, F. parasitica, Cyclotella menghiniana, Cyclotella stelligera and Fragilaria breviatum appeared in the zone, but their relative abundances were very low.
Zone 2: 26–10 cm, 1958–1985 a.d.
Zone 2 was dominated by A. granulata, whose relative abundance was more than 55%. The second dominant species was C. dubius, whose relative abundance ranged from 2.57 to 31.54%. The third species was T. bramaputrae, whose relative abundance was more than 8.5%. Absolute abundance of total diatoms increased in this zone and ranged from 1.94 × 108 to 6.39 × 108 valves g−1. S. minutulus, C. rohomboideo-elliptica, F. crotonensis, F. aff. venter, F. parasitica and G. acuminatum appeared in the zone, but their relative abundances were low, and less than 2.05% Achnanthes sp., Amphora lybica, Cymbella minuta, Cymbella prostata, Gomphonema germanii, Navicula sp., Nitzschia palea, Nitzschia sp., Cyclotella krammeri, Cyclotella ocellate and Stephanodiscus hantzschia appeared in the zone, but their relative abundances were very low, less than 0.5%.
Zone 3: 44–26 cm, 1924–1958 a.d.
Zone 3 was characterized by A. granulata, whose relative abundance was more than 65.49%. T. bramaputrae, whose relative abundance ranged from 10.65 to 18.64%, was the second dominant species. The third species was C. dubius, whose relative abundance ranged from 0.28 to 1.24%. Absolute abundance of total diatoms ranged from 0.9 × 108 to 2.38 × 108 valves g−1. Relative abundance of C. rohomboideo-elliptica ranged from 0.1 to 9.33%. Relative abundances of F. aff. venter, F. parasitica and G. acuminatum were more than 0.2%. F. crotonensis and S. minutulus did not appear in this zone. Achnanthes sp., A. lybica, Cymbella ehrenbergi, Cymbella gracilis, Cocconeis sp., Hipponica sp., G. acuminatum, Navicula sp., Nitzschia sp., Cyclotella menghiniana, C. ocellate and C. stelligera appeared in the zone, but their relative abundances were very low.
Discussion
Past studies indicated that C. rhomboideo-elliptica lives in dystrophic and mesotrophic environments (Ley et al. 1963; Li et al. 2007). A. granulata lives in mesotrophic to hypereutrophic, alkaline environments (Qi 1995). S. minutulus and C. dubius both are considered more eutrophic taxa (Hall and Smol 1999; Reavie et al. 1995; Qi 1995; Edlund et al. 1995; Yang et al. 2005a, b). C. dubius and S. minutulus are also present in samples from lakes of the middle and lower Yangtze River (Yang et al. 2005a, b, 2008). Changes in the absolute and relative abundance of A. granulata, C. rhomboideo-elliptica, S. minutulus and C. dubius in this study suggest that the trophic state of Lake Dianchi has gradually increased over the past 83 years. The composition of fossil species indicates that Lake Dianchi experienced being mesotrophic before 1958, then being eutrophic from 1958–1985, moving to being hypereutrophic after 1985 and throughout the past 83 years; therefore, significant changes in diatom assemblages suggest that limnological conditions during that time have not been stable.
Before 1958, the absolute abundance of total diatoms changed very little, and diatom species were relatively abundant. C. rhomboideo-elliptica, which was endemic to the Yunnan Plateau in modern freshwater habitats, existed in the period (Qi 1995). The Dianchi flora and fauna had developed a relatively high degree of endemism before 1958. Endemism is reflected at all levels of the food web and is especially evident in fish. In this period, because anthropogenic impacts were low, diatoms and fish still held a high degree of endemism (Skuja 1937; Ley et al. 1963; Wang and Dou 1998). This indicates that limnological conditions were stable during that time. After 1958, exotic fish, silver and bighead carp and other fish were introduced into the lake by human impacts (Wang and Dou 1998). In China, the disastrous impacts made by bighead and silver carp have been especially striking in many plateau lakes, where the continuous stocking of fingerlings has taken place on a wide scale since the late 1950s for increasing fish production (Xie and Chen 2001). The invaders have suppressed and in some instances eliminated the native or endemic species (Wang and Dou 1998; Li 2001). After 1958, F. crotonensis appeared in Lake Dianchi. The extent of taxa with widespread distributions may be related to human impact (Kociolek and Spaulding 2000). In this period, exotic fish from the Zhujiang and Yangtze River systems were introduced into the lake by humans. F. crotonensis was common in the Zhujiang and Yangtze River systems (Qi 1995), so it may have entered into the lake with the introduction of fish and gradually came to thrive in the lake. In this period the relative abundance of C. dubius gradually increased, and A granulata and C. rhomboideo-elliptica gradually decreased. C. rohomboideo-elliptica disappeared gradually after 1974. The increasing relative abundance of C. dubius and appearance of S. minutulus were a signal that Lake Dianchi had the beginning and the acceleration of eutrophication. These changes may be related to increasing fish production in the lake (Ley et al. 1963; Nanjing Institute of Geography and Limnology 1989). Silver and bighead carp and other introduced fish can cause a significant decline in zooplankton abundance (Xie and Yang 2000). Grazing pressure of diatoms by zooplankton decreased, which made the total diatom abundance increase from 1.94 × 108 to 6.39 × 108 valves g−1 from 1958–1985.
Aquatic macrophytes decreased from 90 to 1.8% of the lake area after the 1990s (Yu et al. 2000; Yang et al. 2004a, b). Aquatic macrophytes play a key role in structuring food webs in shallow lakes and in maintaining water transparency by direct and indirect effects on phytoplankton growth (Scheffer et al. 1993; Jeppesen et al. 1998; Van Donk and Van de Bund 2002). Empirical studies undertaken in temperate and some subtropical areas have shown that water transparency is generally high in lakes with high macrophyte cover (Jeppesen et al. 1990; Canfield and Hoyer 1992). In the 1950s, the water transparency of Lake Dianchi was 2 m, whereas it was 0.37–0.48 m in 2000 (Zhu 2004). Decreasing water transparency has an effect on diatom communities. Diatom numbers decreased to nine species, and C. dubius became the predominant species after 1985.
Neosalanx taihuensis was largely introduced into the lake in 1979 and became predominant after 1986 (Wang and Dou 1998; Liu and Zhu 1994). After N. taihuensis entered the lake, it replaced endemic fish as the predominant fish taxa (Li 2003). The introduced fish, N. taihuensis, has altered the food web. It has been suggested that the effects of zooplankton grazing and fish community structure on phytoplankton are stronger in eutrophic than in mesotrophic shallow lakes (Leibold 1989; Sarnelle 1993; Jeppesen et al. 2000). Schriver et al. (1995) observed in a mesocosm experiment that at increasing fish densities, phytoplankton shifted from small fast-growing species to cyanobacteria and dinoflagellates. With increasing production of N. taihuensis (2,319 tons in 1989), phytoplankton shifted from diatoms to Chlorophyta and cyanobacteria in Lake Dianchi (Wei et al. 1994; Luo et al. 2006; Zhang et al. 2006). The structure of the phytoplankton community is not only dependent on grazing pressure by herbivorous animals (the so-called top-down forces), but also is determined by nutrient conditions (the so-called bottom-up forces) (Shapiro 1980). With the development of agriculture, industry and urbanization in the region, increasing sewage discharge has had an effect on the trophic status (Li 2003; Wu et al. 2002). In Lake Dianchi, TN and TP showed substantial increases from 1988 to 2000, ranging from about 1.02 to 11.89 mg/l and 0.109 to 1.06 mg/l, respectively (Meng 1999; Tuo 2002; Mao et al. 2005). Increasing TN and TP concentrations have caused a rapid the outburst of cyanobacteria in Lake Dianchi (Nanjing Institute of Geography and Limnology 1989, Wei et al. 1994). The structure of diatom communities may change during cyanobacteria blooms, which may be due to shading by these blooms (Bjöjk-Ramberg and Änell 1985; Hansson 1992). A. granulata and cyanobacteria blooms grow in summer in Lake Dianchi, so maybe shading by cyanobacteria blooms results in an absolute and relative decrease in the abundance of A. granulata (Zhang et al. 2006). C. dubius mainly grows in the spring, and it can grow vastly in Lake Dianchi (Zhang et al. 2006). In this period, epiphytic diatoms (e.g., Gomphonema, Gyrosigma and Cymbella) disappeared entirely, which showed that aquatic macrophytes had mostly disappeared (Yang et al. 2004a, b). The results were consistent with the study of aquatic macrophytes in 2000 (Yu et al. 2000). In this period, the diatom number decreased to nine species, showing that Lake Dianchi was hypereutrophic after 1985. These changes in diatom communities implied that Lake Dianchi was a very sensitively poised system that was responsive to small changes in regional conditions as they affected nutrient loading in the lake as well as physical factors.
In conclusion, our study indicated that eutrophication and fish introduction were the most important processes determining diatom abundance, which increased from 1.94 × 108 to 87.3 × 108 valves g−1 during 1958–2007, while the number of diatom species decreased from 21 to 9 with the disappearance of aquatic vegetation, indicating that aquatic vegetation had a more important role in structuring the diatom community in this eutropic plateau lake. It is very difficult, if not impossible, to separate these from each other, because the two overlap, and it is hard to tell whether one or both are affecting the diatom abundance and community. To understand the mechanisms underlying the changes in the structure of the diatom community in Lake Dianchi, Yunnan Plateau, more comprehensive studies are needed.
References
Appleby PG, Oldfield F (1978) The calculation of 210Pb dates assuming a constant rate of supply of unsupported 210Pb to sediment. Catena 5:1–8
Appleby PG, Nolan PJ, Battarbee DW (1986) 210Pb dating by low background gamma counting. Hydrobiologia 141:21–27
Battarbee RW (1986) Diatom analysis. In: Berglund BE (ed) Handbook of holocene palaeoecology and palaeohydrology. Wiley, Chichester, pp 527–570
Bjöjk-Ramberg S, Änell C (1985) Production and chlorophyll concentration of epipelic and epilithic algae in fertilized and nonfertilized subarctic lakes. Hydrobiologia 126:213–219
Canfield DE, Hoyer MV (1992) Aquatic macrophytes and their relation to limnology of Florida Lakes. Bureau of Aquatic Plant Management, Florida Department of Natural Resources, Florida
Cui YD, Liu XQ, Wang HZ (2008) Macrozoobenthic community of Fuxian Lake, the deepest lake of southwest China. Limnologica 38:116–125
Edlund MB, Stoermer EF, Pilskaln CH (1995) Siliceous microfossil succession in the recent history of two basins in Lake Baikal, Siberia. J Paleolimnol 14:165–184
Hall RI, Smol JP (1999) Diatoms as indicators of lake eutrophication. In: Stoermer EF, Smol JP (eds) The diatoms: applications for the environmental and earth science. Cambridge University Press, London, pp 128–168
Hansson LA (1992) Factors regulating periphytic algal biomass. Limnol Oceanogr 33:121–128
Jeppesen E, Jensen JP, Kristensen P, Søndergaard M, Mortensen M, Sortkjaer O, Olrik K (1990) Fish manipulation as a lake restoration tool in shallow, eutrophic, temperate lakes. II. Threshold levels, longterm stability and conclusions. Hydrobiologia 200(201):219–227
Jeppesen E, Ma Søndergaard, Mo Søndergaard, Christoffersen K (1998) The structuring role of submerged macrophytes in lakes. Springer, New York
Jeppesen E, Jensen JP, Ma Søndergaard, Lauridsen T, Landkildehus F (2000) Trophic structure, species richness and biodiversity in Danish lakes: changes along a phosphorus gradient. Freshw Biol 45:201–218
Kociolek JP, Spaulding SA (2000) Freshwater diatom biogeography. Nova Hedwigia 71:223–241
Krammer K, Lange-Bertalot H (1988) Süßwasserflora von Mitteleuropa. Bacillariophyceae 1. Tei: Naviculaceae. Gustav Fisher Verlag, Jena
Krammer K, Lange-Bertalot H (1991) Süßwasserflora von Mitteleuropa. Bacillariophyceae 4. Tei: Achnanthaceae, Kritische Erganzungen zu Navicula (Lineolatae) und Gomphonema Gesamtliterturverzeichnis Tei 1-4. Gustav Fisher Verla, Jena
Lange-Bertalot H (2001) Navicula sensu stricto, 10 genera separated from Navicula sensu lato, Frustulia. In: Lange-Bertalot H, Gantner Verlag ARG, Ruggell KG(eds) Diatoms of Europe: diatoms of the European Inland waters and comparable habitats 2, pp 526
Leibold MA (1989) Resource edibility and the effects of predators and productivity on the outcome of trophic interactions. Am Nat 134:922–949
Ley S, Yu M, Li K, Ceng C, Chen J, Gao P, Huang H (1963) Limnological survey of the lakes of Yunnan Plateau (in Chinese). Oceanologia et Limnologia Sin 5(2):87–114
Li W (2001) The distribution of surviving Sinocyclocheilus grahami and other endemic fishes in Dianchi Lake Valley (in Chinese). J Jishou Univ 22:72–74
Li D (2003) The ecological analysis of the resources dynamics of Neosalanx taihuensis in Dianchi Lake (in Chinese). Trans Oceanol Limnol 2:69–73
Li YL, Gong Z, Shen J (2007) Freshwater diatoms of eight lakes in the Yunnan Plateau, China. J Freshw Ecol 22:169–171
Liu Z, Zhu S (1994) Food and feeding behavior of ice-fish (Neosalanx taihuensis Chen) in Dianchi Lake (in Chinese). Acta Zool Sin 40:253–261
Luo M, Duan C, Shen X, Yang L (2006) Environmental degradation and loss of species diversity in Dianchi Lake (in Chinese). Mar Fish 28:71–78
Mao Z, Peng W, Wang S, Zhou H (2005) Assessment and control measures for the eutrophication problems of the Dianchi Lake (in Chinese). J China Inst Water Resour Hydropower Res 3:135–142
Meng Y (1999) The development tendency analysis of nitrogen and phosphorous content in Waihai of Dianchi Lake (in Chinese). Yunnan Environ Sci 18:32–33
Nanjing Institute of Geography and Limnology (1989) Environments and sedimentation of fault lakes, Yunnan Province (in Chinese). Science Press, Beijing, pp 1–513
Olsen DM, Dinerstein E (1998) The global 200: a representation approach to conserving the earth’s most biologically valuable ecoregions. Conserv Biol 12:502–515
Qi YZ (1995) Flora Algarum Sinicarum Aquae Dulcis, Tomus IV Bacillariophyta Centricae (in Chinese). Science Press, Beijing
Reavie ED, Hall RI, Smol JP (1995) An expanded weighted-averaging model for inferring past total phosphorus concentrations from diatom assemblages in eutrophic British Columbia (Canada) Lakes. J Paleolimnol 14:49–67
Round FE, Crawford RM, Mann DG (1990) The diatoms biology and morphology of the genera. Cambridge University Press, Cambridge
Rumrich U, Lange-Bertalot H, Rumrich M (2000) Diatoms of the Andes (from Venzuela to Patagonia/Tierra del Fuego), Annotated diatom micrographs. Iconographia Diatomologica, 9. A.R.G. Gantner Verlag K.G
Sarnelle O (1993) Herbivore effects on phytoplankton succession in a eutrophic lake. Ecol Monogr 63:129–149
Scheffer M, Hosper SH, Meijer ML, Moss B, Jeppesen E (1993) Alternative equilibria in shallow lakes. Trends Ecol Evol 8:275–279
Schriver P, Bogestrand J, Jeppesen E, Søndergaard M (1995) Impact of submerged macrophytes on fish zooplankton–phytoplankton interactions: large-scale enclosure experiments in a shallow eutrophic lake. Freshw Biol 33:255–270
Shapiro J (1980) The importance of trophic-level interactions to the abundance and species composition of algae in lakes. In: Barica J, Moore LR (eds) Hypertrophic ecosystems. Junk Publisher, The Hague, pp 105–116
Skuja H (1937) Algae, in Handel-Mazzetti, Symbolae Sinicae. –Botanische Ergebnisse der expedition der Akademic der Wissenschaften in Wien nach Südwest-China. 1914–1918
Stoermer EF, Smol JP (1999) The diatoms: applications for the environmental and earth sciences. Cambridge University Press, London
Tuo Y (2002) Eutrophication of Dianchi and its trend and treatment. Yunnan Environ Sci 21:35–38
Van Donk E, Van de Bund WJ (2002) Impact of charophytes and other submerged macrophytes on plankton communities: what is the role of allelopathy? Aquat Bot 72:261–274
Wang SM, Dou HS (1998) Chinese lakes (in Chinese). Science Press, Beijing
Wei Y, Li R, Shi Z, Zhu H (1994) A survey of the phytoplankton and assessment of the water quality and trophic characteristic of three big lakes from Yunnan Plateau (in Chinese). In: Shi Z, Wei Y, Chen J, Li Y, Zhu H, Li R, Yao Y (eds) Compilation of report on survey of algal resources in the southwestern China. Science Press, Beijing, pp 371–404
Wu XH, Li YX, Hou CD (2002) The current situation analyses of Fuxian Lake (in Chinese). J Yuxi Teach Coll 18(2):66–68
Wu J, Gagan MK, Jiang X, Xia W, Wang S (2004) Sedimentary geochemical evidence for recent eutrophication of Lake Chenghai, Yunnan, China. J Paleolimnol 32:85–94
Wu J, Schleser GH, Lucke A, Li S (2007) A stable isotope record from freshwater lake shells of the eastern Tibetan Plateau, China, during the past two centuries. Boreas 36:38–46
Xie P, Chen Y (2001) Invasive carp in China’ s plateau lakes. Science 294:999–1000
Xie P, Yang Y (2000) Long-term changes of Copepoda community (1957–1996) in a subtropical Chinese lake stocked densely with planktivorous filter-feeding silver and bighead carps. J Plankton Res 22:1757–1778
Yang XD, Wang S, Kamenik C, Schmidt R, Shen J, Zhu L, Li S (2004a) Diatom assemblages and quantitative reconstruction for paleosalinity from a sediment core of Chencuo Lake, southern Tibet. Sci China Ser D Earth Sci 47(6):522–528
Yang Z, Zhang X, Liu A (2004b) Study on aquatic vegetation change in Dianchi Lake (in Chinese). J Southwest For Coll 24:27–30
Yang J, Zhang W, Feng W, Shen Y (2005a) Freshwater testate amoebae of nine Yunnan Plateau lakes, China. J Freshw Ecol 20:743–750
Yang XD, Dong X, Gao G, Pan H, Wu J (2005b) Relationships between surface sediment diatoms and summer water quality in shallow lakes of the middle and lower reaches of Yangtze River. J Integr Plant Biol 47(2):153–164
Yang XD, Anderson NJ, Dong X, Shen J (2008) Surface sediment diatom assemblages and epilimnetic total phosphorus in large, shallow lakes of the Yangtze floodplain: their relationships and implications for assessing long-term eutrophication. Freshw Biol 53:1273–1290
Yu G, Liu Y, Qiu C, Xu X (2000) Macrophyte succession in Dianchi Lake and relations with the environment (in Chinese). J Lake Sci 12:73–80
Zhang RZ (1999) Zoogeography of China (in Chinese). Science Press, Beijing
Zhang M, Li Y, Wang R (2006) Dynamic variation for the species of phytoplankton in Dianchi Lake (in Chinese). J Yunnan Univ 28:73–77
Zhu Y (2004) Succession tendency of water quality of Dianchi Lake and prevention countermeasures (in Chinese). Yunnan Environ Sci 23:97–100
Zhu H, Chen J (2000) Bacillariophyta of the Xizang Plateau (in Chinese). Science Press, Beijing
Acknowledgments
This work was supported by the National Basic Research Program of China (2008CB418103), projects of the National Sciences and Foundation of China (grant nos. 40602039, 40401064 and 40702029) and a project of CAS (KZCX1-YW-14). Special thanks are extended to Dr. Lu Zhang and Xiayun Xiao for their help. Dr. Shigeo Tsujimura and two anonymous referees are acknowledged for their helpful comments.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Gong, Zj., Li, Yl., Shen, J. et al. Diatom community succession in the recent history of a eutrophic Yunnan Plateau lake, Lake Dianchi, in subtropical China. Limnology 10, 247–253 (2009). https://doi.org/10.1007/s10201-009-0282-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10201-009-0282-8